
Comparative Evaluation of Existing and Rationally Designed Novel Antimicrobial Peptides for Treatment of Skin and Soft Tissue Infections
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Peptides
2.2. Antibacterial Activity
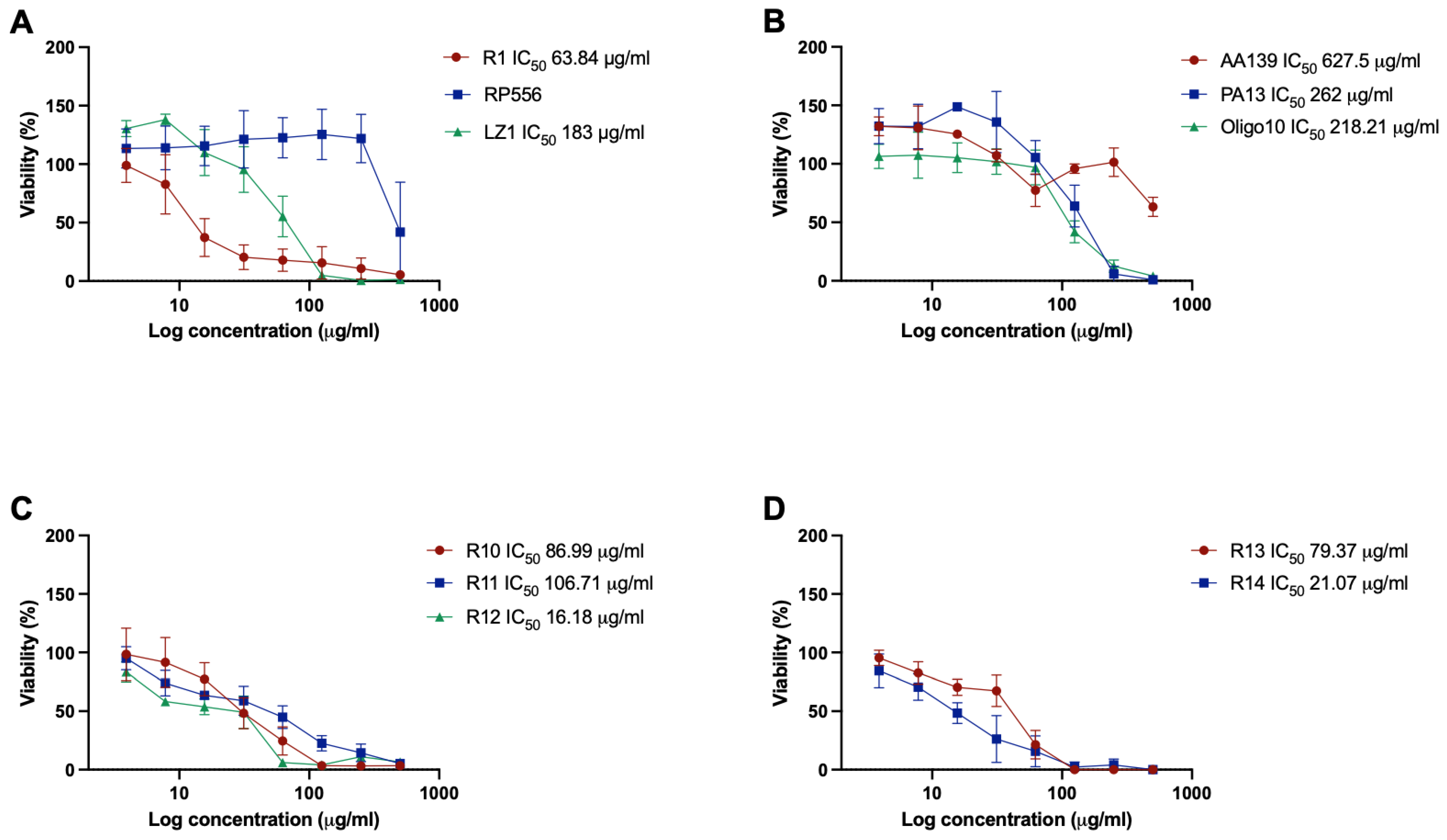
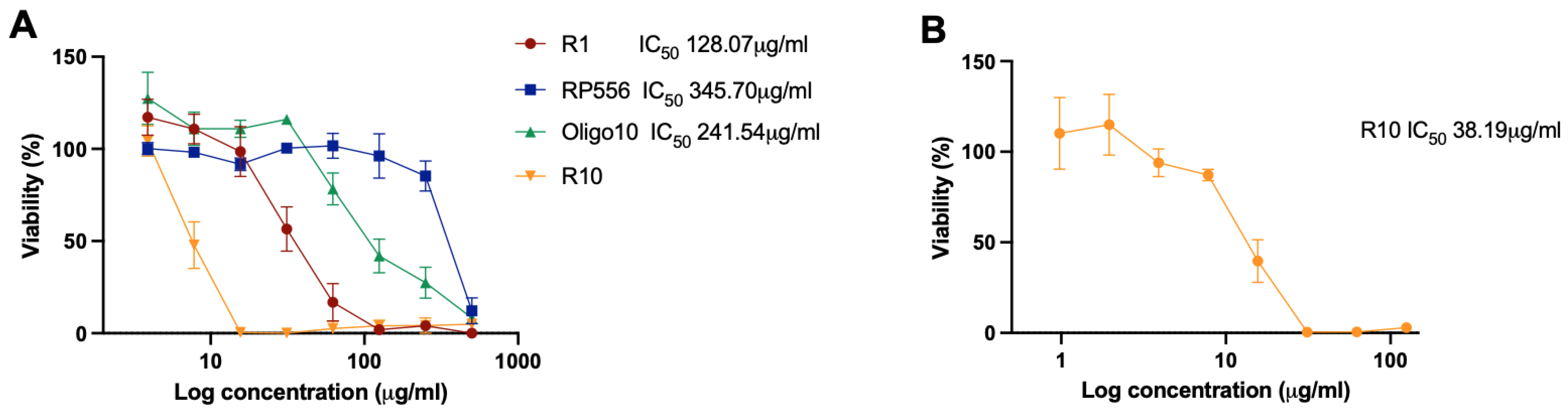
2.3. Cytotoxicity
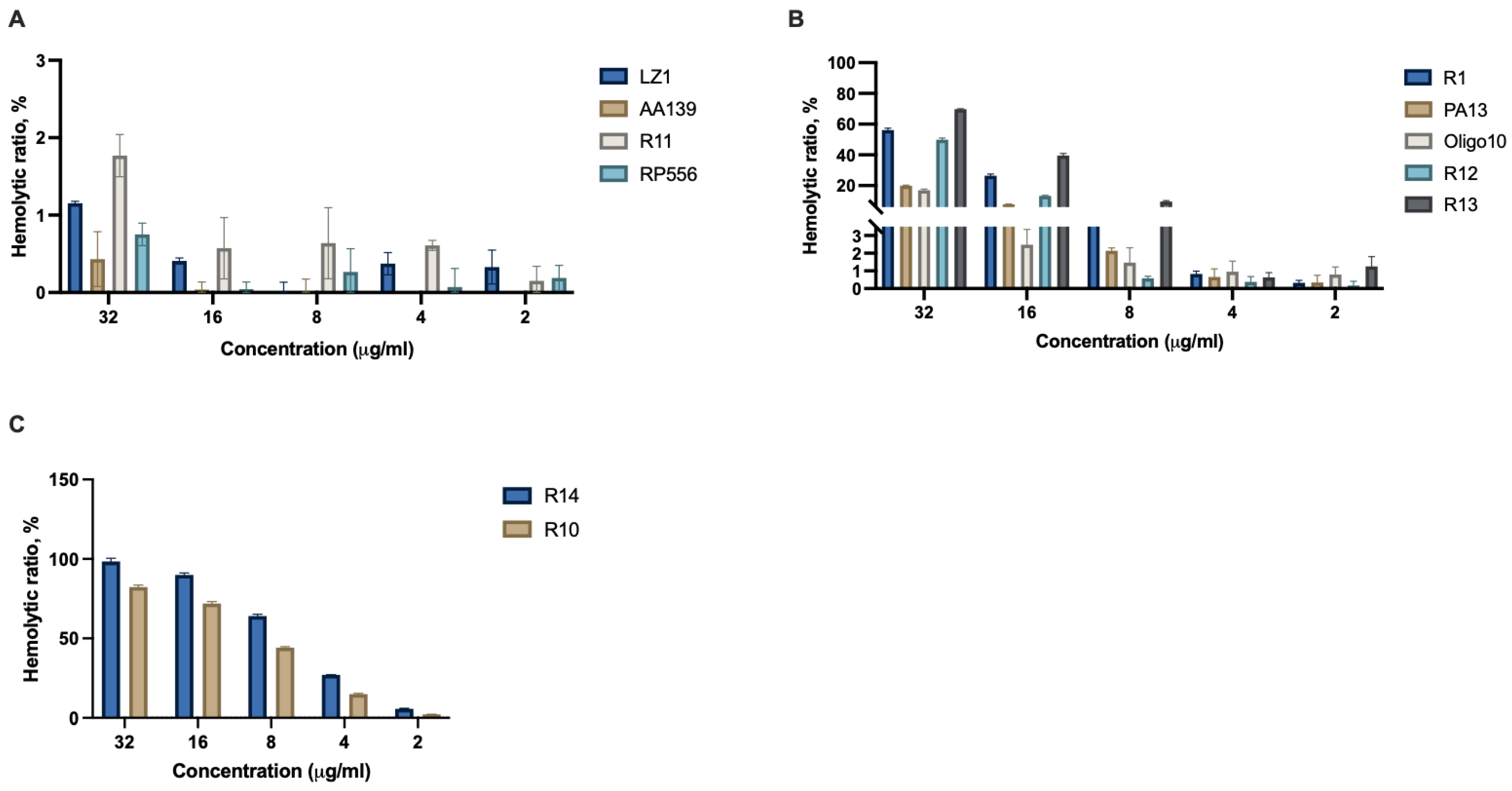
2.4. Hemolysis
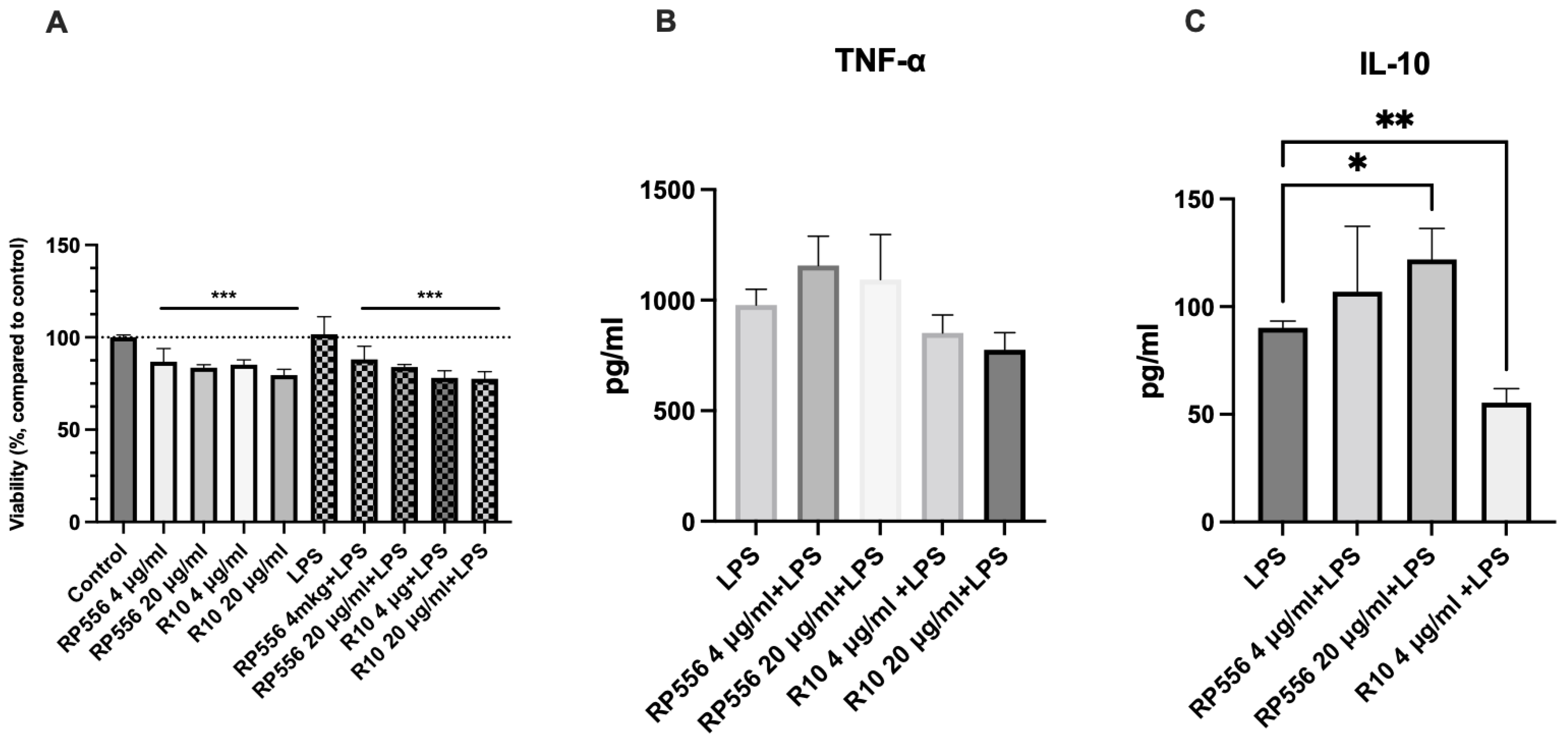
2.5. Immunomodulatory Activity
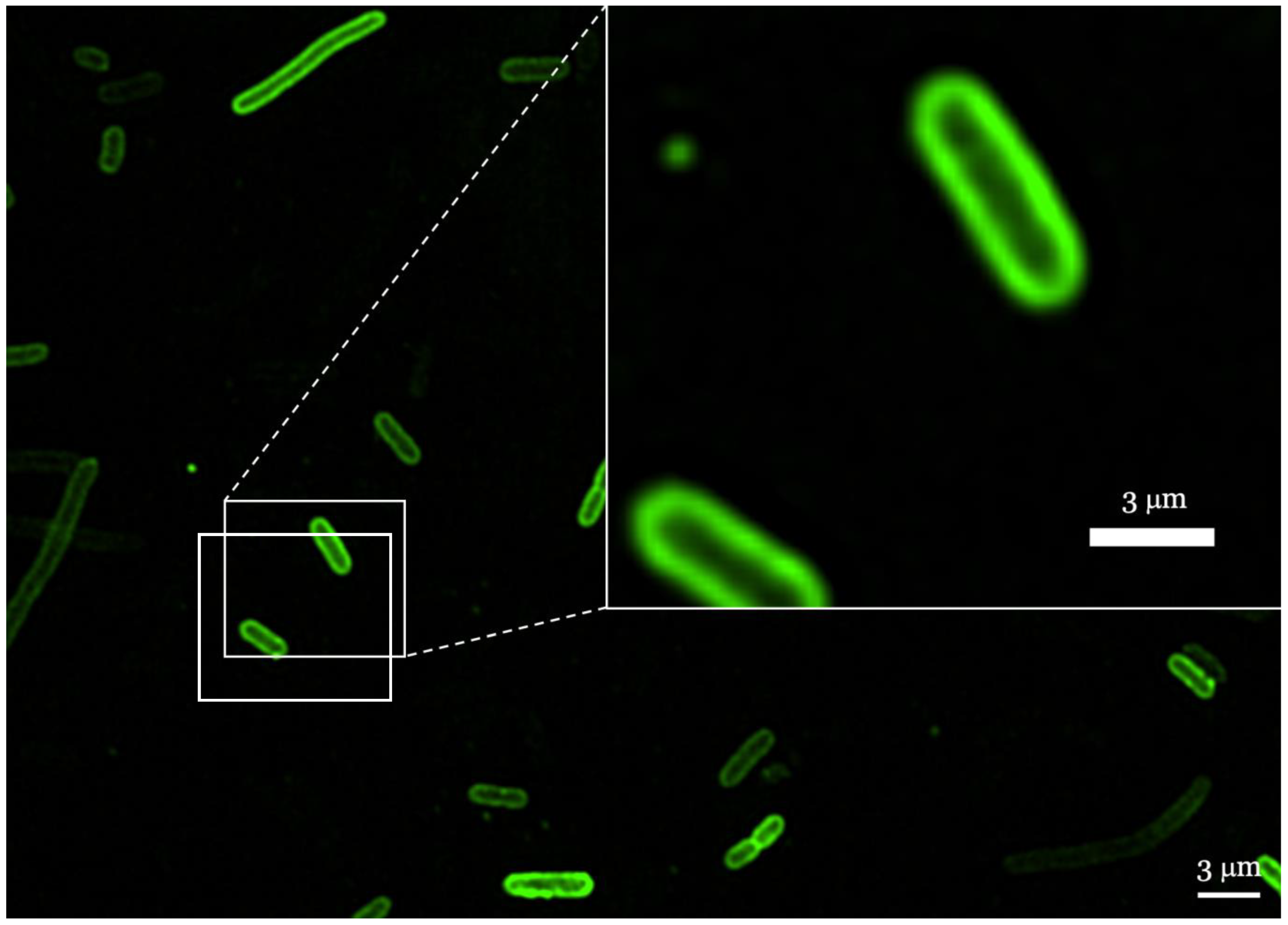
2.6. Localization of AMPs in Membranes
3. Discussion
4. Materials and Methods
4.1. Synthesis of Antimicrobial Peptides
4.2. Quantification of Antibacterial Activity
4.3. Confocal Microscopy
4.4. Cell Lines and Cultivation
4.5. Cytotoxicity Assay
4.6. Hemolysis Assay
4.7. Immunomodulatory Activity
4.7.1. Isolation of PBMNCs and Incubation with Peptides
4.7.2. Viability of PBMNCs
4.7.3. Quantification of TNF-α and IL-10
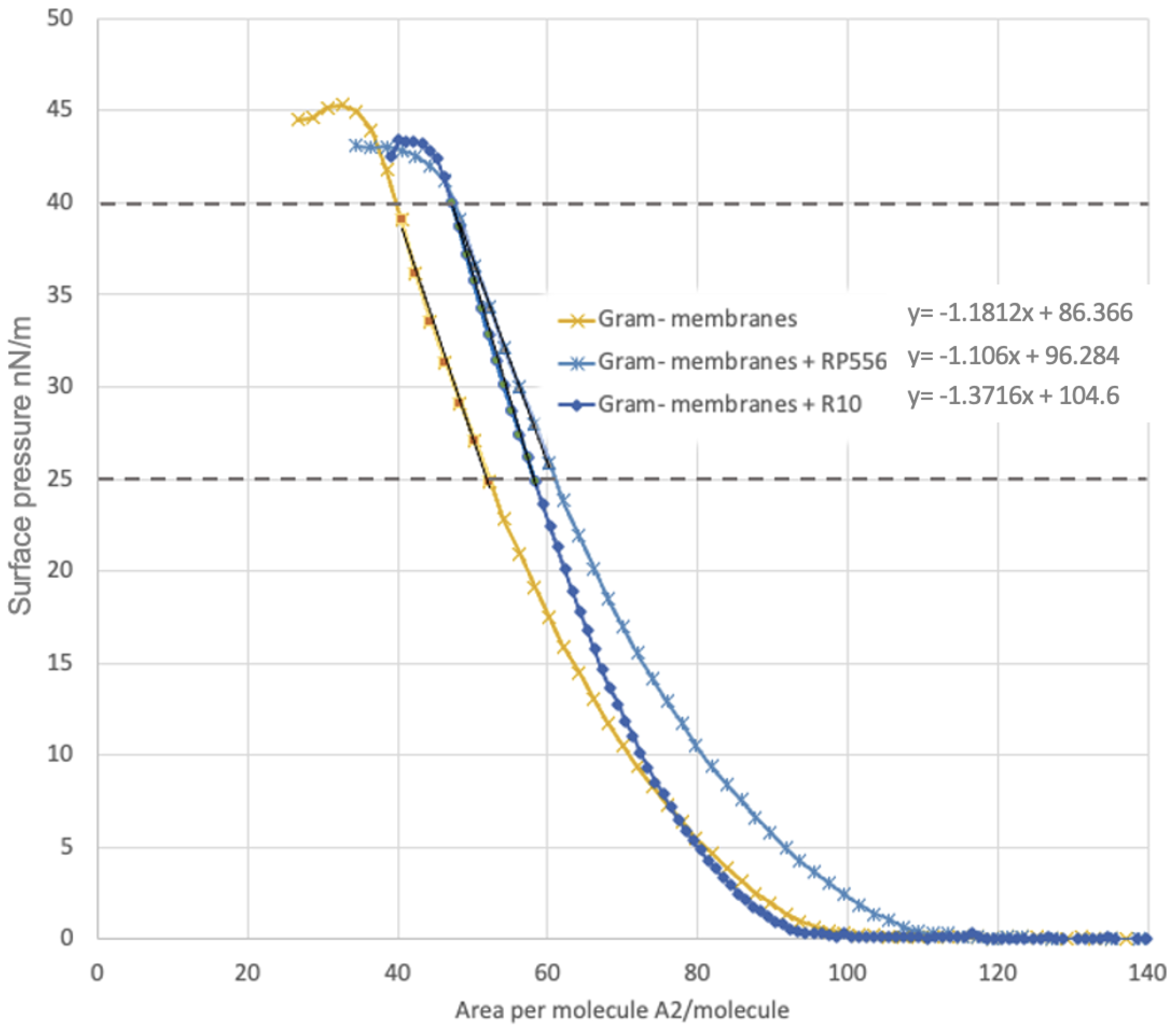
4.8. Langmuir−Blodgett Compression Isotherms
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mistry, R.D. Skin and Soft Tissue Infections. Pediatr. Clin. N. Am. 2013, 60, 1063–1082. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Noviello, S.; Leone, S. Epidemiology and Microbiology of Skin and Soft Tissue Infections. Curr. Opin. Infect. Dis. 2016, 29, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Ki, V.; Rotstein, C. Bacterial Skin and Soft Tissue Infections in Adults: A Review of Their Epidemiology, Pathogenesis, Diagnosis, Treatment and Site Of Care. Can. J. Infect. Dis. Med. Microbiol. 2008, 19, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.S.; Park, H.; Cho, S.; Yoon, H.-S. Antibiotic Susceptibility and Treatment Response in Bacterial Skin Infection. Ann. Dermatol. 2018, 30, 186. [Google Scholar] [CrossRef] [Green Version]
- Bassetti, M.; Baguneid, M.; Bouza, E.; Dryden, M.; Nathwani, D.; Wilcox, M. European Perspective and Update on the Management of Complicated Skin and Soft Tissue Infections Due to Methicillin-Resistant Staphylococcus Aureus after More than 10 Years of Experience with Linezolid. Clin. Microbiol. Infect. 2014, 20, 3–18. [Google Scholar] [CrossRef] [Green Version]
