
Adaptive Laboratory Evolution of Staphylococcus aureus Resistance to Vancomycin and Daptomycin: Mutation Patterns and Cross-Resistance

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Changes during Resistance Selection
2.1.1. Changes during Selection with Vancomycin
2.1.2. Changes during Selection with Daptomycin
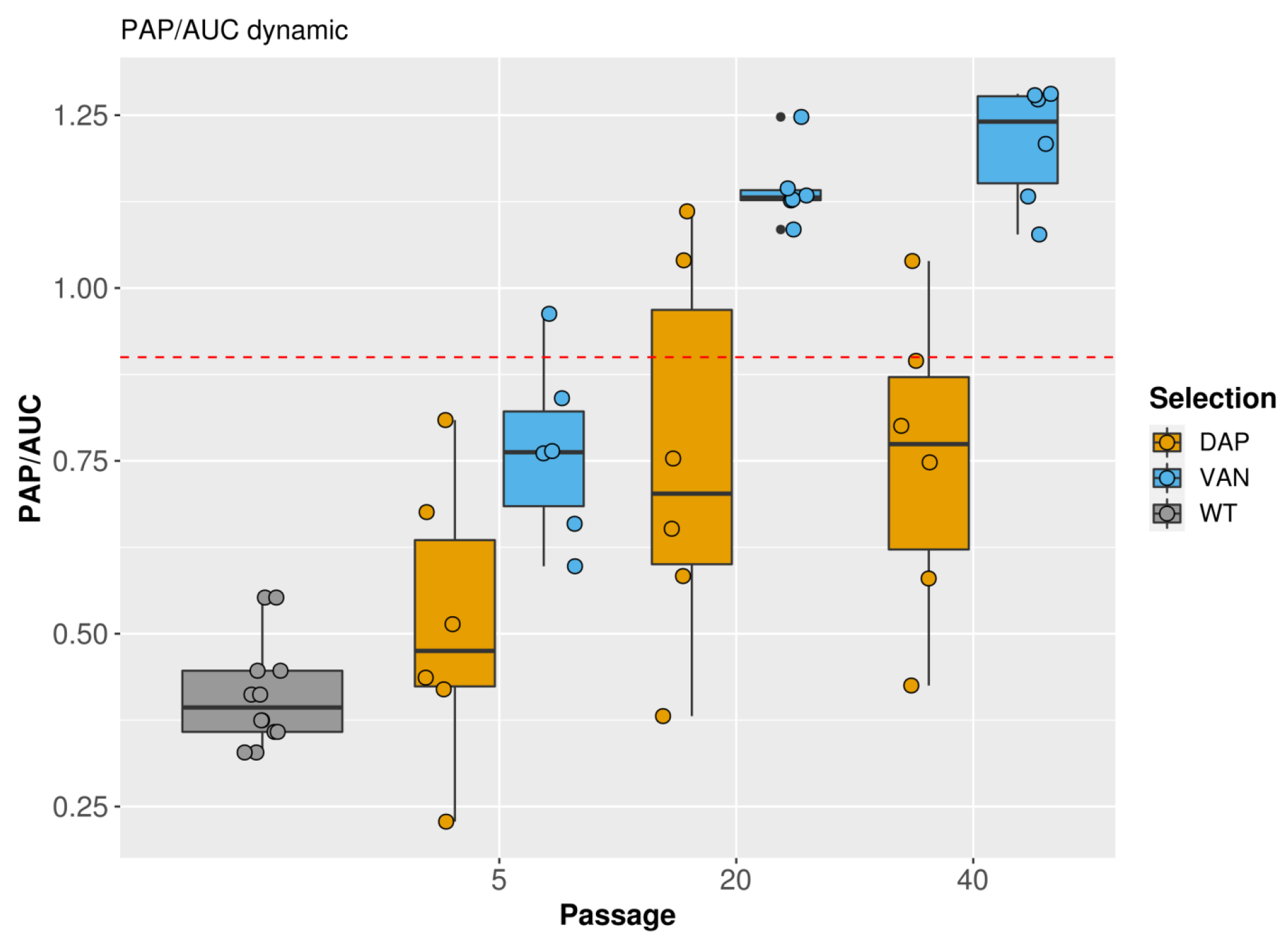
2.2. PAP Analysis
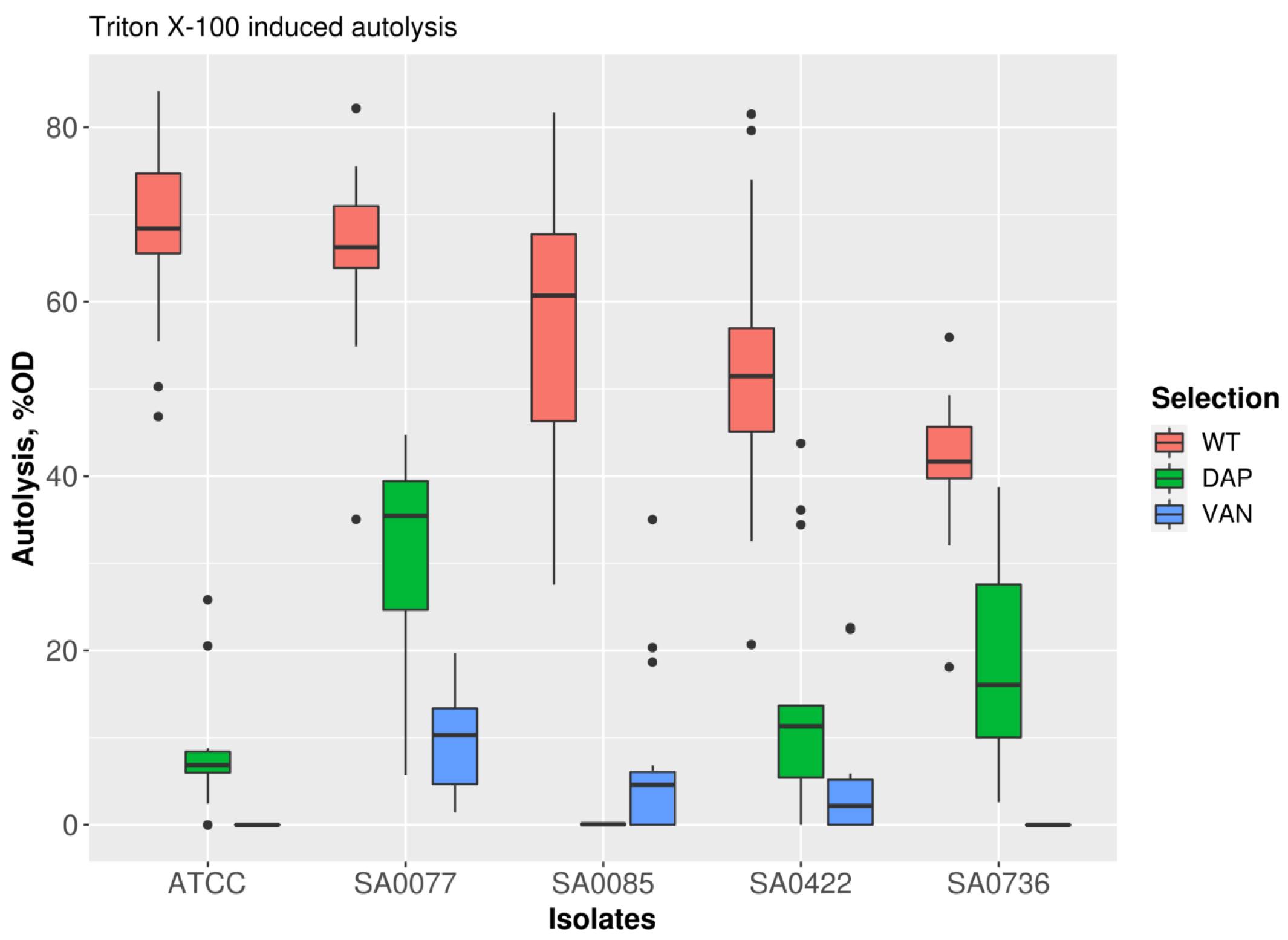
2.3. Induced Autolytic Activity
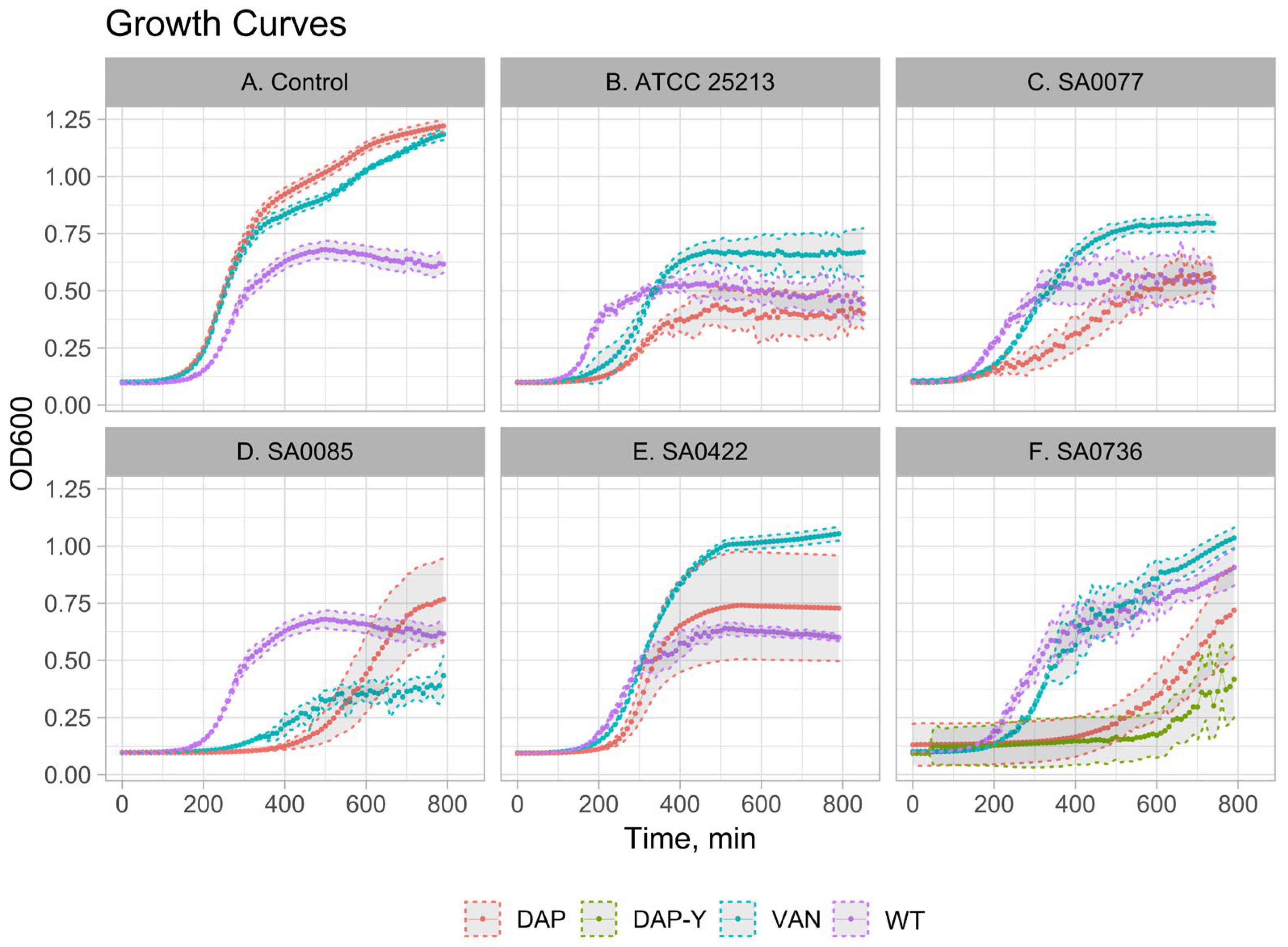
2.4. Growth Kinetics
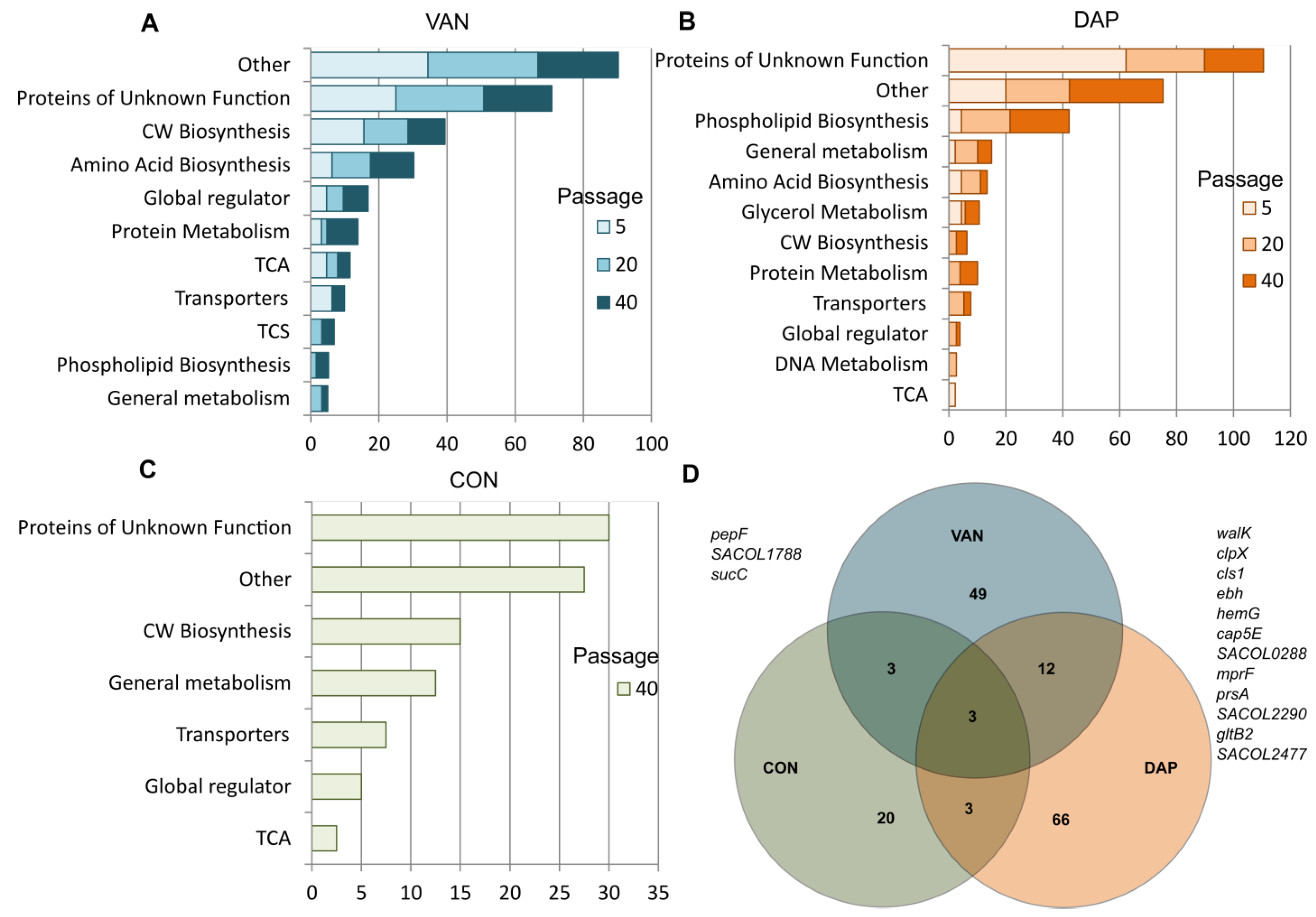
2.5. Whole-Genome Sequencing Analysis Data
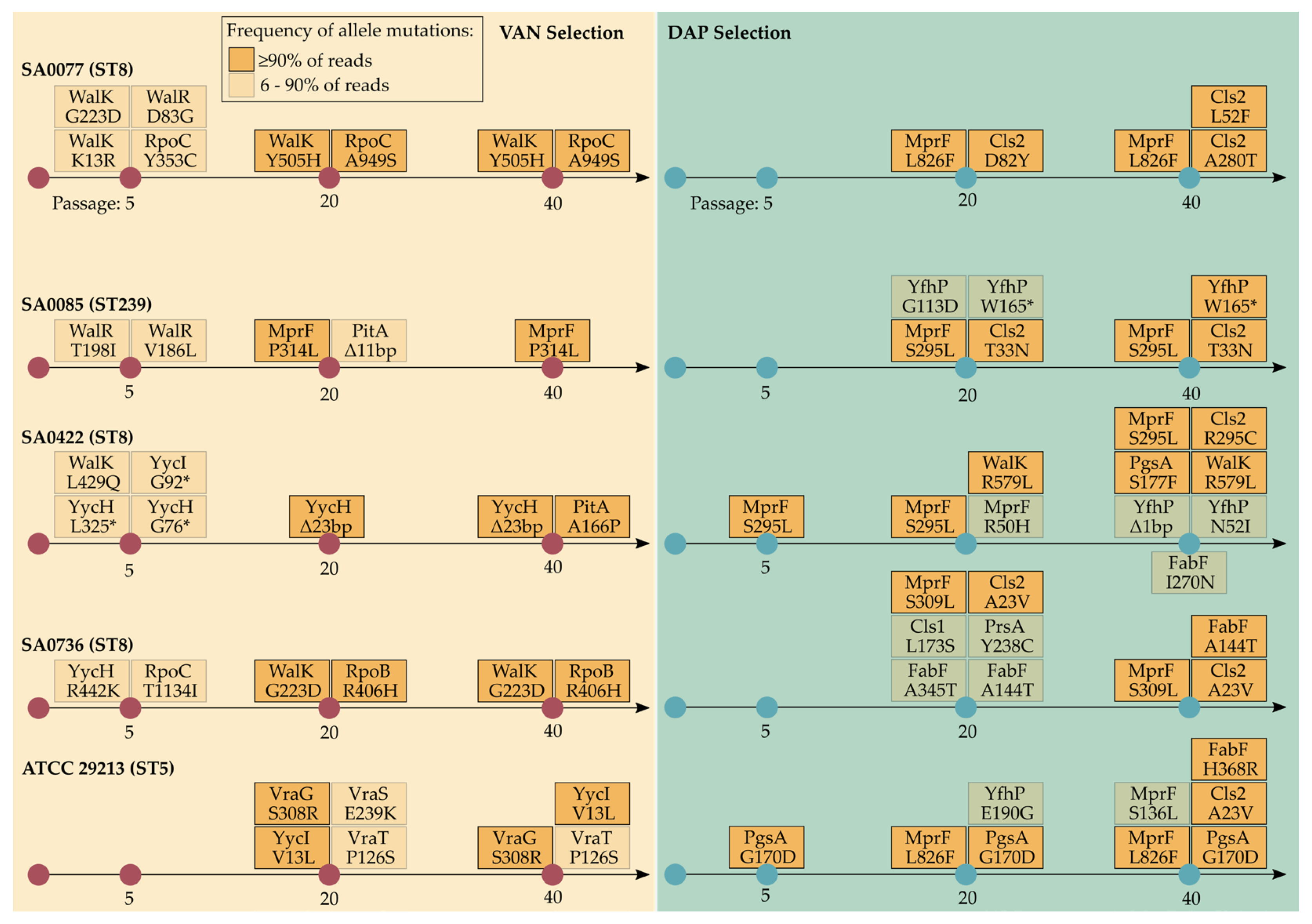
2.6. Genetic Changes during Selection with Vancomycin
2.7. Genetic Changes during Selection with Daptomycin
2.8. Analyzing the Contribution of the Deletion in SACOL1927 (yfhP) to Daptomycin Resistance
2.9. Genetic Changes during Passaging in Antibiotic-Free Media
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Susceptibility Testing
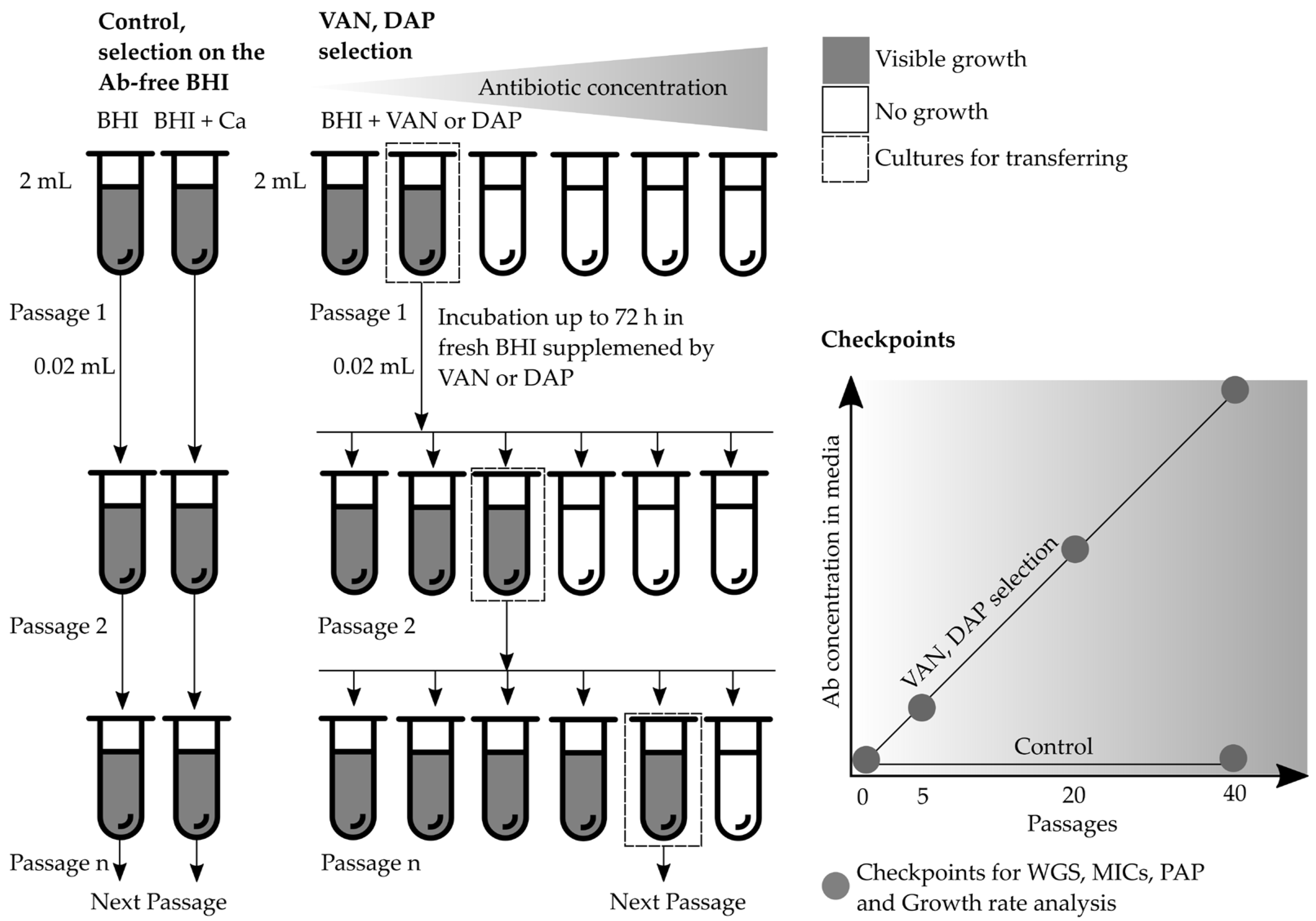
4.3. Multistep Resistance Selection
4.4. Population Analysis Profile
4.5. Measurement of Growth Rate
4.6. Measurement of Induced Autolytic Activity
4.7. S. aureus Genome Editing
4.8. Whole-Genome Sequencing
4.9. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shariati, A.; Dadashi, M.; Moghadam, M.T.; van Belkum, A.; Yaslianifard, S.; Darban-Sarokhalil, D. Global prevalence and distribution of vancomycin resistant, vancomycin intermediate and heterogeneously vancomycin intermediate Staphylococcus aureus clinical isolates: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 12689. [Google Scholar] [CrossRef]
- Watkins, R.R.; Holubar, M.; David, M.Z. Antimicrobial resistance in methicillin-resistant Staphylococcus aureus to newer antimicrobial agents. Antimicrob. Agents Chemother. 2019, 63, e01216-19. [Google Scholar] [CrossRef]
- Lin, Y.T.; Tsai, J.C.; Yamamoto, T.; Chen, H.J.; Hung, W.C.; Hsueh, P.R.; Teng, L.J. Emergence of a small colony variant of vancomycin-intermediate Staphylococcus aureus in a patient with septic arthritis during long-term treatment with daptomycin. J. Antimicrob. Chemother. 2016, 71, 1807–1814. [Google Scholar] [CrossRef]
