
Characterization of Oral Candida spp. Biofilms in Children and Adults Carriers from Eastern Europe and South America
 , , ,
, , ,  and
and 
Abstract
:1. Introduction
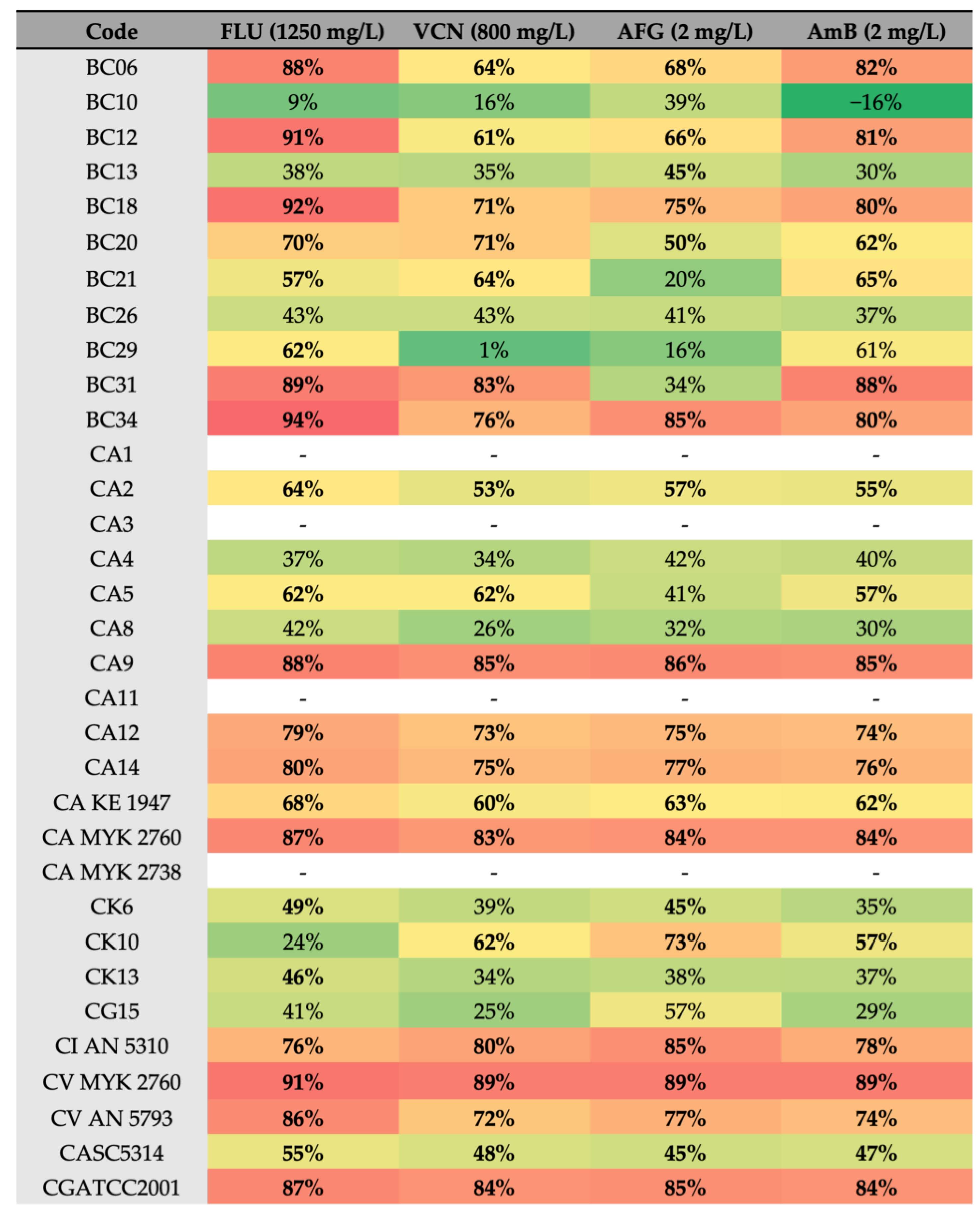
2. Results
2.1. Demographic Data of Study Population and Ethical Aspects
2.2. pBiofilm Formation of Candida spp.
2.3. Biofilm Matrix Composition
2.4. Effect of Antifungals against Biofilm Formation of Different Candida Species
2.5. Confocal Laser Scanning Microscopy
3. Discussion
4. Materials and Methods
4.1. Collection and Identification of the Clinical Isolates
- -
- Children: BC06, BC10, BC12, BC13, BC18, BC20, BC21, BC26, BC29, BC31, and BC34.
