
Assessment of Prevalence and Diversity of Antimicrobial Resistant Escherichia coli from Retail Meats in Southern California
 and
and
Abstract
:1. Introduction
2. Results
2.1. Risk Factors Associated with the Presence of E. coli in Retail Meat Products
2.2. Phenotypic Antimicrobial Resistance of E. coli from Retail Meat Products
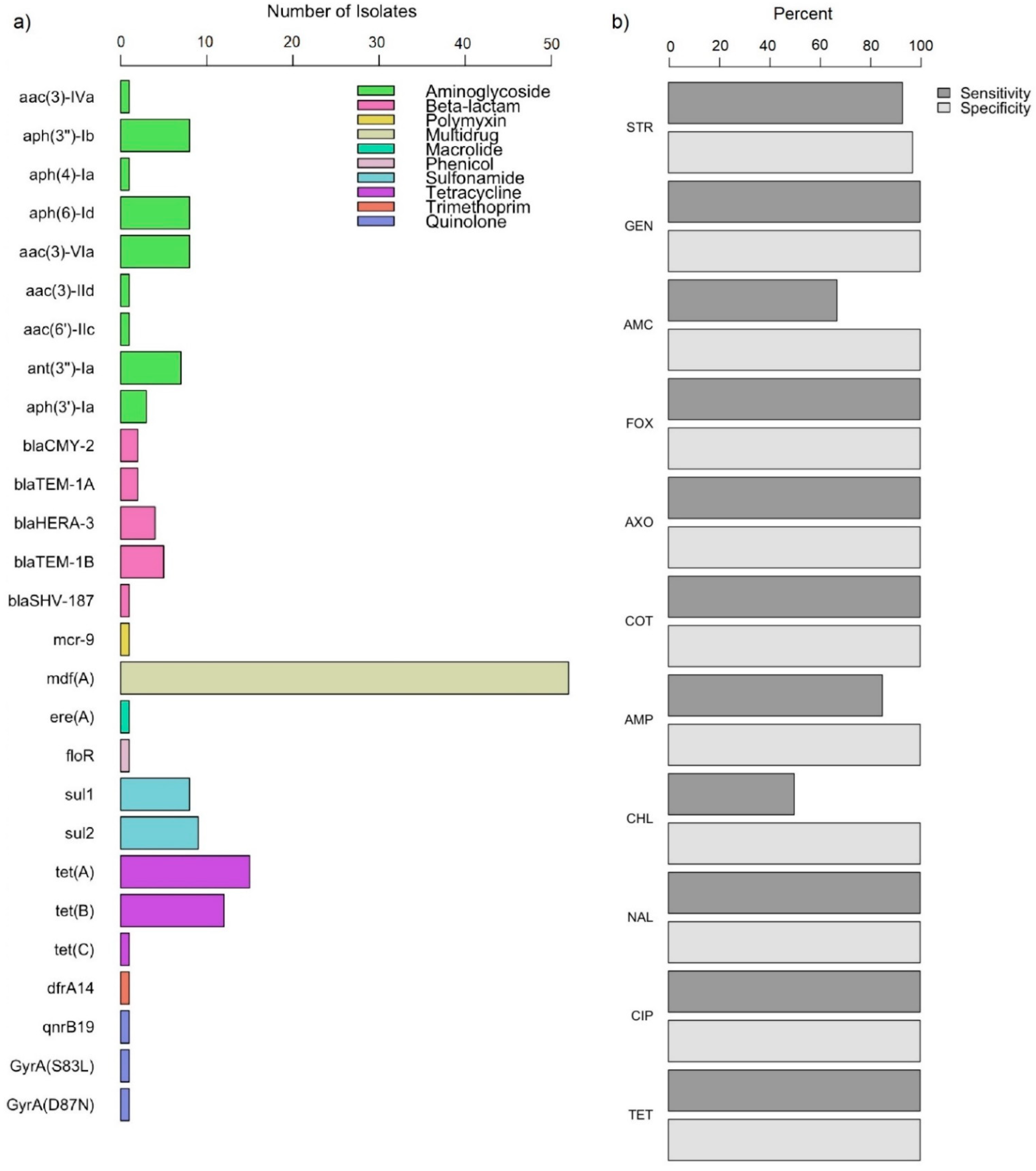
2.3. Genetic Determinants of AMR in E. coli from Retail Meat Products
2.4. Concordance of AMR Phenotype and Genotype
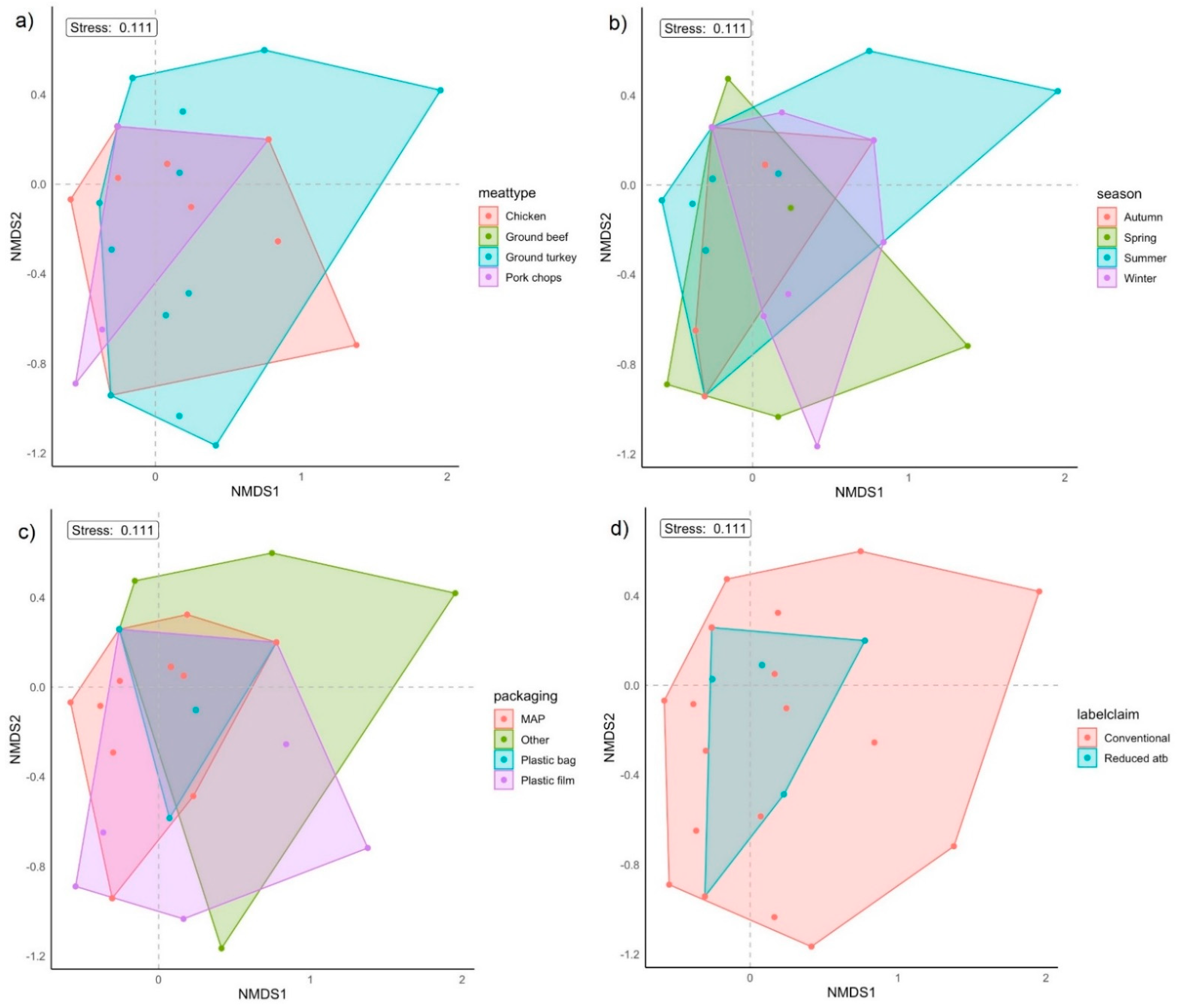
2.5. Clustering Analysis of E. coli Isolates
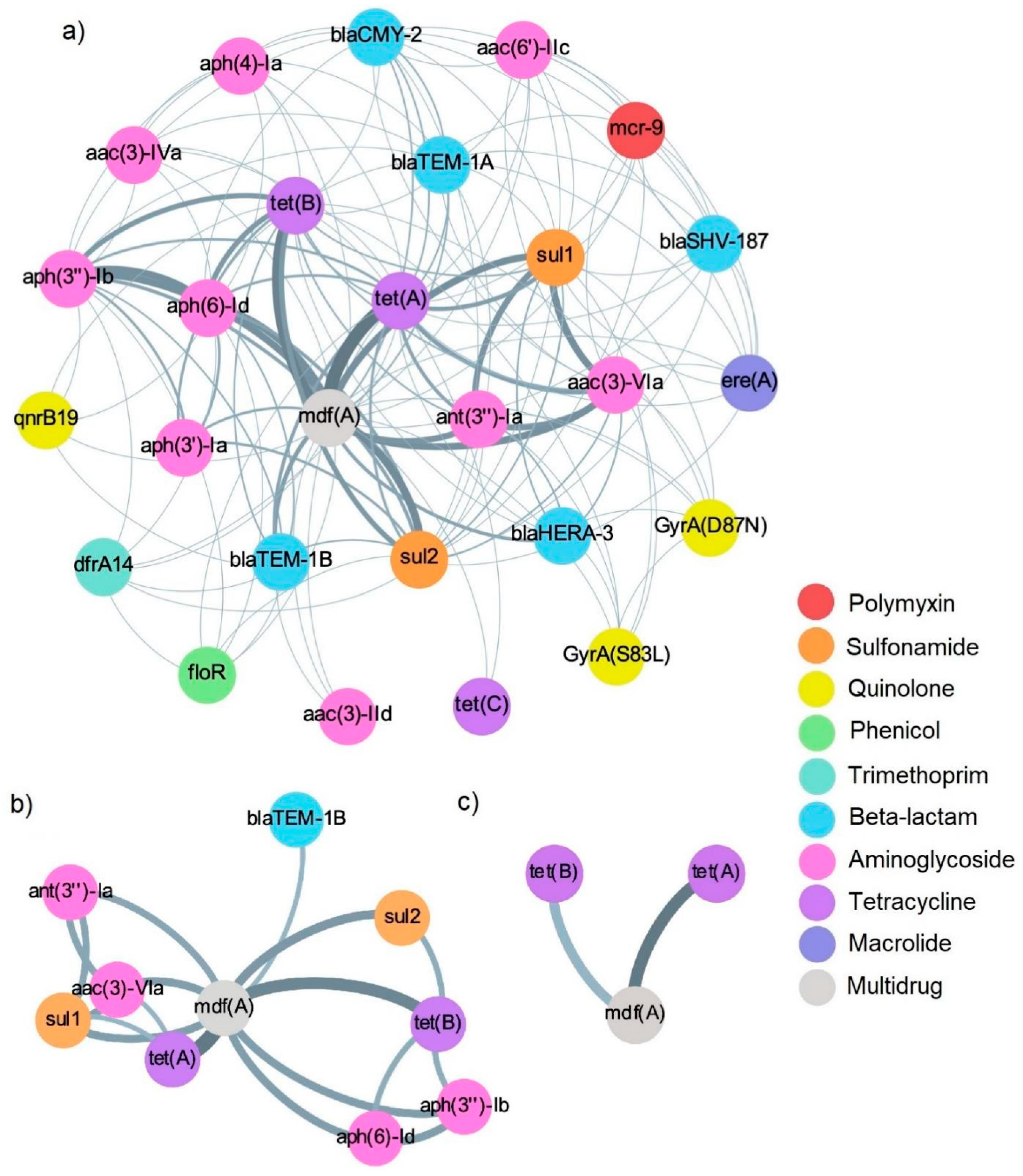
2.6. Co-Occurrence Networks of AMR Genetic Determinants in E. coli Isolates
3. Discussion
4. Materials and Methods
4.1. Study Area and Sampling
4.2. Sample Processing and E. coli Isolation
4.3. Antimicrobial Susceptibility Testing
4.4. Whole-Genome Sequencing and Identification of Resistance Genes
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.G.; Martin, M.J.; Luo, T.L.; Ong, A.C.; Maybank, R.; Corey, B.W.; Harless, C.; Preston, L.N.; Rosado-Mendez, J.A.; Preston, S.B.; et al. A one-year genomic investigation of Escherichia coli epidemiology and nosocomial spread at a large US healthcare network. Genome Med. 2022, 14, 147. [Google Scholar] [CrossRef]
- Yang, X.; Guo, R.; Xie, B.; Lai, Q.; Xu, J.; Hu, N.; Wan, L.; Dai, M.; Zhang, B. Drug resistance of pathogens causing nosocomial infection in orthopedics from 2012 to 2017: A 6-year retrospective study. J. Orthop. Surg. 2021, 16, 100. [Google Scholar] [CrossRef] [PubMed]
- Sikora, A.; Zahra, F. Nosocomial infections. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Emori, T.G.; Gaynes, R.P. An overview of nosocomial infections, including the role of the microbiology laboratory. Clin. Microbiol. Rev. 1993, 6, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Daga, A.P.; Koga, V.L.; Soncini, J.G.M.; De Matos, C.M.; Perugini, M.R.E.; Pelisson, M.; Kobayashi, R.K.T.; Vespero, E.C. Escherichia coli bloodstream infections in patients at a university hospital: Virulence factors and clinical characteristics. Front. Cell. Infect. Microbiol. 2019, 9, 191. [Google Scholar] [CrossRef]
- Rangel, J.; Sparling, P.; Crowe, C.; Griffin, P.; Swerdlow, D. Epidemiology of Escherichia coli O157:H7 outbreaks, United States, 1982–2002. Emerg. Infect. Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef]
