
Plant Secondary Metabolites on Efflux-Mediated Antibiotic Resistant Stenotrophomonas Maltophilia: Potential of Herbal-Derived Efflux Pump Inhibitors
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
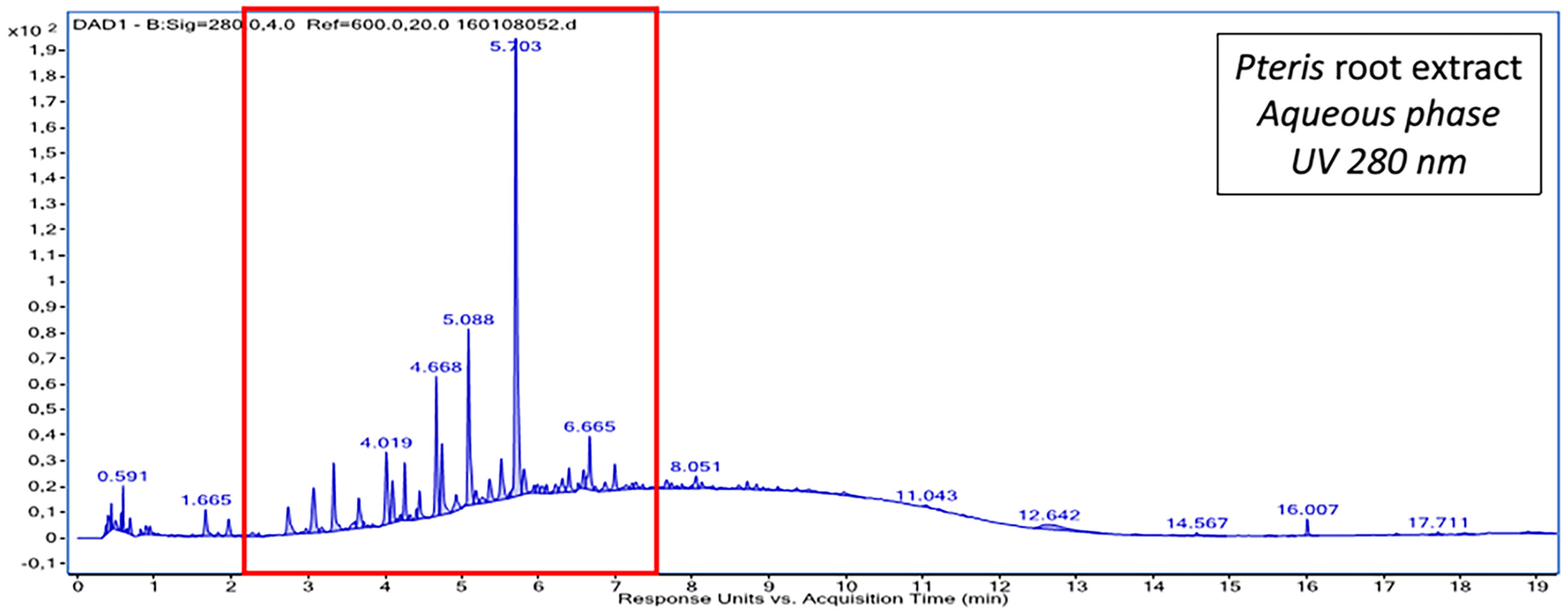
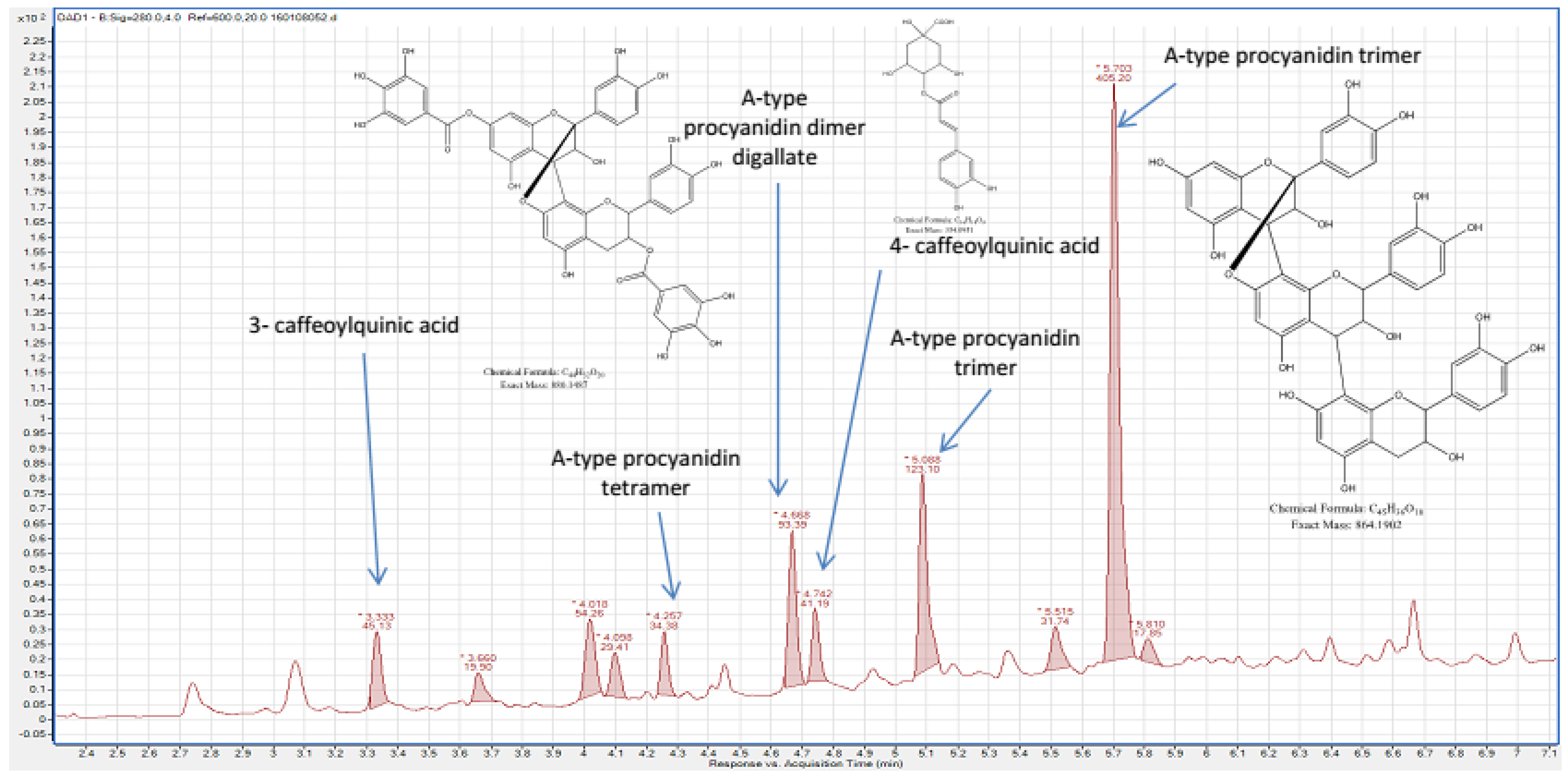
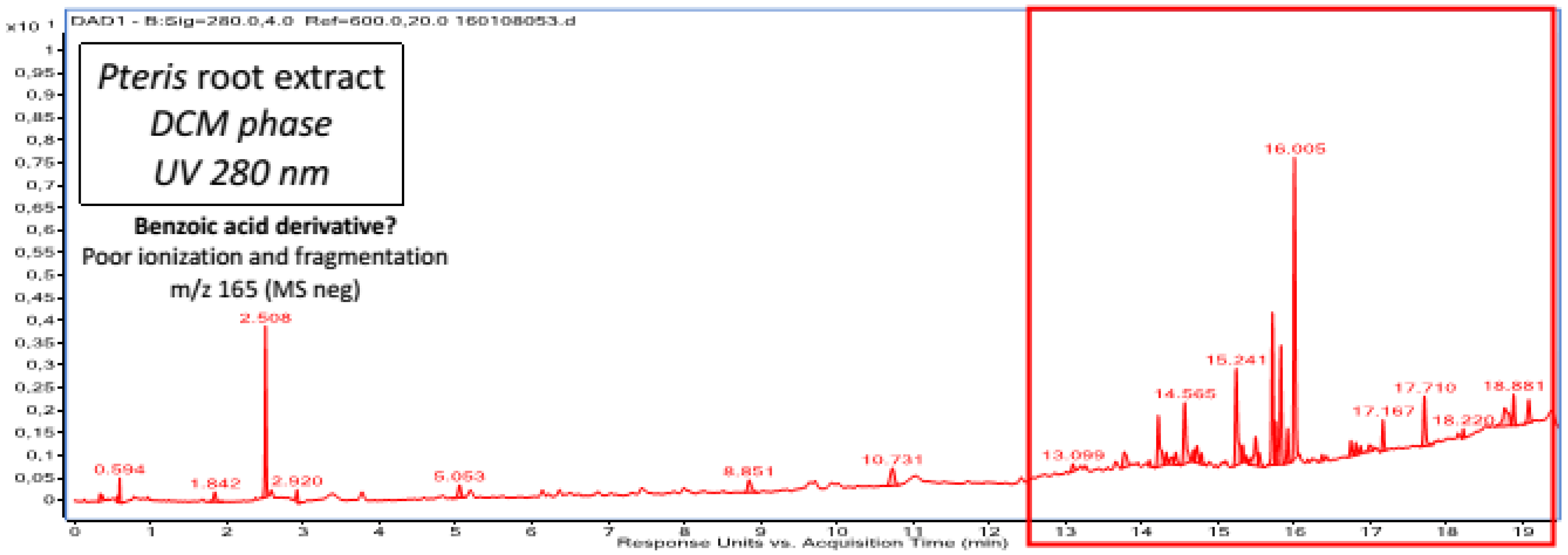
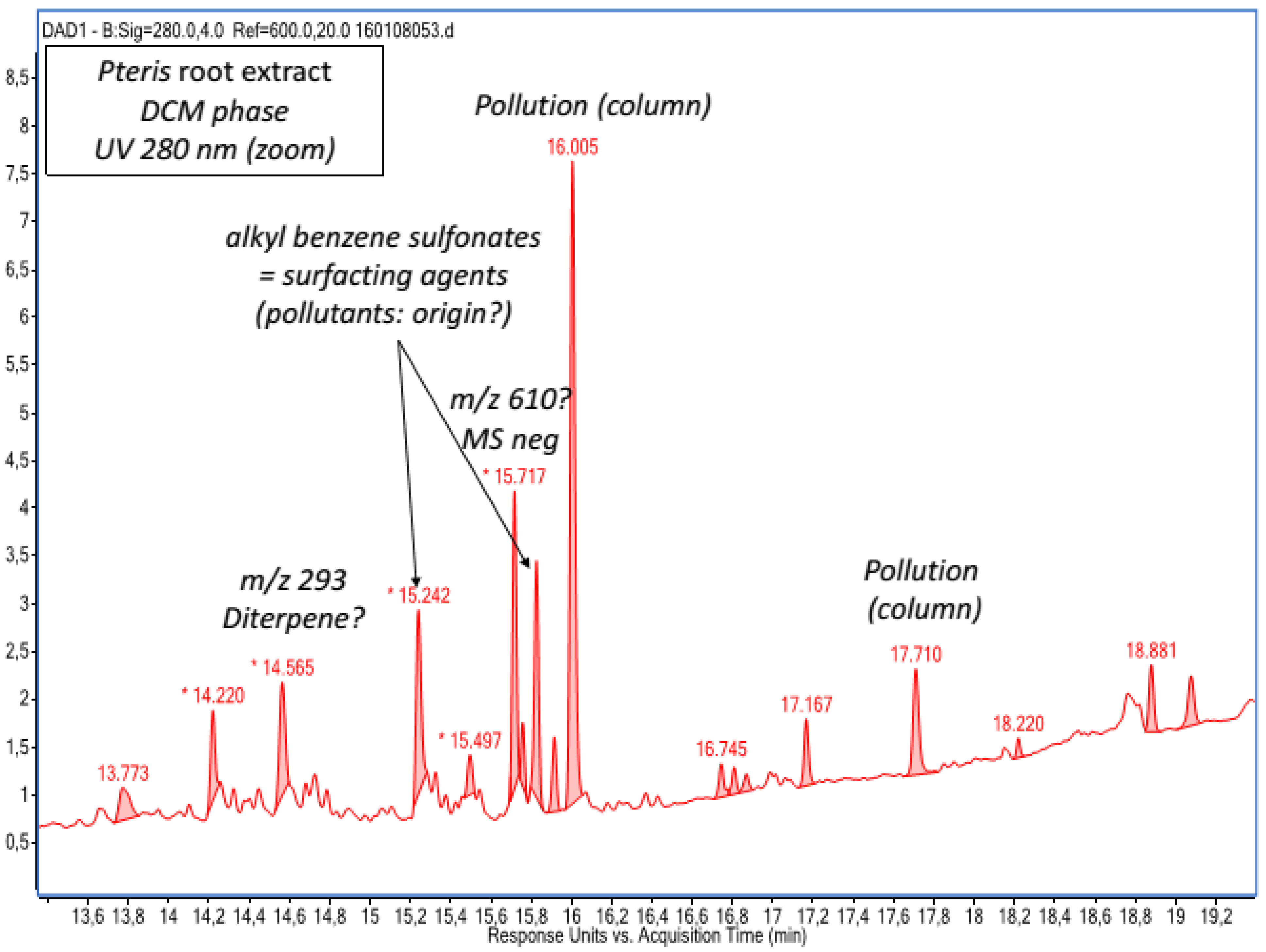
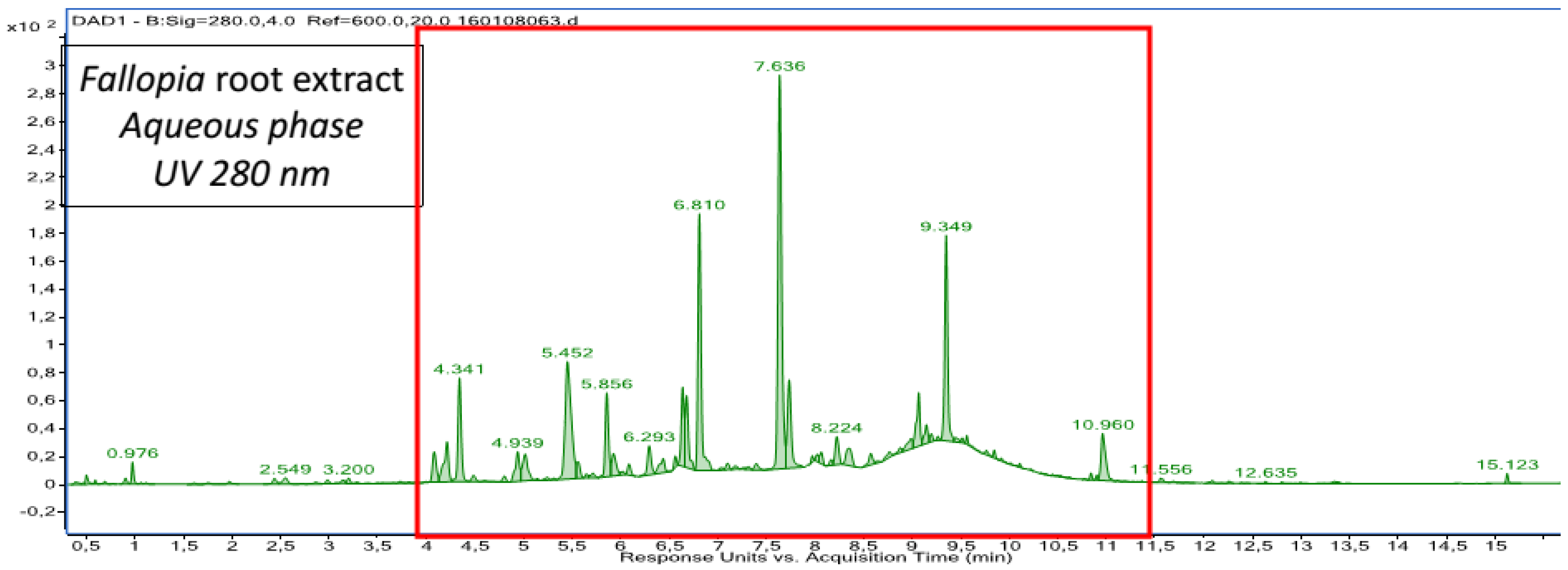
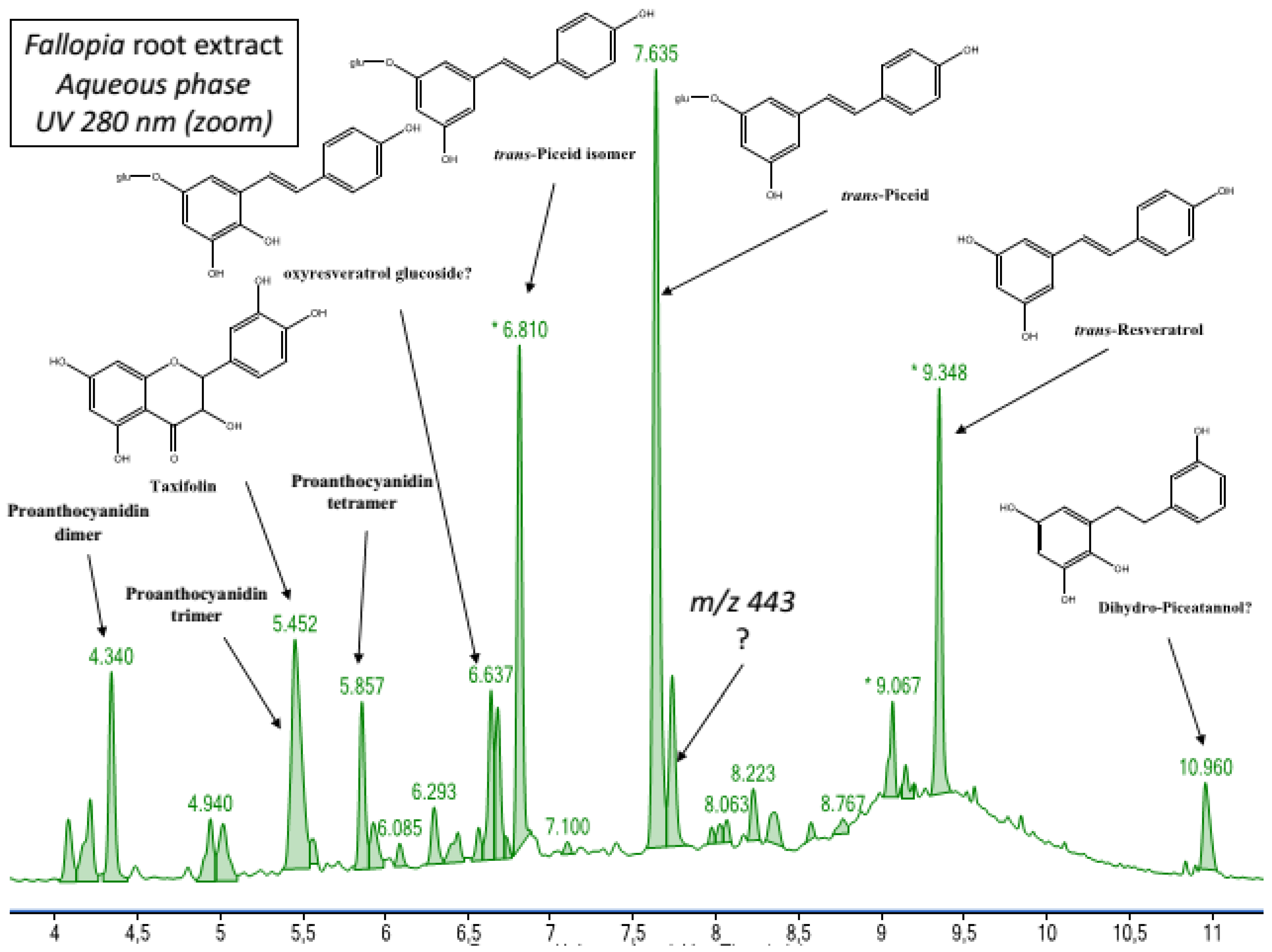
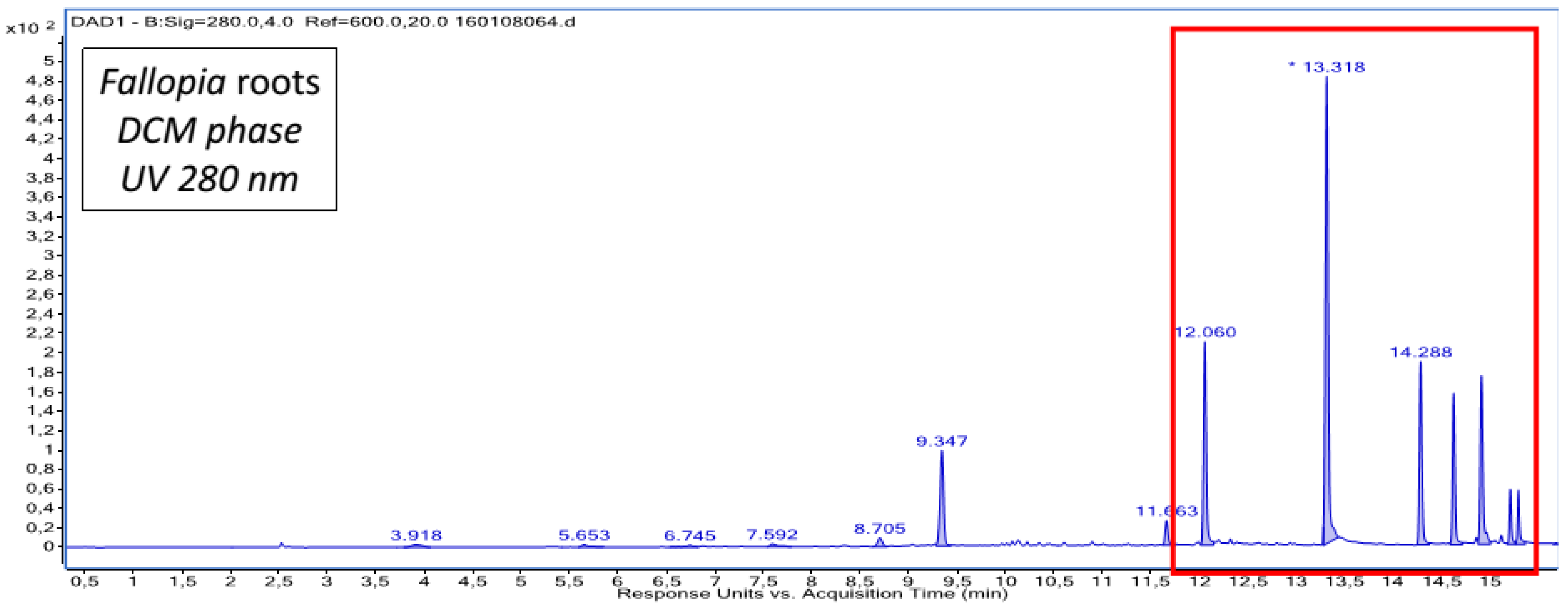
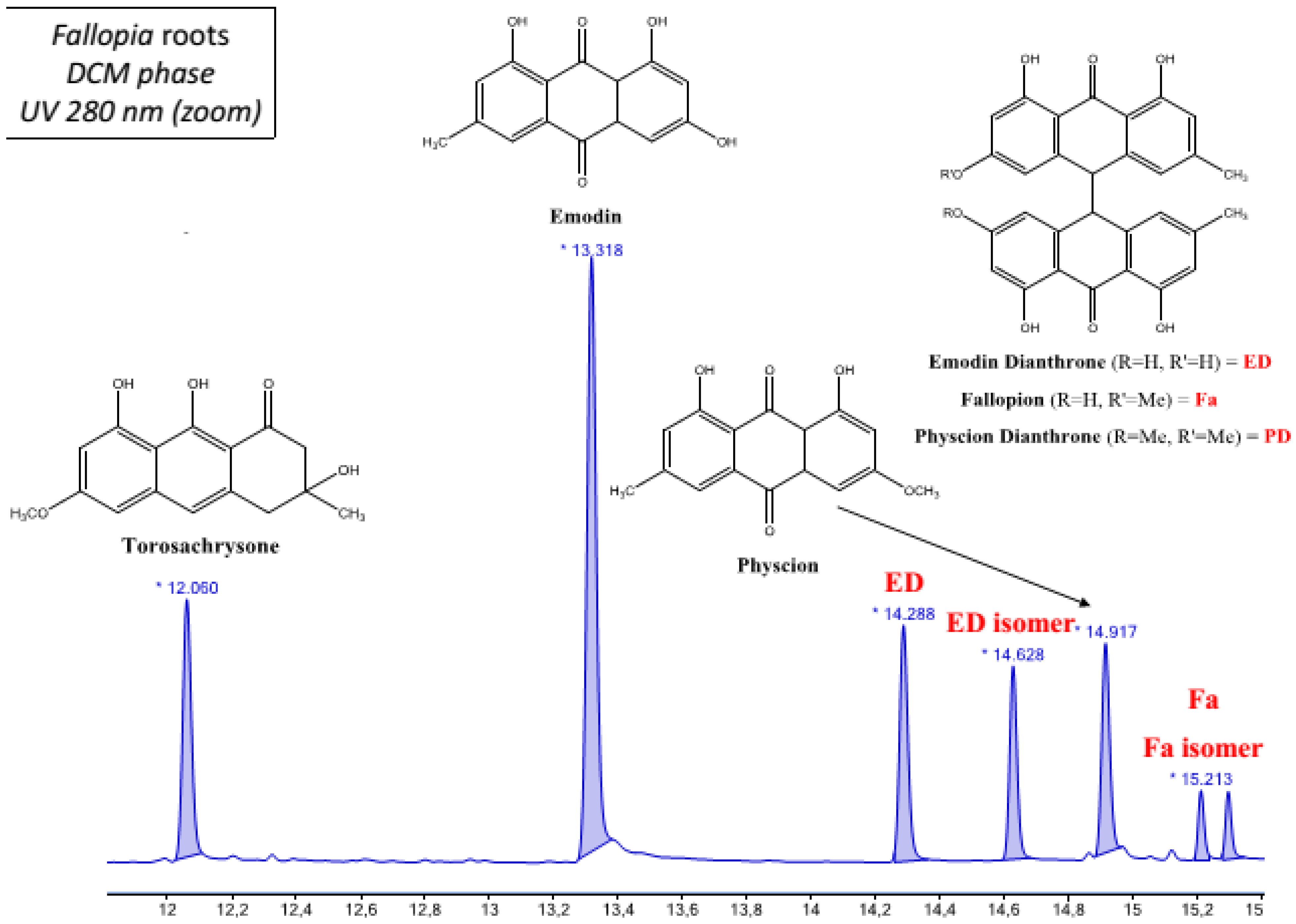
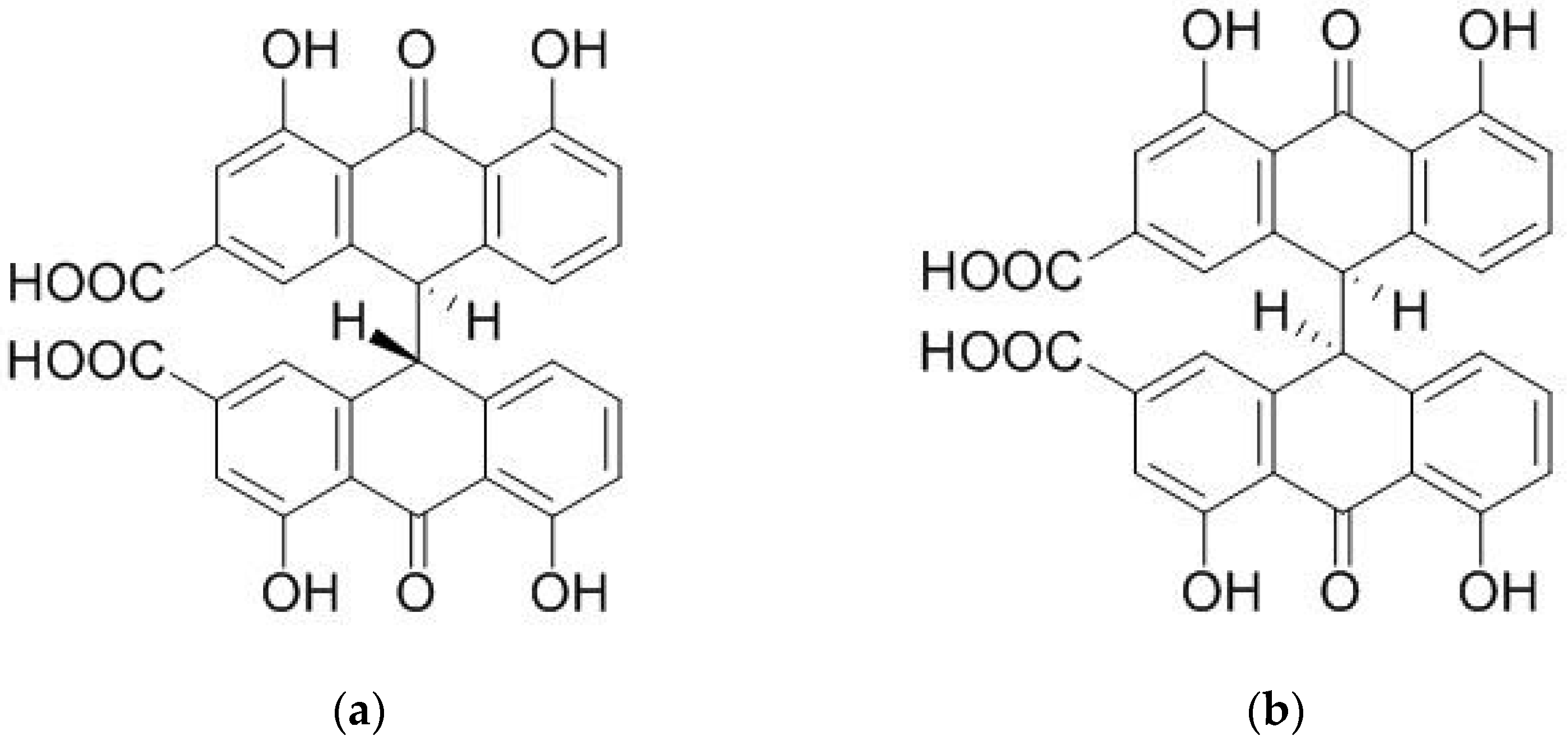
2.1. Chemical Composition of Fractions
2.2. Minimum Inhibitory Concentration Determination
2.3. Efflux Pump Inhibitory Activity Evaluation
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A









References
- Gil-Gil, T.; Martínez, J.L.; Blanco, P. Mechanisms of antimicrobial resistance in Stenotrophomonas maltophilia: A review of current knowledge. Expert Rev. Anti Infect. Ther. 2020, 18, 335–347. [Google Scholar] [CrossRef]
- Youenou, B.; Favre-Bonté, S.; Bodilis, J.; Brothier, E.; Dubost, A.; Muller, D.; Nazaret, S. Comparative Genomics of Environmental and Clinical Stenotrophomonas maltophilia Strains with Different Antibiotic Resistance Profiles. Genome Biol. Evol. 2015, 7, 2484–2505. [Google Scholar] [CrossRef] [Green Version]