- De Macedo, G.H.R.V.; Costa, G.D.E.; Oliveira, E.R.; Damasceno, G.V.; Mendonça, J.S.P.; Silva, L.D.S.; Chagas, V.L.; Bazán, J.M.N.; Aliança, A.S.D.S.; de Cássia Mendonça de Miranda, R.; et al. Interplay between Eskape Pathogens and Immunity in Skin Infections: An Overview of the Major Determinants of Virulence and Antibiotic Resistance. Pathogens 2021, 10, 148. [Google Scholar] [CrossRef]
- Pletzer, D.; Mansour, S.C.; Hancock, R.E.W. Synergy between Conventional Antibiotics and Anti-Biofilm Peptides in a Murine, Sub-Cutaneous Abscess Model Caused by Recalcitrant ESKAPE Pathogens. PLoS Pathog. 2018, 14, e1007084. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef]
- Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report 2022; World Health Organization: Geneva, Switzerland, 2022; ISBN 9789240062702.
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia Coli. Microbiol. Spectr. 2018, 6, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Baum, H.; Marre, R. Antimicrobial Resistance of Escherichia Coli and Therapeutic Implications. Int. J. Med. Microbiol. 2005, 295, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; DeLeo, F.R. Waves of Resistance: Staphylococcus Aureus in the Antibiotic Era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The Characteristic of Virulence, Biofilm and Antibiotic Resistance of Klebsiella Pneumoniae. Int. J. Environ. Res. Public Health 2020, 17, 6278. [Google Scholar] [CrossRef] [PubMed]
- Haenni, M.; Lupo, A.; Madec, J.-Y. Antimicrobial Resistance in Streptococcus Spp. Microbiol. Spectr. 2018, 6, 973. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter Baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef]
- Puca, V.; Marulli, R.Z.; Grande, R.; Vitale, I.; Niro, A.; Molinaro, G.; Prezioso, S.; Muraro, R.; di Giovanni, P. Microbial Species Isolated from Infected Wounds and Antimicrobial Resistance Analysis: Data Emerging from a Three-Years Retrospective Study. Antibiotics 2021, 10, 1162. [Google Scholar] [CrossRef]
- Bharadwaj, A.; Rastogi, A.; Pandey, S.; Gupta, S.; Sohal, J.S. Multidrug-Resistant Bacteria: Their Mechanism of Action and Prophylaxis. Biomed. Res. Int. 2022, 2022, 5419874. [Google Scholar] [CrossRef]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic Resistance in Pseudomonas Aeruginosa—Mechanisms, Epidemiology and Evolution. Drug Resist. Updates 2019, 44, 100640. [Google Scholar] [CrossRef]
- Tan, J.K.L.; Bhate, K. A Global Perspective on the Epidemiology of Acne. Br. J. Dermatol. 2015, 172, 3–12. [Google Scholar] [CrossRef]