- Carstens, C.K.; Salazar, J.K.; Darkoh, C. Multistate outbreaks of foodborne illness in the United States associated with fresh produce from 2010 to 2017. Front. Microbiol. 2019, 10, 2667. [Google Scholar] [CrossRef]
- Luna-Guevara, J.J.; Arenas-Hernandez, M.M.P.; Martínez De La Peña, C.; Silva, J.L.; Luna-Guevara, M.L. The role of pathogenic E. coli in fresh vegetables: Behavior, contamination factors, and preventive measures. Int. J. Microbiol. 2019, 2019, 2894328. [Google Scholar] [CrossRef]
- Olsen, S.J.; Miller, G.; Breuer, T.; Kennedy, M.; Higgins, C.; Walford, J.; Mckee, G.; Fox, K.; Bibb, W.; Mead, P. A waterborne outbreak of Escherichia coli O157:H7 infections and hemolytic uremic syndrome: Implications for rural water systems. Emerg. Infect. Dis. 2002, 8, 370–375. [Google Scholar] [CrossRef]
- Craun, G.F.; Brunkard, J.M.; Yoder, J.S.; Roberts, V.A.; Carpenter, J.; Wade, T.; Calderon, R.L.; Roberts, J.M.; Beach, M.J.; Roy, S.L. Causes of outbreaks associated with drinking water in the United States from 1971 to 2006. Clin. Microbiol. Rev. 2010, 23, 507–528. [Google Scholar] [CrossRef]
- Lim, J.Y.; Yoon, J.W.; Hovde, C.J. A brief overview of Escherichia coli O157:H7 and its plasmid O157. J. Microbiol. Biotechnol. 2010, 20, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, D.A.; Zhao, S.; Tong, E.; Ayers, S.; Singh, A.; Bartholomew, M.J.; McDermott, P.F. Antimicrobial drug resistance in Escherichia coli from humans and food animals, United States, 1950–2002. Emerg. Infect. Dis. 2012, 18, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B.; Bonnie, M. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef] [PubMed]
- Pires, J.; Huisman, J.S.; Bonhoeffer, S.; Van Boeckel, T.P. Increase in antimicrobial resistance in Escherichia coli in food animals between 1980 and 2018 assessed using genomes from public databases. J. Antimicrob. Chemother. 2022, 77, 646–655. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, M.C.; McEwen, S.A.; Pearl, D.L.; Lyytikäinen, O.; Jacobsson, G.; Collignon, P.; Gregson, D.B.; Valiquette, L.; Laupland, K.B. Increasing incidence and antimicrobial resistance in Escherichia coli bloodstream infections: A multinational population-based cohort study. Antimicrob. Resist. Infect. Control 2021, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Dobrindt, U.; Agerer, F.; Michaelis, K.; Janka, A.; Buchrieser, C.; Samuelson, M.; Svanborg, C.; Gottschalk, G.; Karch, H.; Hacker, J. Analysis of genome plasticity in pathogenic and commensal Escherichia coli isolates by use of DNA arrays. J. Bacteriol. 2003, 185, 1831–1840. [Google Scholar] [CrossRef]