- Henderson, P.J.F.; Maher, C.; Elbourne, L.D.H.; Eijkelkamp, B.A.; Paulsen, I.T.; Hassan, K.A. Physiological Functions of Bacterial “Multidrug” Efflux Pumps. Chem. Rev. 2021, 121, 5417–5478. [Google Scholar] [CrossRef]
- Tegos, G.P.; Haynes, M.; Strouse, J.J.; Khan, M.M.T.; Bologa, C.G.; Oprea, T.I.; Sklar, L.A. Microbial efflux pump inhibition: Tactics and strategies. Curr. Pharm. Des. 2011, 17, 1291–1302. [Google Scholar] [CrossRef] [Green Version]
- Michalet, S.; Rouifed, S.; Pellassa-Simon, T.; Fusade-Boyer, M.; Meiffren, G.; Nazaret, S.; Piola, F. Tolerance of Japanese knotweed s.l. to soil artificial polymetallic pollution: Early metabolic responses and performance during vegetative multiplication. Environ. Sci. Pollut. Res. 2017, 24, 20897. [Google Scholar] [CrossRef]
- Pham, H.N.; Michalet, S.; Bodillis, J.; Nguyen, T.D.; Nguyen, T.K.O.; Le, T.P.Q.; Haddad, M.; Nazaret, S.; Dijoux-Franca, M.-G. Impact of metal stress on the production of secondary metabolites in Pteris vittata L. and associated rhizosphere bacterial communities. Environ. Sci. Pollut. Res. Int. 2017, 24, 16735–16750. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, L.; Zheng, Y.; Xue, S.; Zhang, J.; Huang, P.; Zhao, Y.; Hao, X.; He, Z.; Hu, Z.; et al. Insight into the assembly of root-associated microbiome in the medicinal plant Polygonum cuspidatum. Ind. Crops Prod. 2020, 145, 112163. [Google Scholar] [CrossRef]
- Ma, L.Q.; Komar, K.M.; Tu, C.; Zhang, W.; Cai, Y.; Kennelley, E.D. A fern that hyperaccumulates arsenic. Nature 2001, 409, 579. [Google Scholar] [CrossRef]
- An, Z.-Z.; Huang, Z.-C.; Lei, M.; Liao, X.-Y.; Zheng, Y.-M.; Chen, T.-B. Zinc tolerance and accumulation in Pteris vittata L. and its potential for phytoremediation of Zn- and As-contaminated soil. Chemosphere 2006, 62, 796–802. [Google Scholar] [CrossRef]
- Danh, L.T.; Truong, P.; Mammucari, R.; Foster, N. A critical review of the arsenic uptake mechanisms and phytoremediation potential of Pteris vittata. Int. J. Phytoremediation 2014, 16, 429–453. [Google Scholar] [CrossRef]
- Anh, B.T.K.; Kim, D.D.; Tua, T.V.; Kien, N.T.; Anh, D.T. Phytoremediation potential of indigenous plants from Thai Nguyen province, Vietnam. J. Environ. Biol. Acad. Environ. Biol. India 2011, 32, 257–262. [Google Scholar]
