
Novel Mutations Conferring Amoxicillin Resistance in Helicobacter pylori in South Korea

Abstract
:1. Introduction
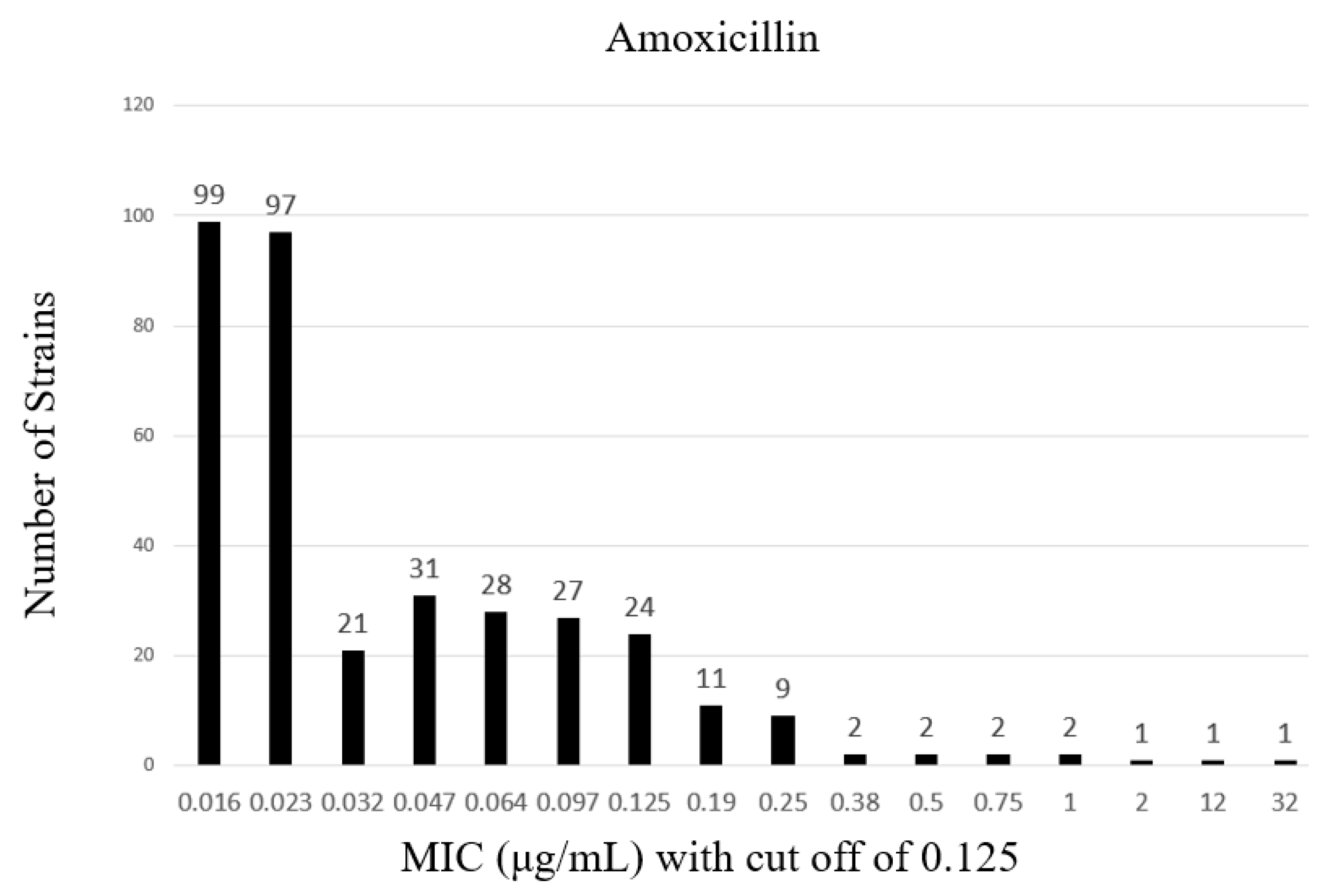
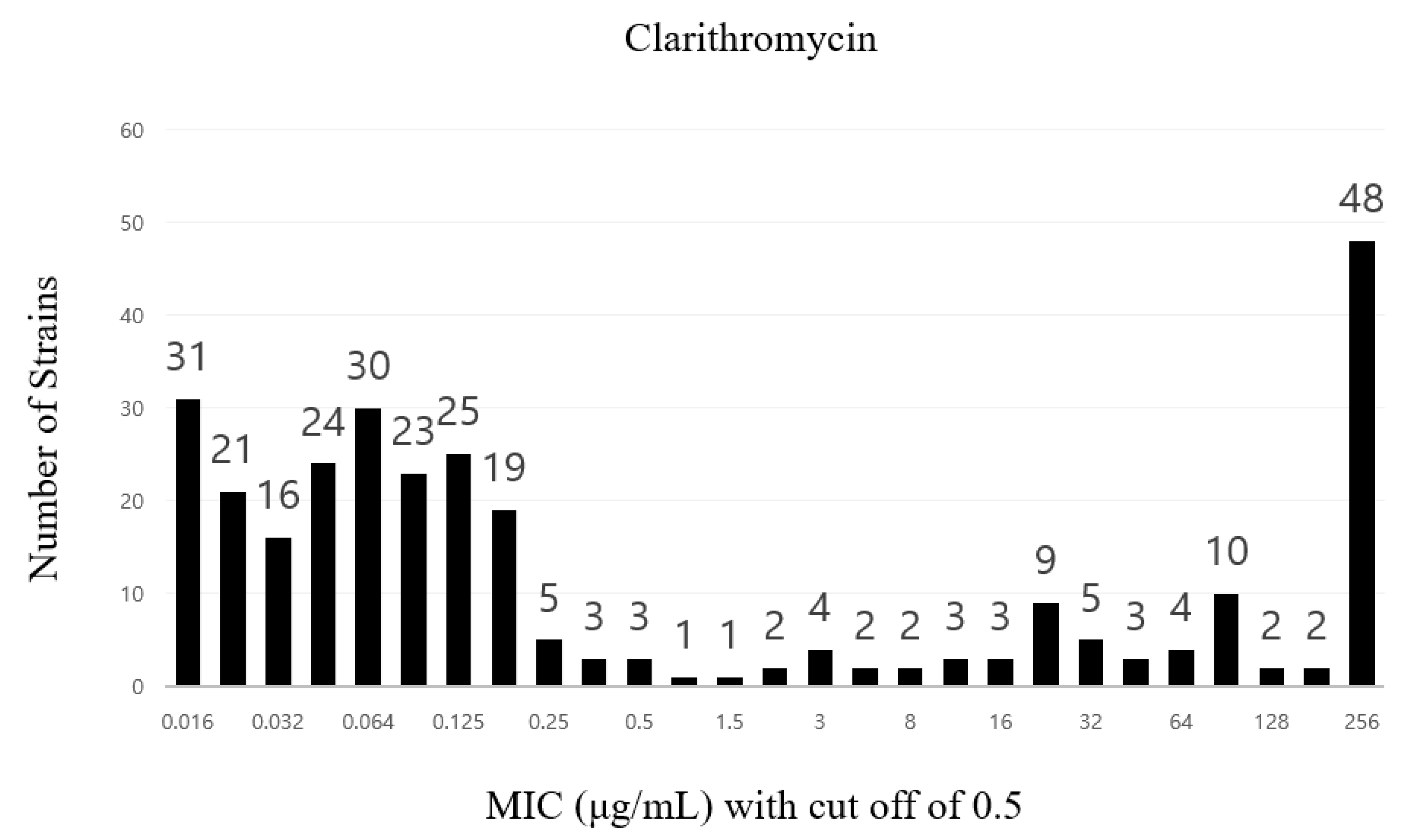
2. Results
3. Discussion
4. Materials and Methods
4.1. H. pylori Isolates and Culture Conditions
4.2. H. pylori Isolates and Culture Conditions
4.3. H. pylori Antibiotic Susceptibility Test
4.4. Whole-Genome Sequencing
4.5. Multilocus Sequencing Typing (MLST)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.Y.; Kim, N. Future trends of Helicobacter pylori eradication therapy in Korea. Korean J. Gastroenterol. 2014, 63, 158–170. [Google Scholar] [CrossRef]
- Lim, S.H.; Kwon, J.W.; Kim, N.; Kim, G.H.; Kang, J.M.; Park, M.J.; Yim, J.Y.; Kim, H.U.; Baik, G.H.; Seo, G.S.; et al. Prevalence and risk factors of Helicobacter pylori infection in Korea: Nationwide multicenter study over 13 years. BMC Gastroenterol. 2013, 13, 104. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.; Jeon, S.W. Changes in the eradication rate of conventional triple therapy for Helicobacter pylori infection in Korea. Korean J. Gastroenterol. 2014, 63, 141–145. [Google Scholar] [CrossRef]
- Park, J.Y.; Dunbar, K.B.; Mitui, M.; Arnold, C.A.; Lam-Himlin, D.M.; Valasek, M.A.; Thung, I.; Okwara, C.; Coss, E.; Cryer, B.; et al. Helicobacter pylori clarithromycin resistance and treatment failure are common in the USA. Dig. Dis. Sci. 2016, 61, 2373–2380. [Google Scholar] [CrossRef]
- Lee, J.H.; Ahn, J.Y.; Choi, K.D.; Jung, H.Y.; Kim, J.M.; Baik, G.H.; Kim, B.W.; Park, J.C.; Jung, H.K.; Cho, S.J.; et al. Nationwide antibiotic resistance mapping of Helicobacter pylori in Korea: A prospective multicenter study. Helicobacter 2019, 24, e12592. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Nguyen, A.T.; Quach, D.T.; Pham, D.T.; Cao, N.M.; Nguyen, U.T.; Dang, A.N.; Tran, M.A.; Quach, L.H.; Tran, K.T.; et al. Emergence of amoxicillin resistance and identification of novel mutations of the pbp1A gene in Helicobacter pylori in Vietnam. BMC Microbiol. 