- Braz, V.S.; Melchior, K.; Moreira, C.G. Escherichia coli as a multifaceted pathogenic and versatile bacterium. Front. Cell. Infect. Microbiol. 2020, 10, 548492. [Google Scholar] [CrossRef]
- Anjum, M.F.; Schmitt, H.; Börjesson, S.; Berendonk, T.U.; Donner, E.; Stehling, E.G.; Boerlin, P.; Topp, E.; Jardine, C.; Li, X.; et al. The potential of using E. coli as an indicator for the surveillance of antimicrobial resistance (AMR) in the environment. Curr. Opin. Microbiol. 2021, 64, 152–158. [Google Scholar] [CrossRef]
- Stubberfield, E.; AbuOun, M.; Sayers, E.; O’Connor, H.M.; Card, R.M.; Anjum, M.F. Use of whole genome sequencing of commensal Escherichia coli in pigs for antimicrobial resistance surveillance, United Kingdom, 2018. Euro Surveill. Bull. Eur. Mal. Transm. Eur. Commun. Dis. Bull. 2019, 24, 1900136. [Google Scholar] [CrossRef]
- Karp, B.E.; Tate, H.; Plumblee, J.R.; Dessai, U.; Whichard, J.M.; Thacker, E.L.; Hale, K.R.; Wilson, W.; Friedman, C.R.; Griffin, P.M.; et al. National Antimicrobial Resistance Monitoring System: Two Decades of advancing Public Health through Integrated Surveillance of Antimicrobial Resistance. Foodborne Pathog. Dis. 2017, 14, 545–557. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Dadgostar, P. Antimicrobial resistance: Implications and costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Tilocca, B.; Roncada, P. Antimicrobial resistance in veterinary medicine: An overview. Int. J. Mol. Sci. 2020, 21, 1914. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, V.; Kasimanickam, M.; Kasimanickam, R. Antibiotics use in food animal production: Escalation of antimicrobial resistance: Where are we now in combating AMR? Med. Sci. 2021, 9, 14. [Google Scholar] [CrossRef]
- Zhao, S.; Blickenstaff, K.; Bodeis-Jones, S.; Gaines, S.A.; Tong, E.; McDermott, P.F. Comparison of the prevalences and antimicrobial resistances of Escherichia coli isolates from different retail meats in the United States, 2002 to 2008. Appl. Environ. Microbiol. 2012, 78, 1701–1707. [Google Scholar] [CrossRef] [PubMed]
- Economou, V.; Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 2015, 8, 49–61. [Google Scholar] [CrossRef]
- Omer, M.K.; Álvarez-Ordoñez, A.; Prieto, M.; Skjerve, E.; Asehun, T.; Alvseike, O.A. A systematic review of bacterial foodborne outbreaks related to red meat and meat products. Foodborne Pathog. Dis. 2018, 15, 598–611. [Google Scholar] [CrossRef]