- Bui, T.K.A.; Dang, D.K.; Nguyen, T.K.; Nguyen, N.M.; Nguyen, Q.T.; Nguyen, H.C. Phytoremediation of heavy metal polluted soil and water in Vietnam. J. Vietnam. Environ. 2014, 6, 47–51. [Google Scholar] [CrossRef]
- Nguyen, N.-L.; Bui, V.-H.; Pham, H.-N.; To, H.-M.; Dijoux-Franca, M.-G.; Vu, C.-T.; Nguyen, K.-O.T. Ionomics and metabolomics analysis reveal the molecular mechanism of metal tolerance of Pteris vittata L. dominating in a mining site in Thai Nguyen province, Vietnam. Environ. Sci. Pollut. Res. 2022, 29, 87268–87280. [Google Scholar] [CrossRef] [PubMed]
- Nguyen Thi, K.-O.; Nguyen, N.-L.; Pham, H.-N.; Sawada, Y.; Hirai, M.Y.; Dauwe, R.; Dijoux-Franca, M.-G. Development of a Pteris vittata L. compound database by widely targeted metabolomics profiling. Biomed. Chromatogr. 2021, 35, e5110. [Google Scholar] [CrossRef]
- Paul, T.; Das, B.; Apte, K. Suchitra Hypoglycemic Activity of Pteris vittata L., a Fern on Alloxan Induced Diabetic Rats. Inven. Rapid Planta Act. 2012, 2, 88–91. [Google Scholar]