2022, 22, 41. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.E.; Park, M.I.; Park, S.J.; Moon, W.; Choi, Y.J.; Cheon, J.H.; Kwon, H.J.; Ku, K.H.; Yoo, C.H.; Kim, J.H.; et al. Trends in Helicobacter pylori eradication rates by first-line triple therapy and related factors in eradication therapy. Korean J. Intern. Med. 2015, 30, 801–807. [Google Scholar] [CrossRef]
- Seta, T.; Takahashi, Y.; Noguchi, Y.; Shikata, S.; Sakai, T.; Sakai, K.; Yamashita, Y.; Nakayama, T. Effectiveness of Helicobacter pylori eradication in the prevention of primary gastric cancer in healthy asymptomatic people: A systematic review and meta-analysis comparing risk ratio with risk difference. PLoS ONE 2017, 12, e0183321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Shin, J.H.; Roe, I.H.; Sohn, S.G.; Lee, J.H.; Kang, G.H.; Lee, H.K.; Jeong, B.C.; Lee, S.H. Impact of clarithromycin resistance on eradication of Helicobacter pylori in infected adults. Antimicrob. Agents Chemother. 2005, 49, 1600–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global prevalence of Helicobacter pylori infection: Systematic review and meta-analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Gasparetto, M.; Pescarin, M.; Guariso, G. Helicobacter pylori eradication therapy: Current availabilities. ISRN Gastroenterol. 2012, 2012, 186734. [Google Scholar] [CrossRef] [Green Version]
- Posteraro, P.; Branca, G.; Sanguinetti, M.; Ranno, S.; Cammarota, G.; Rahimi, S.; De Carlo, M.; Posteraro, B.; Fadda, G. Rapid detection of clarithromycin resistance in Helicobacter pylori using a PCR-based denaturing HPLC assay. J. Antimicrob. Chemother. 2006, 57, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of antibiotic resistance in Helicobacter pylori: A systematic review and meta-analysis in World Health Organization regions. Gastroenterology 2018, 155, 1372–1382.