- Koton, Y.; Or, Z.; Bisharat, N. Septic Thrombophlebitis with Persistent Methicillin-Resistant Staphylococcus aureus Bacteremia and de Novo Resistance to Vancomycin and Daptomycin. Infect. Dis. Rep. 2017, 9, 7008. [Google Scholar] [CrossRef]
- Kelley, P.G.; Gao, W.; Ward, P.B.; Howden, B.P. Daptomycin non-susceptibility in vancomycin-intermediate Staphylococcus aureus (VISA) and heterogeneous-VISA (hVISA): Implications for therapy after vancomycin treatment failure. J. Antimicrob. Chemother. 2011, 66, 1057–1060. [Google Scholar] [CrossRef]
- Cunha, B.A.; Pherez, F.M. Daptomycin resistance and treatment failure following vancomycin for methicillin-resistant Staphylococcus aureus (MRSA) mitral valve acute bacterial endocarditis (ABE). Eur. J. Clin. Microbiol. Infect. Dis. Off Publ. Eur. Soc. Clin. Microbiol. 2009, 28, 831–833. [Google Scholar] [CrossRef]
- Chen, C.J.; Huang, Y.C.; Chiu, C.H. Multiple pathways of cross-resistance to glycopeptides and daptomycin in persistent MRSA bacteraemia. J. Antimicrob. Chemother. 2015, 70, 2965–2972. [Google Scholar] [CrossRef]
- Nannini, E.; Murray, B.E.; Arias, C.A. Resistance or decreased susceptibility to glycopeptides, daptomycin, and linezolid in methicillin-resistant Staphylococcus aureus. Curr. Opin. Pharmacol. 2010, 10, 516–521. [Google Scholar] [CrossRef]
- Sulaiman, J.E.; Long, L.; Qian, P.Y.; Lam, H. Proteome profiling of evolved methicillin-resistant Staphylococcus aureus strains with distinct daptomycin tolerance and resistance phenotypes. Front. Microbiol. 2022, 13, 970146. [Google Scholar] [CrossRef]
- Matsuo, M.; Cui, L.; Kim, J.; Hiramatsu, K. Comprehensive identification of mutations responsible for heterogeneous vancomycin-intermediate Staphylococcus aureus (hVISA)-to-VISA conversion in laboratory-generated VISA strains derived from hVISA clinical strain Mu3. Antimicrob. Agents Chemother. 2013, 57, 5843–5853. [Google Scholar] [CrossRef]
- Katayama, Y.; Sekine, M.; Hishinuma, T.; Aiba, Y.; Hiramatsu, K. Complete Reconstitution of the Vancomycin-Intermediate Staphylococcus aureus Phenotype of Strain Mu50 in Vancomycin-Susceptible S. aureus. Antimicrob. Agents Chemother. 2016, 60, 3730–3742. [Google Scholar] [CrossRef]
- Hartmann, T.; Zhang, B.; Baronian, G.; Schulthess, B.; Homerova, D.; Grubmuller, S.; Kutzner, E.; Gaupp, R.; Bertram, R.; Powers, R.; et al. Catabolite control protein E (CcpE) is a LysR-type transcriptional regulator of tricarboxylic acid cycle activity in Staphylococcus aureus. J. Biol. Chem. 2013, 288, 36116–36128. [Google Scholar] [CrossRef]