- Gong, X.-L.; Chen, Z.-H.; Liang, N.-C. Advances in study on chemical constituents and pharmacological activities of plants of genus Pteris. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China Chin. Mater. Medica 2007, 32, 1382–1387. [Google Scholar]
- Dupuy, J.; Larrieu, G.; Sutra, J.F.; Lespine, A.; Alvinerie, M. Enhancement of moxidectin bioavailability in lamb by a natural flavonoid: Quercetin. Vet. Parasitol. 2003, 112, 337–347. [Google Scholar] [CrossRef]
- Choi, J.-S.; Jo, B.-W.; Kim, Y.-C. Enhanced paclitaxel bioavailability after oral administration of paclitaxel or prodrug to rats pretreated with quercetin. Eur. J. Pharm. Biopharm. Off. J. Arb. Pharm. Verfahr. EV 2004, 57, 313–318. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Chao, P.-D.L.; Hsiu, S.L.; Wen, K.-C.; Hou, Y.-C. Lethal quercetin-digoxin interaction in pigs. Life Sci. 2004, 74, 1191–1197. [Google Scholar] [CrossRef]
- Chiruvella, K.K.; Mohammed, A.; Dampuri, G.; Ghanta, R.G.; Raghavan, S.C. Phytochemical and Antimicrobial Studies of Methyl Angolensate and Luteolin-7-O-glucoside Isolated from Callus Cultures of Soymida febrifuga. Int. J. Biomed. Sci. IJBS 2007, 3, 269–278. [Google Scholar]