- Bryan, F.L. Foodborne diseases in the United States associated with meat and poultry. J. Food Prot. 1980, 43, 140–150. [Google Scholar] [CrossRef]
- Zawack, K.; Li, M.; Booth, J.G.; Love, W.; Lanzas, C.; Gröhn, Y.T. Monitoring antimicrobial resistance in the food supply chain and its implications for FDA policy initiatives. Antimicrob. Agents Chemother. 2016, 60, 5302–5311. [Google Scholar] [CrossRef]
- Food and Drug Administration. The National Antimicrobial Resistance Monitoring System. Available online: https://www.fda.gov/animal-veterinary/antimicrobial-resistance/national-antimicrobial-resistance-monitoring-system (accessed on 6 February 2023).
- Nyirabahizi, E.; Tyson, G.H.; Tate, H.; Kabera, C.; Crarey, E.; Ayers, S.; Strain, E. Northeastern U.S. Salmonella strains from retail meat are more prevalent and more resistant to antimicrobials. J. Food Prot. 2020, 83, 849–857. [Google Scholar] [CrossRef]
- Aronin, S.I.; Gupta, V.; Dunne, M.W.; Watts, J.A.; Yu, K.C. Regional differences in antibiotic-resistant Enterobacterales urine isolates in the United States: 2018–2020. Int. J. Infect. Dis. 2022, 119, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Frost, I.; Van Boeckel, T.P.; Pires, J.; Craig, J.; Laxminarayan, R. Global geographic trends in antimicrobial resistance: The role of international travel. J. Travel Med. 2019, 26, taz036. [Google Scholar] [CrossRef]
- Strachan, N.J.C.; Rotariu, O.; Lopes, B.; Macrae, M.; Fairley, S.; Laing, C.; Gannon, V.; Allison, L.J.; Hanson, M.F.; Dallman, T.; et al. Whole genome sequencing demonstrates that geographic variation of Escherichia coli O157 genotypes dominates host association. Sci. Rep. 2015, 5, 14145. [Google Scholar] [CrossRef] [PubMed]
- Achtman, M. Evolution, population structure, and phylogeography of genetically monomorphic bacterial pathogens. Annu. Rev. Microbiol. 2008, 62, 53–70. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, S. Understanding the contribution of environmental factors in the spread of antimicrobial resistance. Environ. Health Prev. Med. 2015, 20, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Walsh, D.C.I. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Food and Drug Administration. 2018 NARMS Update: Integrated Report Summary Interactive Version. Available online: https://www.fda.gov/animal-veterinary/national-antimicrobial-resistance-monitoring-system/2018-narms-update-integrated-report-summary-interactive-version (accessed on 6 February 2023).