- Su, M.; Davis, M.H.; Peterson, J.; Solis-Lemus, C.; Satola, S.W.; Read, T.D. Effect of genetic background on the evolution of Vancomycin-Intermediate Staphylococcus aureus (VISA). PeerJ 2021, 9, e11764. [Google Scholar] [CrossRef]
- Holmes, N.E.; Turnidge, J.D.; Munckhof, W.J.; Robinson, J.O.; Korman, T.M.; O’Sullivan, M.V.; Anderson, T.L.; Roberts, S.A.; Warren, S.J.; Coombs, G.W.; et al. Genetic and molecular predictors of high vancomycin MIC in Staphylococcus aureus bacteremia isolates. J. Clin. Microbiol. 2014, 52, 3384–3393. [Google Scholar] [CrossRef]
- Thitiananpakorn, K.; Aiba, Y.; Tan, X.E.; Watanabe, S.; Kiga, K.; Sato’o, Y.; Boonsiri, T.; Li, F.Y.; Sasahara, T.; Taki, Y.; et al. Association of mprF mutations with cross-resistance to daptomycin and vancomycin in methicillin-resistant Staphylococcus aureus (MRSA). Sci. Rep. 2020, 10, 16107. [Google Scholar] [CrossRef]
- Berti, A.D.; Baines, S.L.; Howden, B.P.; Sakoulas, G.; Nizet, V.; Proctor, R.A.; Rose, W.E. Heterogeneity of genetic pathways toward daptomycin nonsusceptibility in Staphylococcus aureus determined by adjunctive antibiotics. Antimicrob. Agents Chemother. 2015, 59, 2799–2806. [Google Scholar] [CrossRef]
- Blaskovich, M.A.T.; Hansford, K.A.; Butler, M.S.; Jia, Z.; Mark, A.E.; Cooper, M.A. Developments in Glycopeptide Antibiotics. ACS Infect. Dis. 2018, 4, 715–735. [Google Scholar] [CrossRef]
- Binda, E.; Marinelli, F.; Marcone, G.L. Old and New Glycopeptide Antibiotics: Action and Resistance. Antibiotics 2014, 3, 572–594. [Google Scholar] [CrossRef]
- Karlowsky, J.A.; Nichol, K.; Zhanel, G.G. Telavancin:Mechanisms of action, in vitro activity, and mechanisms of resistance. Clin. Infect. Dis. Off Publ. Infect. Dis. Soc. Am. 2015, 61 (Suppl. 2), S58–S68. [Google Scholar] [CrossRef]
- Werth, B.J.; Jain, R.; Hahn, A.; Cummings, L.; Weaver, T.; Waalkes, A.; Sengupta, D.; Salipante, S.J.; Rakita, R.M.; Butler-Wu, S.M. Emergence of dalbavancin non-susceptible, vancomycin-intermediate Staphylococcus aureus (VISA) after treatment of MRSA central line-associated bloodstream infection with a dalbavancin- and vancomycin-containing regimen. Clin. Microbiol. Infect. Off Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2018, 24, 429.e1–429.e5. [Google Scholar] [CrossRef]