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.M.; Lee, K.H.; Kim, J.H.; Park, S.Y.; Song, Y.G.; Jeon, S.Y.; Park, H. Is Only Clarithromycin susceptibility important for the successful eradication of Helicobacter pylori? Antibiotics 2020, 9, 589. [Google Scholar] [CrossRef] [PubMed]
- Chey, W.D.; Leontiadis, G.I.; Howden, C.W.; Moss, S.F. ACG clinical guideline: Treatment of Helicobacter pylori infection. Am. J. Gastroenterol. 2017, 112, 212–239. [Google Scholar] [CrossRef]
- Pastukh, N.; Binyamin, D.; On, A.; Paritsky, M.; Peretz, A. Geno Type® HelicoDR test in comparison with histology and culture for Helicobacter pylori detection and identification of resistance mutations to clarithromycin and fluoroquinolones. Helicobacter 2017, 22, e12447. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, N.N.; Gallaher, B.; Schiller, N.L. Evolution of amoxicillin resistance of Helicobacter pylori in vitro: Characterization of resistance mechanisms. Microb. Drug Resist. 2014, 20, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Miftahussurur, M.; Cruz, M.; Subsomwong, P.; Jiménez Abreu, J.A.; Hosking, C.; Nagashima, H.; Akada, J.; Yamaoka, Y. Clarithromycin-based triple therapy is still useful as an initial treatment for Helicobacter pylori infection in the Dominican Republic. Am. J. Trop. Med. Hyg. 2017, 96, 1050–1059. [Google Scholar] [CrossRef] [Green Version]
- Tuan, V.P.; Narith, D.; Tshibangu-Kabamba, E.; Dung, H.D.Q.; Viet, P.T.; Sokomoth, S.; Binh, T.T.; Sokhem, S.; Tri, T.D.; Ngov, S.; et al. A next-generation sequencing-based approach to identify genetic determinants of antibiotic resistance in Cambodian Helicobacter pylori clinical isolates. J. Clin. Med. 2019, 8, 858. [Google Scholar] [CrossRef] [Green Version]
- Mehrotra, T.; Devi, T.B.; Kumar, S.; Talukdar, D.; Karmakar, S.P.; Kothidar, A.; Verma, J.; Kumari, S.; Alexander, S.M.; Retnakumar, R.J.; et al. Antimicrobial resistance and virulence in Helicobacter pylori: Genomic insights. Genomics 2021, 113, 3951–3966. [Google Scholar] [CrossRef]