- Zhao, C.; Ge, B.; De Villena, J.; Sudler, R.; Yeh, E.; Zhao, S.; White, D.G.; Wagner, D.; Meng, J. Prevalence of Campylobacter spp., Escherichia coli, and Salmonella serovars in retail chicken, turkey, pork, and beef from the Greater Washington, D.C., area. Appl. Environ. Microbiol. 2001, 67, 5431–5436. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Yang, L.; Wu, X.; Wu, Y.; Shao, B. Prevalence of Escherichia coli and antibiotic resistance in animal-derived food samples—six districts, Beijing, China, 2020. China CDC Wkly. 2021, 3, 999–1004. [Google Scholar] [CrossRef]
- Sheikh, A.A.; Checkley, S.; Avery, B.; Chalmers, G.; Bohaychuk, V.; Boerlin, P.; Reid-Smith, R.; Aslam, M. Antimicrobial resistance and resistance genes in Escherichia coli isolated from retail meat Purchased in Alberta, Canada. Foodborne Pathog. Dis. 2012, 9, 625–631. [Google Scholar] [CrossRef]
- Farhoumand, P.; Hassanzadazar, H.; Soleiman Soltanpour, M.; Aminzare, M.; Abbasi, Z. Prevalence, genotyping and antibiotic resistance of Listeria monocytogenes and Escherichia coli in fresh beef and chicken meats marketed in Zanjan, Iran. Iran. J. Microbiol. 2020, 12, 537–546. [Google Scholar] [CrossRef]
- Van Elsas, J.D.; Semenov, A.V.; Costa, R.; Trevors, J.T. Survival of Escherichia coli in the environment: Fundamental and public health aspects. ISME J. 2011, 5, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.A.; Meadows, S.; Meyers, R.; Parmley, E.J.; Fazil, A. Seasonality and zoonotic foodborne pathogens in Canada: Relationships between climate and Campylobacter, E. coli and Salmonella in meat products. Epidemiol. Infect. 2019, 147, E190. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration. NARMS Now: Integrated Data. Available online: https://www.fda.gov/animal-veterinary/national-antimicrobial-resistance-monitoring-system/narms-now-integrated-data (accessed on 6 February 2023).
- Davis, G.S.; Waits, K.; Nordstrom, L.; Grande, H.; Weaver, B.; Papp, K.; Horwinski, J.; Koch, B.; Hungate, B.A.; Liu, C.M.; et al. Antibiotic-resistant Escherichia coli from retail poultry meat with different antibiotic use claims. BMC Microbiol. 2018, 18, 174. [Google Scholar] [CrossRef]
- Hussain, A.; Shaik, S.; Ranjan, A.; Nandanwar, N.; Tiwari, S.K.; Majid, M.; Baddam, R.; Qureshi, I.A.; Semmler, T.; Wieler, L.H.; et al. Risk of transmission of antimicrobial resistant Escherichia coli from commercial broiler and free-range retail chicken in India. Front. Microbiol. 2017, 8, 2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H.; Kim, Y.; Kim, M.; Kwak, H.; Ryu, S. Whole-genome sequencing-based characteristics in extended-spectrum beta-lactamase-producing Escherichia coli isolated from retail meats in Korea. Microorganisms 2020, 8, 508. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H.; Kim, Y.; Kim, M.; Kwak, H.; Ryu, S. Antimicrobial resistance of Escherichia coli from retail poultry meats in Korea. J. Food Prot. 2020, 83, 1673–1678. [Google Scholar] [CrossRef]
- Food and Drug Administration. Withdrawal of Enrofloxacin for Poultry. Available online: https://www.fda.gov/animal-veterinary/recalls-withdrawals/withdrawal-enrofloxacin-poultry (accessed on 6 February 2023).
- Food and Drug Administration. Extralabel Use and Antimicrobials. Available online: https://www.fda.gov/animal-veterinary/antimicrobial-resistance/extralabel-use-and-antimicrobials (accessed on 6 February 2023).