- Steele, J.M.; Seabury, R.W.; Hale, C.M.; Mogle, B.T. Unsuccessful treatment of methicillin-resistant Staphylococcus aureus endocarditis with dalbavancin. J. Clin. Pharm. Ther. 2018, 43, 101–103. [Google Scholar] [CrossRef]
- Kussmann, M.; Karer, M.; Obermueller, M.; Schmidt, K.; Barousch, W.; Moser, D.; Nehr, M.; Ramharter, M.; Poeppl, W.; Makristathis, A.; et al. Emergence of a dalbavancin induced glycopeptide/lipoglycopeptide non-susceptible Staphylococcus aureus during treatment of a cardiac device-related endocarditis. Emerg. Microbes Infect. 2018, 7, 202. [Google Scholar] [CrossRef]
- Li, S.; Yin, Y.; Chen, H.; Wang, Q.; Wang, X.; Wang, H. Fitness Cost of Daptomycin-Resistant Staphylococcus aureus Obtained from in Vitro Daptomycin Selection Pressure. Front. Microbiol. 2017, 8, 2199. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Jiang, L.; Han, W.; Xie, X.; Jin, Y.; He, X.; Wu, R. Novel Mutation Sites in the Development of Vancomycin- Intermediate Resistance in Staphylococcus aureus. Front. Microbiol. 2016, 7, 2163. [Google Scholar] [CrossRef]
- Cui, L.; Isii, T.; Fukuda, M.; Ochiai, T.; Neoh, H.M.; Camargo, I.L.; Watanabe, Y.; Shoji, M.; Hishinuma, T.; Hiramatsu, K. An RpoB mutation confers dual heteroresistance to daptomycin and vancomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2010, 54, 5222–5233. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, X.; Liu, X.; Chen, C.; Sun, B. Mechanism of reduced vancomycin susceptibility conferred by walK mutation in community-acquired methicillin-resistant Staphylococcus aureus strain MW2. Antimicrob. Agents Chemother. 2015, 59, 1352–1355. [Google Scholar] [CrossRef]
- Hu, Q.; Peng, H.; Rao, X. Molecular Events for Promotion of Vancomycin Resistance in Vancomycin Intermediate Staphylococcus aureus. Front. Microbiol. 2016, 7, 1601. [Google Scholar] [CrossRef]
- Mehta, S.; Cuirolo, A.X.; Plata, K.B.; Riosa, S.; Silverman, J.A.; Rubio, A.; Rosato, R.R.; Rosato, A.E. VraSR two-component regulatory system contributes to mprF-mediated decreased susceptibility to daptomycin in in vivo-selected clinical strains of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 92–102. [Google Scholar] [CrossRef]