- Le, H.-L.; To, D.-C.; Tran, M.-H.; Do, T.-T.; Nguyen, P.-H. Natural PTP1B Inhibitors From Polygonum cuspidatum and Their 2-NBDG Uptake Stimulation. Nat. Prod. Commun. 2020, 15, 1934578X20961201. [Google Scholar] [CrossRef]
- Zhang, H.; Li, C.; Kwok, S.-T.; Zhang, Q.-W.; Chan, S.-W. A Review of the Pharmacological Effects of the Dried Root of Polygonum cuspidatum (Hu Zhang) and Its Constituents. Evid. Based Complement. Alternat. Med. 2013, 2013, e208349. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-C.; Chen, Y.-T.; Chiu, C.-C.; Liao, W.-T.; Liu, Y.-C.; David Wang, H.-M. Polygonum cuspidatum extracts as bioactive antioxidaion, anti-tyrosinase, immune stimulation and anticancer agents. J. Biosci. Bioeng. 2015, 119, 464–469. [Google Scholar] [CrossRef]
- Cucu, A.-A.; Baci, G.-M.; Dezsi, Ş.; Nap, M.-E.; Beteg, F.I.; Bonta, V.; Bobiş, O.; Caprio, E.; Dezmirean, D.S. New Approaches on Japanese Knotweed (Fallopia japonica) Bioactive Compounds and Their Potential of Pharmacological and Beekeeping Activities: Challenges and Future Directions. Plants 2021, 10, 2621. [Google Scholar] [CrossRef]