- Kieffer, N.; Royer, G.; Decousser, J.W.; Bourrel, A.S.; Palmieri, M.; De La Rosa, J.M.O.; Jacquier, H.; Denamur, E.; Nordmann, P.; Poirel, L. Mcr-9, an inducible gene encoding an acquired phosphoethanolamine transferase in Escherichia coli, and its origin. Antimicrob. Agents Chemother. 2019, 63, e00965-19. [Google Scholar] [CrossRef]
- Tyson, G.H.; Li, C.; Hsu, C.H.; Ayers, S.; Borenstein, S.; Mukherjee, S.; Tran, T.T.; McDermot, P.F.; Zhao, S. The mcr-9 gene of Salmonella and Escherichia coli is not associated with colistin resistance in the United States. Antimicrob. Agents Chemother. 2020, 64, e00573-20. [Google Scholar] [CrossRef]
- El-Sayed Ahmed, M.A.E.G.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and its role in the era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef]
- Andrade, F.F.; Silva, D.; Rodrigues, A.; Pina-Vaz, C. Colistin update on its mechanism of action and resistance, present and future challenges. Microorganisms 2020, 8, 1716. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism mcr-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Gogry, F.A.; Siddiqui, M.T.; Sultan, I.; Haq, Q.M.R. Current update on intrinsic and acquired colistin resistance mechanisms in bacteria. Front. Med. 2021, 8, 677720. [Google Scholar] [CrossRef]
- Li, Y.; Dai, X.; Zeng, J.; Gao, Y.; Zhang, Z.; Zhang, L. Characterization of the global distribution and diversified plasmid reservoirs of the colistin resistance gene mcr-9. Sci. Rep. 2020, 10, 8113. [Google Scholar] [CrossRef] [PubMed]
- Kananizadeh, P.; Oshiro, S.; Watanabe, S.; Iwata, S.; Kuwahara-Arai, K.; Shimojima, M.; Ogawa, M.; Tada, T.; Kirikae, T. Emergence of carbapenem-resistant and colistin-susceptible Enterobacter cloacae complex co-harboring blaIMP-1 and mcr-9 in Japan. BMC Infect. Dis. 2020, 20, 282. [Google Scholar] [CrossRef]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of novel mobilized colistin resistance gene mcr-9 in a multidrug-resistant, colistin-susceptible Salmonella enterica serotype Typhimurium isolate. ASM J. 2019, 10, e00853-19. [Google Scholar] [CrossRef]
- Hooper, D.C.; Jacoby, G.A. Topoisomerase inhibitors: Fluoroquinolone mechanisms of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025320. [Google Scholar] [CrossRef] [PubMed]
- Juraschek, K.; Malekzadah, J.; Malorny, B.; Käsbohrer, A.; Schwarz, S.; Meemken, D.; Hammerl, J.A. Characterization of qnrB-carrying plasmids from ESBL- and non-ESBL-producing Escherichia coli. BMC Genom. 2022, 23, 365. [Google Scholar] [CrossRef]
- Ríos, E.; Rodríguez-Avial, I.; Rodríguez-Avial, C.; Hernandez, E.; Picazo, J.J. High percentage of resistance to ciprofloxacin and qnrB19 gene identified in urinary isolates of extended-spectrum β-lactamase-producing Escherichia coli in Madrid, Spain. Diagn. Microbiol. Infect. Dis. 2010, 67, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Coppola, N.; Freire, B.; Umpiérrez, A.; Cordeiro, N.F.; Ávila, P.; Trenchi, G.; Castro, G.; Casaux, M.L.; Fraga, M.; Zunino, P.; et al. Transferable resistance to highest priority critically important antibiotics for human health in Escherichia coli strains obtained from livestockfeces in Uruguay. Front. Vet. Sci. 2020, 7, 588919. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Z.; Zhang, Y.; Zhang, Z.; Lei, L.; Xia, Z. Increasing prevalence of ESBL-producing multidrug resistance Escherichia coli from diseased pets in Beijing, China from 2012 to 2017. Front. Microbiol. 2019, 10, 2852. [Google Scholar] [CrossRef]
- Tyson, G.H.; McDermott, P.F.; Li, C.; Chen, Y.; Tadesse, D.A.; Mukherjee, S.; Bodeis-Jones, S.; Kabera, C.; Gaines, S.A.; Loneragan, G.H.; et al. WGS accurately predicts antimicrobial resistance in Escherichia coli. J. Antimicrob. Chemother. 2015, 70, 2763–2769. [Google Scholar] [CrossRef]
- Jeamsripong, S.; Li, X.; Aly, S.S.; Su, Z.; Pereira, R.V.; Atwill, E.R. Antibiotic resistance genes and associated phenotypes in Escherichia coli and Enterococcus from cattle at different production stages on a dairy farm in central California. Antibiotics 2021, 10, 1042. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Atwill, E.R.; Pitesky, M.; Huang, A.; Lavelle, K.; Rickard, M.; Shafii, M.; Hung-Fan, M.; Li, X. Antimicrobial resistance profiles of non-typhoidal Salmonella from retail meat products in California, 2018. Front. Microbiol. 2022, 13, 835699. [Google Scholar] [CrossRef]