- Heng, A.H.S.; Chew, F.T. Systematic Review of the Epidemiology of Acne Vulgaris. Sci. Rep. 2020, 10, 5754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagnelie, M.; Poinas, A.; Dréno, B. What Is New in Adult Acne for the Last 2 Years: Focus on Acne Pathophysiology and Treatments. Int. J. Derm. 2022, 61, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Platsidaki, E.; Dessinioti, C. Recent Advances in Understanding Propionibacterium Acnes (Cutibacterium Acnes) in Acne. F1000Research 2018, 7, 1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuchayi, S.M.; Makrantonaki, E.; Ganceviciene, R.; Dessinioti, C.; Feldman, S.R.; Zouboulis, C.C. Acne Vulgaris. Nat. Rev. Dis. Prim. 2015, 1, 15029. [Google Scholar] [CrossRef] [PubMed]
- Dessinioti, C.; Katsambas, A. Antibiotics and Antimicrobial Resistance in Acne: Epidemiological Trends and Clinical Practice Considerations. Yale J. Biol. Med. 2022, 95, 429–443. [Google Scholar] [PubMed]
- Dessinioti, C.; Katsambas, A. Propionibacterium Acnes and Antimicrobial Resistance in Acne. Clin. Derm. 2017, 35, 163–167. [Google Scholar] [CrossRef]
- Chauhan, P.N.; Sharma, A.; Rasheed, H.; Mathur, H.; Sharma, P. Treatment Opportunities and Technological Progress Prospective for Acne Vulgaris. Curr. Drug Deliv. 2022, 19, 1037–1048. [Google Scholar] [CrossRef]
- Bassetti, M.; del Puente, F.; Magnasco, L.; Giacobbe, D.R. Innovative Therapies for Acute Bacterial Skin and Skin-Structure Infections (ABSSSI) Caused by Methicillin-Resistant Staphylococcus Aureus: Advances in Phase I and II Trials. Expert Opin. Investig. Drugs 2020, 29, 495–506. [Google Scholar] [CrossRef]
- Giacobbe, D.R.; Dettori, S.; Corcione, S.; Vena, A.; Sepulcri, C.; Maraolo, A.E.; de Rosa, F.G.; Bassetti, M. Emerging Treatment Options for Acute Bacterial Skin and Skin Structure Infections and Bloodstream Infections Caused by Staphylococcus Aureus: A Comprehensive Review of the Evidence. Infect. Drug Resist. 2022, 15, 2137–2157. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Sánchez-Pérez, A.; Calo-Mata, P.; Villa, T.G. Antimicrobial Peptides (AMPs): Ancient Compounds That Represent Novel Weapons in the Fight against Bacteria. Biochem. Pharm. 2017, 133, 117–138. [Google Scholar] [CrossRef]
- Gómez, D.M.; Lone, Y.-C.; Salazar, L.M.; Trojan, J. Antimicrobial Peptides, Novel Solution for the Treatment of Precancerous Disease Acne—A Review. Trends Med. 2019, 19, 1–6. [Google Scholar] [CrossRef]