- -
- Adults: CA1, CA2, CA4, CA8, CA9, CA11, CA14, CK6, CK10, and CIAN 5310.
- -
- Elders: CA3, CA5, CA12, CAKE 1947, CAMYK 2760, CA MYK 2738, CG15, CVMYK 2760, and CVAN 5793.
4.2. Inoculum Preparation
4.3. Antifungal Drugs
4.4. Biofilm Formation and Crystal Violet Assay
4.5. Quantification of Matrix Polysaccharides
4.6. Quantification of Matrix Protein
4.7. Assessment of the Spatial Arrangement of Candida spp. Cells in Biofilms by CLSM
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Code | Species | O.D.570 | Age | Gender |
|---|---|---|---|---|
| BC06 | C. krusei | 3.34 ± 0.08 | 7 | Female |
| BC10 | C. glabrata | 0.19 ± 0.02 | 4 | Female |
| BC12 | C. krusei | 2.05 ± 0.19 | 3 | Male |
| BC13 | C. krusei | 0.52 ± 0.41 | 6 | Female |
| BC18 | C. krusei | 3.06 ± 0.17 | 2 | Female |
| BC20 | C. krusei | 0.45 ± 0.06 | 3 | Male |
| BC21 | C. glabrata | 0.37 ± 0.08 | 1 | Female |
| BC26 | C. krusei | 0.37 ± 0.10 | 1 | Female |
| BC29 | C. albicans | 0.31 ± 0.02 | 5 | Female |
| BC31 | C. krusei | 2.88 ± 0.39 | 5 | Female |
| BC34 | C. krusei | 3.01 ± 0.41 | 6 | Female |
| CA1 | C. albicans | 0.62 ± 0.17 | 14 | Male |
| CA2 | C. albicans | 1.25 ± 0.36 | 46 | Female |
| CA3 | C. albicans | 0.45 ± 0.12 | 66 | Male |
| CA4 | C. albicans | 1.01± 0.14 | 55 | Male |
| CA5 | C. albicans | 0.92 ± 0.17 | 70 | Male |
| CA8 | C. albicans | 1.00 ± 0.27 | 34 | Male |
| CA9 | C. albicans | 1.06 ± 0.18 | 20 | Female |
| CA11 | C. albicans | 0.68 ± 0.19 | 29 | Female |
| CA12 | C. albicans | 0.91 ± 0.32 | 72 | Female |
| CA14 | C. albicans | 0.94 ± 0.44 | 28 | Male |
| CA KE 1947 | C. albicans | 0.81 ± 0.29 | 66 | Female |
| CA MYK 2760 | C. albicans | 1.37 ± 0.16 | 74 | Male |
| CA MYK 2738 | C. albicans | 0.47± 0.09 | 71 | Male |
| CK6 | C. krusei | 1.13 ± 0.20 | 26 | Male |
| CK10 | C. krusei | 0.82 ± 0.20 | 56 | Male |
| CK13 | C. krusei | 0.65 ± 0.20 | 49 | Male |
| CI AN 5310 | C. intermedia | 1.30 ± 0.09 | 37 | Female |
| CG15 | C. glabrata | 1.44 ± 0.14 | 68 | Male |
| CV MYK 2760 | C. valida | 1.17 ± 0.17 | 78 | Female |
| CV AN 5793 | C. valida | 1.45 ± 0.14 | 70 | Male |
Appendix B
| BC06 | BC10 | BC12 | BC13 | BC18 | BC20 | BC21 | BC26 | BC29 | BC31 | BC34 | CASC 5314 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein (mg/g) | 33.19 ± 4.60 | 35.22 ± 4.70 | 11.01 ± 1.35 | 13.45 ± 1.59 | 19.55 ± 2.55 | 37.63 ± 5.10 | 35.84 ± 3.30 | 95.26 ± 13.27 | 15.05 ± 1.63 | 27.48 ± 3.70 | 13.82 ± 0.70 | 1.24 ± 0.05 |
| Polysaccharides (mg/g) | 19.43 ± 6.07 | 5.55 ± 0.49 | 5.36 ± 0.44 | 21.57 ± 11.11 | 18.34 ± 1.87 | 6.34 ± 0.64 | 173.78 ± 2.26 | 16.96 ± 2.24 | 11.13 ± 5.86 | 6.80 ± 0.61 | 9.79 ± 2.54 | 0.03 ± 0.05 |
| CA2 | CA4 | CA8 | CA9 | CA14 | CK6 | CK10 | CIAN 5310 | CASC 5314 | |
|---|---|---|---|---|---|---|---|---|---|
| Protein (mg/g) | 8.87 ± 1.08 | 46.86 ± 0.58 | 10.52 ± 1.29 | 14.59 ± 1.91 | 11.46 ± 1.44 | 3.16 ± 0.23 | 1.13 ± 0.36 | 2.99 ± 0.24 | 1.24 ± 0.05 |
| Polysaccharides (mg/g) | 5.54 ± 0.74 | 69.13 ± 33.98 | 6.36 ± 0.66 | 7.31 ± 0.90 | 8.34 ± 1.04 | 3.15 ± 2.45 | 2.91 ± 0.26 | 5.70 ± 0.62 | 0.03 ± 0.05 |
| CA5 | CA12 | CAKE 1947 | CAMYK 2760 | CG15 | CVMYK 2760 | CVAN 5793 | CASC 5314 | |
|---|---|---|---|---|---|---|---|---|
| Protein (mg/g) | 1.48 ± 0.44 | 4.43 ± 3.90 | 6.68 ± 0.75 | 0.60 ± 0.39 | 3.02 ± 1.85 | 26.97 ± 3.36 | 16.19 ± 1.91 | 1.24 ± 0.05 |