- Sukri, A.; Hanafiah, A.; Yusoff, H.; Shamsul Nizam, N.A.; Nameyrra, Z.; Wong, Z.; Raja Ali, R.A. Multidrug-Resistant Helicobacter pylori Strains: A Five-Year Surveillance Study and Its Genome Characteristics. Antibiotics 2022, 11, 1391. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Yoshiyama, H.; Nakazawa, T.; Park, I.-D.; Chang, M.-W.; Yanai, H.; Okita, K.; Shirai, M. A change in PBP1 is involved in amoxicillin resistance of clinical isolates of Helicobacter pylori. J. Antimicrob. Chemother. 2002, 50, 849–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westblom, T.U.; Madan, E.; Midkiff, B.R. Egg yolk emulsion agar, a new medium for the cultivation of Helicobacter pylori. J. Clin. Microbiol. 1991, 29, 819–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| No. of Total Clinical Specimens | 1952 | ||
|---|---|---|---|
| No. of H. pylori Identified by MALDI-TOF-MS | 368 | ||
| AST Results Using an E-Test | Antibiotic Type | MIC Breakpoint (mg/L) | No. of Strains |
| Amoxicillin | >0.125 | 31 | |
| Clarithromycin | >0.5 | 101 | |
| Metronidazole | >8 | 68 | |
| Tetracycline | >1 | 20 | |
| Levofloxacin | >1 | 114 |
| Gene 1 | Common SNPs in Nine Amoxicillin-Resistant Strains |
|---|---|
| pbp1a (8) | M17V, F125L, D479E, D535N, S589G, K648Q, R649K, R656P |
| pbp2 (7) | N16T, A26T, M97T, V218I, K240E, K359E, S456H, N |
| nhaC (4) | N53Y, H, I, V70I, G338E, Y450H |
| hofH (9) | S99G, I177V, K183H, N302D, L323R, F381L, T421S, N, Y436H, D452N |
| hofC (16) | A11T, A42S, A92V, G95S, K97R, Q175E, I197V, I206V, K221N, N222K, G230S, V242I, L349F, P382I, V383I, T491N |
| hefC (2) | S316G, I361V |
| H. pylori Strain | MIC90 (ug/mL) a | ST (Strain Type) | Allele | ||||||
|---|---|---|---|---|---|---|---|---|---|
| atpA | epf | mutY | ppa | trpC | ure1 | yphC | |||