- Benfield, A.H.; Henriques, S.T. Mode-of-Action of Antimicrobial Peptides: Membrane Disruption vs. Intracellular Mechanisms. Front. Med. Technol. 2020, 2, 610997. [Google Scholar] [CrossRef] [PubMed]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The Value of Antimicrobial Peptides in the Age of Resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef] [PubMed]
- Nuri, R.; Shprung, T.; Shai, Y. Defensive Remodeling: How Bacterial Surface Properties and Biofilm Formation Promote Resistance to Antimicrobial Peptides. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 3089–3100. [Google Scholar] [CrossRef] [Green Version]
- Felgueiras, H.P. An Insight into Biomolecules for the Treatment of Skin Infectious Diseases. Pharmaceutics 2021, 13, 1012. [Google Scholar] [CrossRef]
- Greco, I.; Molchanova, N.; Holmedal, E.; Jenssen, H.; Hummel, B.D.; Watts, J.L.; Håkansson, J.; Hansen, P.R.; Svenson, J. Correlation between Hemolytic Activity, Cytotoxicity and Systemic in Vivo Toxicity of Synthetic Antimicrobial Peptides. Sci. Rep. 2020, 10, 13206. [Google Scholar] [CrossRef]
- Lin, Q.; Deslouches, B.; Montelaro, R.C.; Di, Y.P. Prevention of ESKAPE Pathogen Biofilm Formation by Antimicrobial Peptides WLBU2 and LL37. Int. J. Antimicrob. Agents 2018, 52, 667–672. [Google Scholar] [CrossRef]
- Ridyard, K.E.; Overhage, J. The Potential of Human Peptide LL-37 as an Antimicrobial and Anti-Biofilm Agent. Antibiotics 2021, 10, 650. [Google Scholar] [CrossRef]
- Duplantier, A.J.; van Hoek, M.L. The Human Cathelicidin Antimicrobial Peptide LL-37 as a Potential Treatment for Polymicrobial Infected Wounds. Front. Immunol. 2013, 4, 143. [Google Scholar] [CrossRef] [Green Version]
- Batoni, G.; Maisetta, G.; Esin, S. Antimicrobial Peptides and Their Interaction with Biofilms of Medically Relevant Bacteria. Biochim. Biophys. Acta (BBA) Biomembr. 2016, 1858, 1044–1060. [Google Scholar] [CrossRef]
- Ting, D.S.J.; Beuerman, R.W.; Dua, H.S.; Lakshminarayanan, R.; Mohammed, I. Strategies in Translating the Therapeutic Potentials of Host Defense Peptides. Front. Immunol. 2020, 11, 983. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Lu, T.K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial Host Defence Peptides: Functions and Clinical Potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- McInturff, J.E.; Wang, S.