| Polysaccharides (mg/g) | 5.88 ± 3.45 | 9.28 ± 5.42 | 11.47 ± 1.59 | 1.27 ± 0.66 | 5.22 ± 0.56 | 26.58 ± 14.36 | 4.16 ± 0.07 | 0.03 ± 0.05 |
References
- Rajendra Santosh, A.B.; Muddana, K.; Bakki, S.R. Fungal Infections of Oral Cavity: Diagnosis, Management, and Association with COVID-19. SN Compr. Clin. Med. 2021, 3, 1373–1384. [Google Scholar] [CrossRef] [PubMed]
- de la Rosa-García, E.; Olalde-Hernández, M.J.; Irigoyen-Camacho, M.E.; Mondragón-Padilla, A.; Mendoza-Juache, A.; Sánchez-Vargas, L.O. Antifungal Susceptibility of Oral Isolates of Candida Species from Chronic Kidney Disease Patients on Chronic Dialysis. J. Mycol. Med. 2020, 30, 101009. [Google Scholar] [CrossRef] [PubMed]
- Gallè, F.; Sanguinetti, M.; Colella, G.; Di Onofrio, V.; Torelli, R.; Rossano, F.; Liguori, G. Oral Candidosis: Characterization of a Sample of Recurrent Infections and Study of Resistance Determinants. New Microbiol. 2011, 34, 379–389. [Google Scholar] [PubMed]
- Taylor, M.; Raja, A. Oral Candidiasis (Thrush); StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Fotos, P.G.; Vincent, S.D.; Hellstein, J.W. Oral Candidosis. Oral Surg. Oral Med. Oral Pathol. 1992, 74, 41–49. [Google Scholar] [CrossRef]
- Staniszewska, M. Virulence Factors in Candida Species. Curr. Protein Pept. Sci. 2020, 21, 313–323. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial Biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Rodrigues, M.E.; Henriques, M. Susceptibility of Candida glabrata Biofilms to Echinocandins: Alterations in the Matrix Composition. Biofouling 2018, 34, 569–578. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Rodrigues, M.; Silva, S.; Henriques, M. Candida glabrata Biofilms: How Far Have We Come? J. Fungi 2017, 3, 11. [Google Scholar] [CrossRef]
- Pokhrel, S.; Boonmee, N.; Tulyaprawat, O.; Pharkjaksu, S.; Thaipisutikul, I.; Chairatana, P.; Ngamskulrungroj, P.; Mitrpant, C. Assessment of Biofilm Formation by Candida albicans Strains Isolated from Hemocultures and Their Role in Pathogenesis in the Zebrafish Model. J. Fungi 2022, 8, 1014. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Gonçalves, B.; Rodrigues, M.E.; Silva, S.; Azeredo, J.; Henriques, M. The Effectiveness of Voriconazole in Therapy of Candida glabrata’s Biofilms Oral Infections and Its Influence on the Matrix Composition and Gene Expression. Mycopathologia 2017, 182, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Henriques, M. Portrait of Matrix Gene Expression in Candida glabrata Biofilms with Stress Induced by Different Drugs. Genes 2018, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Correia, A.; Vilanova, M.; Henriques, M. Inflammatory Cell Recruitment in Candida glabrata Biofilm Cell-Infected Mice Receiving Antifungal Chemotherapy. J. Clin. Med. 2019, 8, 142. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Alves, D.F.; Henriques, M. Combination of Posaconazole and Amphotericin b in the Treatment of Candida glabrata Biofilms. Microorganisms 2018, 6, 123. [Google Scholar] [CrossRef]
- Černáková, L.; Líšková, A.; Lengyelová, L.; Rodrigues, C.F. Prevalence and Antifungal Susceptibility Profile of Oral Candida spp. Isolates from a Hospital in Slovakia. Medicina 2022, 58, 576. [Google Scholar] [CrossRef]