- McDermott, P.F.; Tyson, G.H.; Kabera, C.; Chen, Y.; Li, C.; Folster, J.P.; Ayers, S.L.; Lam, C.; Tate, H.P.; Zhao, S. Whole-genome sequencing for detecting antimicrobial resistance in nontyphoidal Salmonella. Antimicrob. Agents Chemother. 2016, 60, 5515–5520. [Google Scholar] [CrossRef]
- Neuert, S.; Nair, S.; Day, M.R.; Doumith, M.; Ashton, P.M.; Mellor, K.C.; Jenkins, C.; Hopkins, K.L.; Woodford, N.; de Pinna, E.; et al. Prediction of phenotypic antimicrobial resistance profiles from whole genome sequences of non-typhoidal Salmonella enterica. Front. Microbiol. 2018, 9, 592. [Google Scholar] [CrossRef]
- Carroll, L.M.; Buehler, A.J.; Gaballa, A.; Siler, J.D.; Cummings, K.J.; Cheng, R.A.; Wiedmann, M. Monitoring the microevolution of Salmonella enterica in healthy dairy cattle populations at the individual farm level using whole-genome sequencing. Front. Microbiol. 2021, 12, 763669. [Google Scholar] [CrossRef] [PubMed]
- Arredondo-Alonso, S.; Willems, R.J.; van Schaik, W.; Schürch, A.C. On the (im)possibility of reconstructing plasmids from whole-genome short-read sequencing data. Microb. Genomics 2017, 3. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.H.; Li, C.; Hsu, C.H.; Bodeis-Jones, S.; McDermott, P.F. Diverse fluoroquinolone resistance plasmids from retail meat E. coli in the United States. Front. Microbiol. 2019, 10, 2826. [Google Scholar] [CrossRef]
- Long, Y.; Lu, X.; Ni, X.; Liu, J.; Wang, M.; Li, X.; Li, Z.; Zhou, H.; Li, Z.; Wu, K.; et al. High carriage rate of the multiple resistant plasmids harboring quinolone resistance genes in Enterobacter spp. isolated from healthy individuals. Antibiotics 2022, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lei, C.; Cheng, H.; Yang, X.; Huang, Z.; Chen, X.; Ju, Z.; Zhang, H.; Wang, H. Widespread dissemination of plasmid-mediated tigecycline resistance gene tet(X4) in Enterobacterales of porcine origin. Microbiol. Spectr. 2022, 10, e01615-22. [Google Scholar] [CrossRef]
- Melnyk, A.H.; Wong, A.; Kassen, R. The fitness costs of antibiotic resistance mutations. Evol. Appl. 2015, 8, 273–283. [Google Scholar] [CrossRef]
- Schrag, S.; Perrot, V.; Levin, B. Adaptation to the fitness costs of antibiotic resistance in Escherichia coli. Proc. R. Soc. B Biol. Sci. 1997, 264, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration. NARMS Retail Meat Surveillance Laboratory Protocol. Available online: https://www.fda.gov/media/93332/download (accessed on 6 February 2023).
- Food and Drug Administration. The National Antimicrobial Resistance Monitoring System Manual of Laboratory Methods. Available online: https://www.fda.gov/media/101423/download (accessed on 6 February 2023).
- Food and Drug Administration. 2021 NARMS Interpretative Criteria for Susceptibility Testing. Available online: https://www.fda.gov/media/108180/download (accessed on 6 February 2023).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package 2022. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 6 February 2023).
- Anderson, M.J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. Proc. Int. AAAI Conf. Web Soc. Media 2009, 3, 361–362. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Risk Factor | Univariable Models | Multivariable Model | |||
|---|---|---|---|---|---|
| E. coli Positive n/N (%) | OR (95% CI) | p-Value | OR (95% CI) | p-Value | |
| Meat Type | |||||
| Chicken | 38/56 (67.86%) | 8.44 (3.55, 20.09) | <0.0001 | 9.43 (3.84, 23.21) | <0.0001 |
| Ground Turkey | 38/54 (70.37%) | 9.50 (3.93, 22.95) | <0.0001 | 11.00 (4.39, 27.54) | <0.0001 |
| Pork Chop | 18/56 (32.14%) | 1.90 (0.80, 4.51) | 0.15 | 1.93 (0.79, 4.71) | 0.15 |
| Ground Beef | 11/55 (20.00%) | 1.00 | - | 1.00 | - |
| Season | |||||
| Spring | 22/60 (36.67%) | 0.30 (0.13, 0.69) | 0.0046 | 0.22 (0.083, 0.55) | 0.0015 |
| Summer | 28/60 (46.67%) | 0.45 (0.20, 1.03) | 0.0592 | 0.36 (0.14, 0.91) | 0.031 |
| Autumn | 28/60 (46.67%) | 0.45 (0.20, 1.03) | 0.0592 | 0.36 (0.14, 0.91) | 0.031 |
| Winter | 27/41 (65.85%) | 1.00 | - | 1.00 | - |
| Packaging type | |||||
| Modified atmosphere packaging (MAP) | 55/85 (64.71%) | 3.40 (1.55, 7.48) | 0.0023 | - | - |
| Plastic film | 26/64 (40.63%) | 1.27 (0.56, 2.88) | 0.57 | - | - |
| Other (Vacuum, chub, paper) | 10/32 (31.25%) | 0.84 (0.31, 2.27) | 0.74 | - | - |
| Plastic bag | 14/40 (35.00%) | 1.00 | - | - | - |