- Ernst, C.M.; Slavetinsky, C.J.; Kuhn, S.; Hauser, J.N.; Nega, M.; Mishra, N.N.; Gekeler, C.; Bayer, A.S.; Peschel, A. Gain-of-Function Mutations in the Phospholipid Flippase MprF Confer Specific Daptomycin Resistance. mBio 2018, 9, e01659-18. [Google Scholar] [CrossRef]
- Sulaiman, J.E.; Wu, L.; Lam, H. Mutation in the Two-Component System Regulator YycH Leads to Daptomycin Tolerance in Methicillin-Resistant Staphylococcus aureus upon Evolution with a Population Bottleneck. Microbiol. Spectr. 2022, 10, e0168722. [Google Scholar] [CrossRef]
- Cameron, D.R.; Jiang, J.H.; Kostoulias, X.; Foxwell, D.J.; Peleg, A.Y. Vancomycin susceptibility in methicillin-resistant Staphylococcus aureus is mediated by YycHI activation of the WalRK essential two-component regulatory system. Sci. Rep. 2016, 6, 30823. [Google Scholar] [CrossRef]
- Boyle-Vavra, S.; Challapalli, M.; Daum, R.S. Resistance to autolysis in vancomycin-selected Staphylococcus aureus isolates precedes vancomycin-intermediate resistance. Antimicrob. Agents Chemother. 2003, 47, 2036–2039. [Google Scholar] [CrossRef]
- Howden, B.P.; McEvoy, C.R.; Allen, D.L.; Chua, K.; Gao, W.; Harrison, P.F.; Bell, J.; Coombs, G.; Bennett-Wood, V.; Porter, J.L.; et al. Evolution of multidrug resistance during Staphylococcus aureus infection involves mutation of the essential two component regulator WalKR. PLoS Pathog. 2011, 7, e1002359. [Google Scholar] [CrossRef]
- Friedman, L.; Alder, J.D.; Silverman, J.A. Genetic changes that correlate with reduced susceptibility to daptomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 2137–2145. [Google Scholar] [CrossRef]
- Hines, K.M.; Shen, T.; Ashford, N.K.; Waalkes, A.; Penewit, K.; Holmes, E.A.; McLean, K.; Salipante, S.J.; Werth, B.J.; Xu, L. Occurrence of cross-resistance and beta-lactam seesaw effect in glycopeptide-, lipopeptide- and lipoglycopeptide-resistant MRSA correlates with membrane phosphatidylglycerol levels. J. Antimicrob. Chemother. 2020, 75, 1182–1186. [Google Scholar] [CrossRef]
- Song, Y.; Lunde, C.S.; Benton, B.M.; Wilkinson, B.J. Studies on the mechanism of telavancin decreased susceptibility in a laboratory-derived mutant. Microb. Drug Resist. 2013, 19, 247–255. [Google Scholar] [CrossRef]