- Fornari, G.; Vicente, V.A.; Gomes, R.R.; Muro, M.D.; Pinheiro, R.L.; Ferrari, C.; Herkert, P.F.; Takimura, M.; de Carvalho, N.S.; Queiroz-Telles, F. Susceptibility and Molecular Characterization of Candida Species from Patients with Vulvovaginitis. Braz. J. Microbiol. 2016, 47, 373–380. [Google Scholar] [CrossRef]
- Sucupira, P.H.F.; Moura, T.R.; Gurgel, I.L.S.; Pereira, T.T.P.; Padovan, A.C.B.; Teixeira, M.M.; Bahia, D.; Soriani, F.M. In Vitro and in Vivo Characterization of Host–Pathogen Interactions of the L3881 Candida albicans Clinical Isolate. Front. Microbiol. 2022, 13, 901442. [Google Scholar] [CrossRef]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef]
- Watamoto, T.; Samaranayake, L.P.; Egusa, H.; Yatani, H.; Seneviratne, C.J. Transcriptional Regulation of Drug-Resistance Genes in Candida albicans Biofilms in Response to Antifungals. J. Med. Microbiol. 2011, 60, 1241–1247. [Google Scholar] [CrossRef]
- Steinbach, W.J. Pediatric Invasive Candidiasis: Epidemiology and Diagnosis in Children. J. Fungi 2016, 2, 5. [Google Scholar] [CrossRef]
- Childers, N.K.; Stinnett, E.A.; Wheeler, P.; Wright, J.T.; Castleberry, R.P.; Dasanayake, A.P. Oral Complications in Children with Cancer. Oral Surg. Oral Med. Oral Pathol. 1993, 75, 41–47. [Google Scholar] [CrossRef] [PubMed]
- González Gravina, H.; González De Morán, E.; Zambrano, O.; Chourio, M.L.; Rodríguez De Valero, S.; Robertis, S.; Mesa, L.; Mesa, D.L. Oral Candidiasis in Children and Adolescents with Cancer: Identification of Candida spp. Med. Oral Patol. Oral Cir. Bucal (Int.) 2007, 12, E419–E423. [Google Scholar]
- Shirazi, J.H.; Ali, M.I.; Akhtar, Z.; Jamal, A.; Rashid, A. Pediatric Oropharyngeal Candidiasis: A Comprehensive Study on Risk Factors and Most Prevalent Species of Candida. Pak. J. Pharm. Sci. 2019, 32, 2873–2878. [Google Scholar] [CrossRef] [PubMed]
- Palazzi, D.L.; Arrieta, A.; Castagnola, E.; Halasa, N.; Hubbard, S.; Brozovich, A.A.; Fisher, B.T.; Steinbach, W.J. Candida Speciation, Antifungal Treatment and Adverse Events in Pediatric Invasive Candidiasis: Results from 441 Infections in a Prospective, Multi-National Study. Pediatr. Infect. Dis. J. 2014, 33, 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Rajeshwari, R.; Vyasam, S.; Chandran, J.; Porwal, S.; Ebenezer, K.; Thokchom, M.; James, E.J.; Karuppusami, R. Risk Factors for Candida Infection among Children Admitted to a Pediatric Intensive Care Unit in a Tertiary Care Centre in Southern India. Indian J. Crit. Care Med. 2022, 26, 717–722. [Google Scholar] [CrossRef]
- Steinbach, W.J.; Roilides, E.; Berman, D.; Hoffman, J.A.; Groll, A.H.; Bin-Hussain, I.; Palazzi, D.L.; Castagnola, E.; Halasa, N.; Velegraki, A.; et al. Results From a Prospective, International, Epidemiologic Study of Invasive Candidiasis in Children and Neonates. Pediatr. Infect. Dis. J. 2012, 31, 1252–1257. [Google Scholar] [CrossRef]
- Kim, E.J.; Lee, E.; Kwak, Y.G.; Yoo, H.M.; Choi, J.Y.; Kim, S.R.; Shin, M.J.; Yoo, S.Y.; Cho, N.H.; Choi, Y.H. Trends in the Epidemiology of Candidemia in Intensive Care Units From 2006 to 2017: Results From the Korean National Healthcare-Associated Infections Surveillance System. Front. Med. (Lausanne) 2020, 7, 606976. [Google Scholar] [CrossRef]