- Li, L.; Song, X.; Yin, Z.; Jia, R.; Li, Z.; Zhou, X.; Zou, Y.; Li, L.; Yin, L.; Yue, G.; et al. The antibacterial activity and action mechanism of emodin from Polygonum cuspidatum against Haemophilus parasuis in vitro. Microbiol. Res. 2016, 186–187, 139–145. [Google Scholar] [CrossRef]
- de Oliveira-Garcia, D.; Dall’Agnol, M.; Rosales, M.; Azzuz, A.C.G.S.; Alcántara, N.; Martinez, M.B.; Girón, J.A. Fimbriae and adherence of Stenotrophomonas maltophilia to epithelial cells and to abiotic surfaces. Cell. Microbiol. 2003, 5, 625–636. [Google Scholar] [CrossRef]
- Alonso, A.; Sanchez, P.; Martínez, J.L. Stenotrophomonas maltophilia D457R contains a cluster of genes from gram-positive bacteria involved in antibiotic and heavy metal resistance. Antimicrob. Agents Chemother. 2000, 44, 1778–1782. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, X.Z.; Poole, K. Fluoroquinolone susceptibilities of efflux-mediated multidrug-resistant Pseudomonas aeruginosa, Stenotrophomonas maltophilia and Burkholderia cepacia. J. Antimicrob. Chemother. 2001, 48, 549–552. [Google Scholar] [CrossRef]
- Li, X.; Hu, J.; Wang, B.; Sheng, L.; Liu, Z.; Yang, S.; Li, Y. Inhibitory effects of herbal constituents on P-glycoprotein in vitro and in vivo: Herb-drug interactions mediated via P-gp. Toxicol. Appl. Pharmacol. 2014, 275, 163–175. [Google Scholar] [CrossRef]