- Werth, B.J.; Ashford, N.K.; Penewit, K.; Waalkes, A.; Holmes, E.A.; Ross, D.H.; Shen, T.; Hines, K.M.; Salipante, S.J.; Xu, L. Dalbavancin exposure in vitro selects for dalbavancin-non-susceptible and vancomycin-intermediate strains of methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. Off Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2021, 27, 910.e1–910.e8. [Google Scholar] [CrossRef]
- Camargo, I.L.; Neoh, H.M.; Cui, L.; Hiramatsu, K. Serial daptomycin selection generates daptomycin-nonsusceptible Staphylococcus aureus strains with a heterogeneous vancomycin-intermediate phenotype. Antimicrob. Agents Chemother. 2008, 52, 4289–4299. [Google Scholar] [CrossRef]
- Gostev, V.; Sopova, J.; Kalinogorskaya, O.; Tsvetkova, I.; Sidorenko, S. Selection of Resistance to Daptomycin in Methicillin-Resistant Staphylococcus aureus: Role of Homo- and Hetero-Mutations. Russ. J. Genet. 2020, 56, 282–291. [Google Scholar] [CrossRef]
- Fait, A.; Seif, Y.; Mikkelsen, K.; Poudel, S.; Wells, J.M.; Palsson, B.O.; Ingmer, H. Adaptive laboratory evolution and independent component analysis disentangle complex vancomycin adaptation trajectories. Proc. Natl. Acad. Sci. USA 2022, 119, e2118262119. [Google Scholar] [CrossRef]
- Mechler, L.; Bonetti, E.J.; Reichert, S.; Flotenmeyer, M.; Schrenzel, J.; Bertram, R.; Francois, P.; Gotz, F. Daptomycin Tolerance in the Staphylococcus aureus pitA6 Mutant Is Due to Upregulation of the dlt Operon. Antimicrob. Agents Chemother. 2016, 60, 2684–2691. [Google Scholar] [CrossRef]
- De Carvalho, C.; Taglialegna, A.; Rosato, A.E. Impact of PrsA on membrane lipid composition during daptomycin-resistance-mediated beta-lactam sensitization in clinical MRSA strains. J. Antimicrob. Chemother. 2021, 77, 135–147. [Google Scholar] [CrossRef]
- Lopatkin, A.J.; Bening, S.C.; Manson, A.L.; Stokes, J.M.; Kohanski, M.A.; Badran, A.H.; Earl, A.M.; Cheney, N.J.; Yang, J.H.; Collins, J.J. Clinically relevant mutations in core metabolic genes confer antibiotic resistance. Science 2021, 371, eaba0862. [Google Scholar] [CrossRef]
- Deatherage, D.E.; Traverse, C.C.; Wolf, L.N.; Barrick, J.E. Detecting rare structural variation in evolving microbial populations from new sequence junctions using breseq. Front. Genet. 2014, 5, 468. [Google Scholar] [CrossRef]