- Kumari, A.; Mankotia, S.; Chaubey, B.; Luthra, M.; Singh, R. Role of Biofilm Morphology, Matrix Content and Surface Hydrophobicity in the Biofilm-Forming Capacity of Various Candida Species. J. Med. Microbiol. 2018, 67, 889–892. [Google Scholar] [CrossRef]
- Singh, R.; Kumari, A.; Kaur, K.; Sethi, P.; Chakrabarti, A. Relevance of Antifungal Penetration in Biofilm-Associated Resistance of Candida albicans and Non-albicans Candida Species. J. Med. Microbiol. 2018, 67, 922–926. [Google Scholar] [CrossRef]
- Ferreira, J.A.G.; Carr, J.H.; Starling, C.E.F.; de Resende, M.A.; Donlan, R.M. Biofilm Formation and Effect of Caspofungin on Biofilm Structure of Candida Species Bloodstream Isolates. Antimicrob. Agents Chemother. 2009, 53, 4377–4384. [Google Scholar] [CrossRef]
- Gebremedhin, S.; Dorocka-Bobkowska, B.; Prylinski, M.; Konopka, K.; Duzgunes, N. Miconazole Activity Against Candida Biofilms Developed On Acrylic Discs. J. Physiol. Pharmacol. 2014, 4, 593–600. [Google Scholar]
- Mitchell, K.F.; Zarnowski, R.; Sanchez, H.; Edward, J.A.; Reinicke, E.L.; Nett, J.E.; Mitchell, A.P.; Andes, D.R. Community Participation in Biofilm Matrix Assembly and Function. Proc. Natl. Acad. Sci. USA 2015, 112, 4092–4097. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.E.; Andes, D.R. Contributions of the Biofilm Matrix to Candida Pathogenesis. J. Fungi 2020, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Thiyahuddin, N.M.; Lamping, E.; Rich, A.M.; Cannon, R.D. Yeast Species in the Oral Cavities of Older People: A Comparison between People Living in Their Own Homes and Those in Rest Homes. J. Fungi 2019, 5, 30. [Google Scholar] [CrossRef]
- Rodrigues, C.F. Candida Glabrata Biofilms: Mechanisms of Antifungal Resistance and Matrix Role. Doctoral Thesis, Universidade do Minho, Braga, Portugal, 2018. [Google Scholar]
- Pierce, C.G.; Srinivasan, A.; Uppuluri, P.; Ramasubramanian, A.K.; López-Ribot, J.L. Antifungal Therapy with an Emphasis on Biofilms. Curr. Opin. Pharmacol. 2013, 13, 726–730. [Google Scholar] [CrossRef]
- Sharma, J.; Rosiana, S.; Razzaq, I.; Shapiro, R.S. Linking Cellular Morphogenesis with Antifungal Treatment and Susceptibility in Candida Pathogens. J. Fungi 2019, 5, 17. [Google Scholar] [CrossRef]
- Ha, K.C.; White, T.C. Effects of Azole Antifungal Drugs on the Transition from Yeast Cells to Hyphae in Susceptible and Resistant Isolates of the Pathogenic Yeast Candida albicans. Antimicrob. Agents Chemother. 1999, 43, 763–768. [Google Scholar] [CrossRef]
- Nett, J.; Lincoln, L.; Marchillo, K.; Massey, R.; Holoyda, K.; Hoff, B.; VanHandel, M.; Andes, D. Putative Role of β-1,3 Glucans in Candida albicans Biofilm Resistance. Antimicrob. Agents Chemother. 2007, 51, 510–520. [Google Scholar] [CrossRef]
- Rad, K.K.; Falahati, M.; Roudbary, M.; Farahyar, S.; Nami, S. Overexpression of MDR-1 and CDR-2 genes in fluconazole resistance of Candida albicans isolated from patients with vulvovaginal candidiasis. Curr. Med. Mycol. 2016, 2, 24–29. [Google Scholar] [CrossRef]
- Mitchell, K.F.; Zarnowski, R.; Andes, D.R. Fungal Super Glue: The Biofilm Matrix and Its Composition, Assembly, and Functions. PLoS Pathog. 2016, 12, e1005828. [Google Scholar] [CrossRef]