-J.; Machleidt, T.; Richard Lin, T.; Oren, A.; Hertz, C.J.; Krutzik, S.R.; Hart, S.; Zeh, K.; Anderson, D.H.; et al. Granulysin-Derived Peptides Demonstrate Antimicrobial and Anti-Inflammatory Effects Against Propionibacterium Acnes. J. Investig. Dermatol. 2005, 125, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Harder, J.; Tsuruta, D.; Murakami, M.; Kurokawa, I. What Is the Role of Antimicrobial Peptides (AMP) in Acne Vulgaris? Exp. Derm. 2013, 22, 386–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodburn, K.W.; Jaynes, J.; Clemens, L.E. Designed Antimicrobial Peptides for Topical Treatment of Antibiotic Resistant Acne Vulgaris. Antibiotics 2020, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Mu, L.; Tang, J.; Duan, Z.; Wang, F.; Wei, L.; Rong, M.; Lai, R. A Small Peptide with Therapeutic Potential for Inflammatory Acne Vulgaris. PLoS ONE 2013, 8, e72923. [Google Scholar] [CrossRef] [Green Version]
- Han, R.; Blencke, H.-M.; Cheng, H.; Li, C. The Antimicrobial Effect of CEN1HC-Br against Propionibacterium Acnes and Its Therapeutic and Anti-Inflammatory Effects on Acne Vulgaris. Peptides 2018, 99, 36–43. [Google Scholar] [CrossRef]
- Elliott, A.G.; Huang, J.X.; Neve, S.; Zuegg, J.; Edwards, I.A.; Cain, A.K.; Boinett, C.J.; Barquist, L.; Lundberg, C.V.; Steen, J.; et al. An Amphipathic Peptide with Antibiotic Activity against Multidrug-Resistant Gram-Negative Bacteria. Nat. Commun. 2020, 11, 3184. [Google Scholar] [CrossRef]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W.; Somsri, S.; Aunpad, R. A Novel, Rationally Designed, Hybrid Antimicrobial Peptide, Inspired by Cathelicidin and Aurein, Exhibits Membrane-Active Mechanisms against Pseudomonas Aeruginosa. Sci. Rep. 2020, 10, 9117. [Google Scholar] [CrossRef]
- Timmons, P.B.; Hewage, C.M. APPTEST Is a Novel Protocol for the Automatic Prediction of Peptide Tertiary Structures. Brief. Bioinform. 2021, 22, 1–9. [Google Scholar] [CrossRef]
- Weber, M.; Steinle, H.; Golombek, S.; Hann, L.; Schlensak, C.; Wendel, H.P.; Avci-Adali, M. Blood-Contacting Biomaterials: In Vitro Evaluation of the Hemocompatibility. Front. Bioeng. Biotechnol. 2018, 6, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojewska, M.; Smułek, W.; Kaczorek, E.; Prochaska, K. Langmuir Monolayer Techniques for the Investigation of Model Bacterial Membranes and Antibiotic Biodegradation Mechanisms. Membranes 2021, 11, 707. [Google Scholar] [CrossRef] [PubMed]