- Pfeltz, R.F.; Schmidt, J.L.; Wilkinson, B.J. A microdilution plating method for population analysis of antibiotic-resistant staphylococci. Microb. Drug Resist. 2001, 7, 289–295. [Google Scholar] [CrossRef]
- Sprouffske, K.; Wagner, A. Growthcurver: An R package for obtaining interpretable metrics from microbial growth curves. BMC Bioinform. 2016, 17, 172. [Google Scholar] [CrossRef]
- Penewit, K.; Holmes, E.A.; McLean, K.; Ren, M.; Waalkes, A.; Salipante, S.J. Efficient and Scalable Precision Genome Editing in Staphylococcus aureus through Conditional Recombineering and CRISPR/Cas9-Mediated Counterselection. mBio 2018, 9, e00067-18. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Brown, J.; Pirrung, M.; McCue, L.A. FQC Dashboard: Integrates FastQC results into a web-based, interactive, and extensible FASTQ quality control tool. Bioinformatics 2017, 33, 3137–3139. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; GGenome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Barrick, J.E.; Colburn, G.; Deatherage, D.E.; Traverse, C.C.; Strand, M.D.; Borges, J.J.; Knoester, D.B.; Reba, A.; Meyer, A.G. Identifying structural variation in haploid microbial genomes from short-read resequencing data using breseq. BMC Genomics 2014, 15, 1039. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Selecting Agent | Passage | MIC, µg/mL | |||||
|---|---|---|---|---|---|---|---|---|
| VAN | TEC | DAP | ORI | DAL | TLV | |||
| SA0077 | VAN | 0 | 1 | 0.25 | 0.25 | <0.03 | 0.03 | 0.125 |
| 5 | 4 | 8 | 2 | 0.125 | 0.25 | 0.06 | ||
| 20 | 8 | 8 | 2 | 2 | 1 | 0.5 | ||
| 40 | 8 | 8 | 2 | 2 | 2 | 1 | ||
| SA0085 | VAN | 0 | 1 | 0.125 | 0.5 | <0.03 | <0.016 | 0.06 |
| 5 | 4 | 1 | 1 | 0.06 | 0.06 | 0.06 | ||
| 20 | 4 | 2 | 2 | 2 | 0.5 | 0.25 | ||
| 40 | 8 | 8 | 2 | 1 | 0.25 | 0.25 | ||
| SA0422 | VAN | 0 | 1 | 0.5 | 0.25 | <0.03 | 0.03 | 0.125 |
| 5 | 2 | 2 | 1 | 0.06 | 0.125 | 0.25 | ||
| 20 | 4 | 4 | 1 | 0.25 | 0.5 | 0.5 | ||
| 40 | 4 | 4 | 2 | 0.5 | 1 | 1 | ||
| SA0736 | VAN | 0 | 0.5 | 0.25 | 0.25 | <0.03 | 0.03 | 0.125 |
| 5 | 4 | 4 | 0.5 | 0.25 | 0.125 | 0.25 | ||
| 20 | 8 | 8 | 0.5 | 2 | 1 | 0.5 | ||
| 40 | 8 | 8 | 2 | 2 | 1 | 0.5 | ||
| ATCC 29213 | VAN | 0 | 0.25 | 0.06 | 0.125 | 0.06 | <0.016 | <0.03 |
| 5 | 0.25 | 0.06 | 0.25 | 0.06 | <0.016 | <0.03 | ||