- Al-Fattani, M.A.; Douglas, L.J. Biofilm Matrix of Candida albicans and Candida tropicalis: Chemical Composition and Role in Drug Resistance. J. Med. Microbiol. 2006, 55, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Douglas, C.M. Fungal ß(1,3)-D-Glucan Synthesis. Med. Mycol. 2001, 39, 55–66. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Boas, D.V.; Haynes, K.; Henriques, M. The MNN2 Gene Knockout Modulates the Antifungal Resistance of Biofilms of Candida glabrata. Biomolecules 2018, 8, 130. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of Fluconazole Resistance in Candida albicans Biofilms: Phase-Specific Role of Efflux Pumps and Membrane Sterols. Infect. Immun. 2003, 71, 4333–4340. [Google Scholar] [CrossRef]
- Fonseca, E.; Silva, S.; Rodrigues, C.F.; Alves, C.T.; Azeredo, J.; Henriques, M. Effects of Fluconazole on Candida Glabrata Biofilms and Its Relationship with ABC Transporter Gene Expression. Biofouling 2014, 30, 447–457. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs for Antifungal Agents Version 10.0, Valid from 2020-02-04. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Clinical_breakpoints/AFST_BP_v10.0_200204_updatd_links_200924.pdf (accessed on 23 February 2023).
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Vande, W.K.; Wickes, B.L.; López-Ribot, J.L. Standardized method for in vitro antifungal susceptibility testing of Candida albicans biofilms. Antimicrob Agents Chemother 2001, 45, 2475–2479, PMCID:PMC90680. [Google Scholar] [CrossRef] [PubMed]
- Delattin, N.; Cammue, B.P.; Thevissen, K. Reactive Oxygen Species-Inducing Antifungal Agents and Their Activity against Fungal Biofilms. Future Med. Chem. 2014, 6, 77–90. [Google Scholar] [CrossRef]
- Radwan, M.A.; AlQuadeib, B.T.; Šiller, L.; Wright, M.C.; Horrocks, B. Oral Administration of Amphotericin B Nanoparticles: Antifungal Activity, Bioavailability and Toxicity in Rats. Drug Deliv. 2017, 24, 40–50. [Google Scholar] [CrossRef]
- Rosato, A.; Piarulli, M.; Schiavone, B.P.I.; Catalano, A.; Carocci, A.; Carrieri, A.; Carone, A.; Caggiano, G.; Franchini, C.; Corbo, F.; et al. In Vitro Effectiveness of Anidulafungin against Candida spp. Biofilms. J. Antibiot. (Tokyo) 2013, 66, 701–704. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, A.M.C.V.; Lopes, B.O.; Leite, A.C.R.d.M.; Cruz, G.S.; Brito, É.H.S.d.; Lima, L.F.d.; Černáková, L.; Azevedo, N.F.; Rodrigues, C.F. Characterization of Oral Candida spp. Biofilms in Children and Adults Carriers from Eastern Europe and South America. Antibiotics 2023, 12, 797. https://doi.org/10.3390/antibiotics12050797
Alves AMCV, Lopes BO, Leite ACRdM, Cruz GS, Brito ÉHSd, Lima LFd, Černáková L, Azevedo NF, Rodrigues CF. Characterization of Oral Candida spp. Biofilms in Children and Adults Carriers from Eastern Europe and South America. Antibiotics. 2023; 12(5):797. https://doi.org/10.3390/antibiotics12050797
Chicago/Turabian StyleAlves, Anelise Maria Costa Vasconcelos, Beatriz Oliveira Lopes, Ana Caroline Rocha de Melo Leite, Gabriela Silva Cruz, Érika Helena Salles de Brito, Laritza Ferreira de Lima, Lucia Černáková, Nuno Filipe Azevedo, and Célia Fortuna Rodrigues. 2023. "Characterization of Oral Candida spp. Biofilms in Children and Adults Carriers from Eastern Europe and South America" Antibiotics 12, no. 5: 797. https://doi.org/10.3390/antibiotics12050797