- Seelig, A. Local Anesthetics and Pressure: A Comparison of Dibucaine Binding to Lipid Monolayers and Bilayers. Biochim. Biophys. Acta (BBA) Biomembr. 1987, 899, 196–204. [Google Scholar] [CrossRef]
- Fathi, F.; Ghobeh, M.; Tabarzad, M. Anti-Microbial Peptides: Strategies of Design and Development and Their Promising Wound-Healing Activities. Mol. Biol. Rep. 2022, 49, 9001–9012. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, S.; Rahman, N.A.A.; Peile, E.; Rahman, M.; Sartelli, M.; Hassali, M.A.; Islam, T.; Islam, S.; Haque, M. Microbial Resistance Movements: An Overview of Global Public Health Threats Posed by Antimicrobial Resistance, and How Best to Counter. Front. Public Health 2020, 8, 535668. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.; Straus, S. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Mishra, B.; Lushnikova, T.; Narayana, J.L.; Wang, G. Amino Acid Composition Determines Peptide Activity Spectrum and Hot-Spot-Based Design of Merecidin. Adv. Biosyst. 2018, 2, 1700259. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, Y.; Wang, Q.; Hang, B.; Sun, Y.; Wei, X.; Hu, J. Potential of Novel Antimicrobial Peptide P3 from Bovine Erythrocytes and Its Analogs to Disrupt Bacterial Membranes In Vitro and Display Activity against Drug-Resistant Bacteria in a Mouse Model. Antimicrob. Agents Chemother. 2015, 59, 2835–2841. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.K.; Lee, H.H.; Seo, C.H.; Park, Y. Antimicrobial and Immunomodulatory Properties and Applications of Marine-Derived Proteins and Peptides. Mar. Drugs 2019, 17, 350. [Google Scholar] [CrossRef] [Green Version]
- Tobiasz, A.; Nowicka, D.; Szepietowski, J.C. Acne Vulgaris—Novel Treatment Options and Factors Affecting Therapy Adherence: A Narrative Review. J. Clin. Med. 2022, 11, 7535. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Balaji, S.; Le, L.D.; Crombleholme, T.M.; Keswani, S.G. Regenerative Wound Healing: The Role of Interleukin-10. Adv. Wound Care 2014, 3, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leech, J.M.; Lacey, K.A.; Mulcahy, M.E.; Medina, E.; McLoughlin, R.M. IL-10 Plays Opposing Roles during Staphylococcus Aureus Systemic and Localized Infections. J. Immunol. 