| H-13 | <256 | - | 1400 b | 1893 b | 472 c | 1822 b | 426 b | 3013 b | 72 c |
| H-33 | <256 | - | 825 c | 2423 b | 3149 b | 1957 b | 735 c | 44 b | 1932 b |
| H-58 | 8 | - | 2894 b | 1815 b | 1970 b | 426 | 54 b | 54 c | 1971 b |
| H-64 | 16 | - | 1216 b | 2421 b | 1968 c | 1517 | 3015 b | 1979 b | 3037 b |
| H-69 | <256 | - | 2720 b | 2703 b | 1979 c | 64 b | 3236 c | 2096 b | 2678 b |
| H-76 | 2 | - | 2894 b | 1815 b | 1970 b | 426 | 54 b | 54 c | 1971 b |
| H-103 | 48 | - | 2750 c | 1278 b | 493 c | 1066 b | 62 c | 2804 b | 3076 b |
| H-124 | 3 | - | 50 b | 2331 b | 725 b | 2754 b | 50 b | 2804 b | 3295 c |
| H-189 | <256 | - | 2750 c | 499 b | 2360 b | 1125 b | 1526 c | 2804 b | 3076 b |
| H-285 | 192 | - | 3114 b | 1167 b | 452 b | 51 | 2976 | 905 b | 1986 b |
| H-297 | 6 | - | 53 b | 424 b | 3149 b | 168 b | 967 b | 2000 b | 1962 b |
| H-313 | <256 | - | 50 b | 2930 b | 50 b | 64 b | 3254 c | 2829 b | 919 b |
| H-325 | 96 | - | 2753 b | 2714 b | 1355 b | 1917 b | 967 b | 2103 b | 34 c |
| H-15 | 0.016 | - | 1096 b | 1908 b | 725 c | 940 b | 50 b | 424 b | 515 b |
| H-35 | 0.047 | - | 3114 b | 704 b | 3188 b | 1066 b | 1241 b | 883 b | 2076 c |
| H-114 | 0.016 | - | 1870 b | 701 b | 2969 c | 459 b | 50 c | 3254 b | 373 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.Y.; Lee, E.H.; Kim, D.; Song, Y.G.; Jeong, S.J. Novel Mutations Conferring Amoxicillin Resistance in Helicobacter pylori in South Korea. Antibiotics 2023, 12, 748. https://doi.org/10.3390/antibiotics12040748
Park SY, Lee EH, Kim D, Song YG, Jeong SJ. Novel Mutations Conferring Amoxicillin Resistance in Helicobacter pylori in South Korea. Antibiotics. 2023; 12(4):748. https://doi.org/10.3390/antibiotics12040748
Chicago/Turabian StylePark, Soon Young, Eun Hwa Lee, Dokyun Kim, Young Goo Song, and Su Jin Jeong. 2023. "Novel Mutations Conferring Amoxicillin Resistance in Helicobacter pylori in South Korea" Antibiotics 12, no. 4: 748. https://doi.org/10.3390/antibiotics12040748