| 20 | 2 | 2 | 2 | 0.06 | <0.016 | 0.125 | ||
| 40 | 8 | 8 | 2 | 4 | 2 | 0.5 | ||
| SA0077 | DAP | 0 | 1 | 0.25 | 0.25 | <0.03 | 0.03 | 0.125 |
| 5 | 2 | 0.125 | 0.5 | 2 | <0.03 | 0.03 | ||
| 20 | 4 | 0.25 | 8 | 4 | 0.125 | 0.125 | ||
| 40 | 4 | 0.25 | 32 | 4 | 0.125 | 0.125 | ||
| SA0085 | DAP | 0 | 1 | 0.125 | 0.5 | <0.03 | <0.016 | 0.06 |
| 5 | 1 | 0.125 | 0.5 | <0.03 | <0.016 | 0.06 | ||
| 20 | 2 | 8 | 32 | 0.06 | 0.03 | 0.06 | ||
| 40 | 4 | 8 | >64 | 1 | 0.125 | 0.25 | ||
| SA0422 | DAP | 0 | 1 | 0.5 | 0.25 | <0.03 | 0.03 | 0.125 |
| 5 | 1 | 0.25 | 8 | 0.125 | 0.06 | 0.125 | ||
| 20 | 2 | 1 | 16 | 0.5 | 0.25 | 0.25 | ||
| 40 | 2 | 1 | >64 | 0.5 | 0.25 | 0.25 | ||
| SA0736 | DAP | 0 | 0.5 | 0.25 | 0.25 | <0.03 | 0.03 | 0.125 |
| 5 | 0.5 | 0.25 | 1 | <0.03 | 0.03 | 0.125 | ||
| 20 | 4 | 8 | 32 | 2 | 0.25 | 0.125 | ||
| 40W | 4 | 8 | >64 | 4 | 0.5 | 0.125 | ||
| 40Y | 4 | 8 | >64 | 4 | 0.5 | 0.125 | ||
| ATCC 29213 | DAP | 0 | 0.25 | 0.06 | 0.125 | 0.06 | <0.016 | <0.03 |
| 5 | 1 | 0.5 | 0.5 | 0.06 | 0.03 | <0.03 | ||
| 20 | 2 | 2 | 32 | 0.125 | 0.06 | 0.125 | ||
| 40 | 4 | 4 | 64 | 2 | 0.125 | 0.25 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gostev, V.; Kalinogorskaya, O.; Sopova, J.; Sulian, O.; Chulkova, P.; Velizhanina, M.; Tsvetkova, I.; Ageevets, I.; Ageevets, V.; Sidorenko, S. Adaptive Laboratory Evolution of Staphylococcus aureus Resistance to Vancomycin and Daptomycin: Mutation Patterns and Cross-Resistance. Antibiotics 2023, 12, 928. https://doi.org/10.3390/antibiotics12050928
Gostev V, Kalinogorskaya O, Sopova J, Sulian O, Chulkova P, Velizhanina M, Tsvetkova I, Ageevets I, Ageevets V, Sidorenko S. Adaptive Laboratory Evolution of Staphylococcus aureus Resistance to Vancomycin and Daptomycin: Mutation Patterns and Cross-Resistance. Antibiotics. 2023; 12(5):928. https://doi.org/10.3390/antibiotics12050928
Chicago/Turabian StyleGostev, Vladimir, Olga Kalinogorskaya, Julia Sopova, Ofelia Sulian, Polina Chulkova, Maria Velizhanina, Irina Tsvetkova, Irina Ageevets, Vladimir Ageevets, and Sergey Sidorenko. 2023. "Adaptive Laboratory Evolution of Staphylococcus aureus Resistance to Vancomycin and Daptomycin: Mutation Patterns and Cross-Resistance" Antibiotics 12, no. 5: 928. https://doi.org/10.3390/antibiotics12050928