2017, 198, 2352–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travkova, O.G.; Moehwald, H.; Brezesinski, G. The Interaction of Antimicrobial Peptides with Membranes. Adv. Colloid Interface Sci. 2017, 247, 521–532. [Google Scholar] [CrossRef]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Rutkis, R.; Ļaša, Z.; Rubina, M.; Ščerbaka, R.; Kalniņš, G.; Bogans, J.; Kalnenieks, U. Antimicrobial Activity of Zymomonas Mobilis Is Related to Its Aerobic Catabolism and Acid Resistance. Fermentation 2022, 8, 77. [Google Scholar] [CrossRef]
- Hoyo, J.; Tzanov, J.T.-B.T. Lipid-Lipid Interactions of Escherichia Coli Mimetic Inner Membrane at Human Physiological Temperature. Gen. Physiol. Biophys. 2020, 39, 195–202. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Size kDa | Secondary Structure | Net Charge at pH7 | Reference |
|---|---|---|---|---|---|
| RP556 | RWCFKVCYKGICYKKCK | 2.16 | antipar.—β | +5.8 | [47] |
| LZ1 | VKRWKKWWRKWKKWV | 2.22 | α-helix | +8.0 | [48] |
| AA139 | GFCWYVCARRNGARVCYRRCN | 2.56 | antipar.—β | +4.8 | [50] |
| PA13 | KIAKRIWKRIWKILRRR | 2.32 | α-helix | +9.0 | [51] |
| Oligo10 | FAKALKALLKALKAL | 1.59 | α-helix | +4.0 | Commercial |
| R1 | FIKKFAKKFKKFIKKFAKFAFAF | 2.86 | α-helix | +9.0 | This work |
| R10 | KKIAKKFWKKFWKFWKIFKK | 2.74 | α-helix | +10.0 | This work |
| R11 | KFCLKFCFKGFCFKACGK | 2.11 | antipar.—β | +9.7 | This work |
| R12 | IAKKFWKKFWKFWKIFKKIA | 2.67 | α-helix | 8.0 | This work |
| R13 | IAKKFWPKFWKFWKIFKKIA | 2.67 | α-helix | 7.0 | This work |
| R14 | KKIAKKFWKKFWKFWPKFWKIFKK | 3.31 | α-helix | 11.0 | This work |
| MIC μg/mL | |||||||
|---|---|---|---|---|---|---|---|
| Name | Reference | E. coli | P. aeruginosa | K. pneumonia | E. faecium | S. aureus | C. acnes |
| RP556 | [53] | 4.0 | 2.0 | 4.0 | 2.0 | 1.0 | 2.0 |
| LZ1 | [48] | 31.0 | 8.0 | 16.0 | 4.0 | 2.0 | 8.0 |
| AA139 | [50] | 4.0 | 16.0 | 8.0 | 16.0 | 8.0 | 63.0 |
| PA13 | [51] | 31.0 | 16.0 | 16.0 | 4.0 | 2.0 | 16.0 |
| Oligo10 | Commercial product | 16.0 | 63.0 | 63.0 | 16.0 | 4.0 | 2.0 |
| R1 | This work | 8.0 | 8.0 | 4.0 | 16.0 | 8.0 | 2.0 |
| R10 | This work | 2.0 | 2.0 | 8.0 | 4.0 | 4.0 | 0.5 |
| R11 | This work | 125.0 | 63.0 | 63.0 | 16.0 | 16.0 | 0.3 |
| R12 | This work | 8.0 | 4.0 | 31.0 | 16.0 | 8.0 | 0.5 |
| R13 | This work | 4.0 | 4.0 | 8.0 | 16.0 | 4.0 | 0.3 |
| R14 | This work | 16.0 | 4.0 | 4.0 | 8.0 | 8.0 | 1.0 |
| Therapeutic Index | |||||||
|---|---|---|---|---|---|---|---|
| Reference | E. coli | P. aeruginosa | K. pneumonia | E. faecium | S. aureus | C. acnes | |