| Label Claim | |||||
| Reduced antibiotic claim | 27/37 (72.97%) | 3.67 (1.68, 8.02) | 0.0011 | - | - |
| Conventional | 78/184 (42.39%) | 1.00 | - | - | - |
| Presence of Salmonella | |||||
| Yes | 8/11 (72.73%) | 3.11 (0.80, 12.04) | 0.10 | - | - |
| No | 97/210 (46.19%) | 1.00 | - | - | - |
| Overall prevalence | 105/221 (47.51%) | - | - | - | - |
| Number of Isolates Resistant to Antimicrobial Drugs (%) | ||||||
|---|---|---|---|---|---|---|
| CLSI Class | Antimicrobial Agent | Chicken (n = 38) | Ground Turkey (n = 38) | Pork Chop (n = 18) | Ground Beef (n = 11) | All Samples (n = 105) |
| Aminoglycosides | STR | 10 (26.32%) | 17 (44.74%) | 5 (27.78%) | 0 (0%) | 32 (30.48%) |
| GEN | 9 (23.68%) | 7 (18.42%) | 1 (5.56%) | 0 (0%) | 17 (16.19%) | |
| Beta-lactam combination agents | AMC | 1 (2.63%) | 2 (5.26%) | 0 (0%) | 0 (0%) | 3 (2.86%) |
| Cephems | FOX | 1 (2.63%) | 1 (2.63%) | 0 (0%) | 0 (0%) | 2 (1.90%) |
| AXO | 1 (2.63%) | 1 (2.63%) | 0 (0%) | 0 (0%) | 2 (1.90%) | |
| Folate pathway antagonists | COT | 0 (0%) | 2 (5.26%) | 0 (0%) | 0 (0%) | 2 (1.90%) |
| Macrolides | AZI | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Penems | MER | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Penicillins | AMP | 5 (13.16%) | 14 (36.84%) | 3 (16.67%) | 0 (0%) | 22 (20.95%) |
| Phenicols | CHL | 0 (0%) | 2 (5.26%) | 0 (0%) | 0 (0%) | 2 (1.90%) |
| Quinolones | NAL | 0 (0%) | 1 (2.63%) | 1 (5.56%) | 0 (0%) | 2 (1.90%) |
| CIP | 0 (0%) | 1 (2.63%) | 0 (0%) | 0 (0%) | 1 (0.95%) | |
| Tetracycline | TET | 15 (39.47%) | 24 (63.16%) | 7 (38.89%) | 0 (0%) | 46 (43.81%) |
| Meat Type | Antimicrobial Drug (Abbreviation) | |||||||
|---|---|---|---|---|---|---|---|---|
| Ampicillin (AMP) | Gentamicin (GEN) | Streptomycin (STR) | Tetracycline (TET) | |||||
| OR (95% CI) | p-Value | OR (95% CI) | p-Value | OR (95% CI) | p-Value | OR (95% CI) | p-Value | |
| Chicken | 1.31 (0.23, 9.20) | 1.00 | 8.47 (1.05, 394.18) | 0.042 | 1.70 (0.45, 7.27) | 0.56 | 2.03 (0.63, 7.08) | 0.29 |
| Ground turkey | 4.94 (1.17, 30.11) | 0.025 | 6.18 (0.72, 294.92) | 0.13 | 3.81 (1.10, 15.56) | 0.032 | 5.24 (1.65, 18.55) | 0.003 |
| Non-poultry (beef, pork) | 1.00 | - | 1.00 | - | 1.00 | - | 1.00 | - |
| Antibiogram Pattern | Number of Isolates (n/N %) | Multidrug Resistant |
|---|---|---|
| Susceptible to all drugs in MIC panel | 51 (48.57%) | No |
| TET | 12 (11.43%) | No |
| STR-TET | 8 (7.62%) | No |
| AMP-TET | 5 (4.76%) | No |
| GEN-STR-TET | 5 (4.76%) | No |
| GEN-STR | 4 (3.81%) | No |
| AMP | 2 (1.90%) | No |
| AMP-GEN-STR-TET | 2 (1.90%) | Yes |
| AMP-GEN-TET | 2 (1.90%) | Yes |
| AMP-STR-TET | 2 (1.90%) | Yes |
| AMC-AMP-FOX-AXO-GEN-STR-TET | 1 (0.95%) | Yes |
| AMC-AMP-FOX-AXO-GEN-TET | 1 (0.95%) | Yes |
| AMC-AMP-STR-TET | 1 (0.95%) | Yes |
| AMP-CHL-STR-TET | 1 (0.95%) | Yes |
| AMP-CHL-STR-TET-COT | 1 (0.95%) | Yes |
| AMP-GEN-NAL-STR-TET-CIP | 1 (0.95%) | Yes |
| AMP-GEN-STR | 1 (0.95%) | No |
| AMP-NAL-STR-TET-CIPdsc * | 1 (0.95%) | Yes |
| AMP-STR-TET-CIPdsc * | 1 (0.95%) | Yes |
| STR | 1 (0.95%) | No |
| STR-TET-CIPdsc | 1 (0.95%) | No |
| STR-TET-COT | 1 (0.95%) | Yes |
| Grouping Factor | PERMDISP2 p-Value (F) | PERMANOVA p-Value (R2) | ANOSIM p-Value (R) |
|---|---|---|---|
| Meat type | 0.010 (4.25) | 0.10 (0.088) | 0.89 (−0.059) |
| Season | 0.41 (0.97) | 0.73 (0.044) | 0.80 (−0.022) |
| Packaging type | 0.78 (0.36) | 0.55 (0.053) | 0.24 (0.038) |
| Label claim | 0.39 (0.78) | 0.80 (0.0094) | 0.87 (−0.11) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.Y.; Lavelle, K.; Huang, A.; Atwill, E.R.; Pitesky, M.; Li, X. Assessment of Prevalence and Diversity of Antimicrobial Resistant Escherichia coli from Retail Meats in Southern California. Antibiotics 2023, 12, 782. https://doi.org/10.3390/antibiotics12040782
Lee KY, Lavelle K, Huang A, Atwill ER, Pitesky M, Li X. Assessment of Prevalence and Diversity of Antimicrobial Resistant Escherichia coli from Retail Meats in Southern California. Antibiotics. 2023; 12(4):782. https://doi.org/10.3390/antibiotics12040782
Chicago/Turabian StyleLee, Katie Yen, Kurtis Lavelle, Anny Huang, Edward Robert Atwill, Maurice Pitesky, and Xunde Li. 2023. "Assessment of Prevalence and Diversity of Antimicrobial Resistant Escherichia coli from Retail Meats in Southern California" Antibiotics 12, no. 4: 782. https://doi.org/10.3390/antibiotics12040782