| RP556 | [47] | 96.5 | 193.0 | 96.5 | 193.0 | 386.0 | 193.0 |
| LZ1 | [48] | 5.9 | 22.9 | 11.4 | 45.8 | 91.5 | 22.9 |
| AA139 | [50] | 157.0 | 39.3 | 78.5 | 39.3 | 78.5 | 10.0 |
| PA13 | [51] | 8.5 | 16.4 | 16.4 | 65.5 | 131.0 | 16.4 |
| Oligo10 | Commercial product | 13.6 | 3.5 | 3.5 | 13.6 | 54.5 | 109.0 |
| R1 | This work | 8.6 | 8.6 | 17.3 | 4.3 | 8.6 | 34.5 |
| R11 | this work | 0.9 | 1.7 | 1.7 | 6.7 | 6.7 | 356.7 |
| R12 | this work | 2.0 | 4.0 | 0.5 | 1.0 | 2.0 | 32.0 |
| R13 | this work | 19.8 | 19.8 | 9.9 | 4.9 | 19.8 | 263.3 |
| R14 | this work | 2.2 | 8.8 | 8.8 | 4.4 | 4.4 | 35.0 |
| Therapeutic Index | |||||||
|---|---|---|---|---|---|---|---|
| Reference | E. coli | P. aeruginosa | K. pneumonia | E. faecium | S. aureus | C. acnes | |
| R1 | This work | 16.01 | 16.01 | 32.02 | 8.0 | 16.01 | 64.04 |
| RP556 | [47] | 86.43 | 172.85 | 86.43 | 172.85 | 345.7 | 172.85 |
| Oligo10 | Commercial product | 15.1 | 3.83 | 3.83 | 15.1 | 60.39 | 120.77 |
| R10 | this work | 19.1 | 19.1 | 4.77 | 9.55 | 9.55 | 76.38 |
| Sample | Aπ→0 | π Collapse mN/m |
|---|---|---|
| Gram-negative membranes | 73.1 | 45.3 |
| Gram-negative membranes + RP556 | 83.4 | 43.6 |
| Gram-negative membranes + R10 | 76.3 | 43.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramata-Stunda, A.; Boroduskis, M.; Kaktina, E.; Patetko, L.; Kalnenieks, U.; Lasa, Z.; Rubina, M.; Strazdina, I.; Kalnins, G.; Rutkis, R. Comparative Evaluation of Existing and Rationally Designed Novel Antimicrobial Peptides for Treatment of Skin and Soft Tissue Infections. Antibiotics 2023, 12, 551. https://doi.org/10.3390/antibiotics12030551
Ramata-Stunda A, Boroduskis M, Kaktina E, Patetko L, Kalnenieks U, Lasa Z, Rubina M, Strazdina I, Kalnins G, Rutkis R. Comparative Evaluation of Existing and Rationally Designed Novel Antimicrobial Peptides for Treatment of Skin and Soft Tissue Infections. Antibiotics. 2023; 12(3):551. https://doi.org/10.3390/antibiotics12030551
Chicago/Turabian StyleRamata-Stunda, Anna, Martins Boroduskis, Elza Kaktina, Liene Patetko, Uldis Kalnenieks, Zane Lasa, Marta Rubina, Inese Strazdina, Gints Kalnins, and Reinis Rutkis. 2023. "Comparative Evaluation of Existing and Rationally Designed Novel Antimicrobial Peptides for Treatment of Skin and Soft Tissue Infections" Antibiotics 12, no. 3: 551. https://doi.org/10.3390/antibiotics12030551