- Li, X.; Dong, Y.; Wang, W.; Wang, H.; Chen, Y.; Shi, G.; Yi, J.; Wang, J. Emodin as an effective agent in targeting cancer stem-like side population cells of gallbladder carcinoma. Stem Cells Dev. 2013, 22, 554–566. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, A.K.; Mohanty, P. Bacterial efflux pumps involved in multidrug resistance and their inhibitors: Rejuvinating the antimicrobial chemotherapy. Recent Patents Anti-Infect. Drug Disc. 2012, 7, 73–89. [Google Scholar] [CrossRef]
- Rao, M.; Padyana, S.; Dipin, K.M.; Kumar, S.; Nayak, B.B.; Varela, M.F. Antimicrobial Compounds of Plant Origin as Efflux Pump Inhibitors: New Avenues for Controlling Multidrug Resistant Pathogens. J. Antimicrob. Agents 2018, 4, 1–6. [Google Scholar] [CrossRef]
- Huang, Y.-Q.; Huang, G.-R.; Wu, M.-H.; Tang, H.-Y.; Huang, Z.-S.; Zhou, X.-H.; Yu, W.-Q.; Su, J.-W.; Mo, X.-Q.; Chen, B.-P.; et al. Inhibitory effects of emodin, baicalin, schizandrin and berberine on hefA gene: Treatment of Helicobacter pylori-induced multidrug resistance. World J. Gastroenterol. WJG 2015, 21, 4225–4231. [Google Scholar] [CrossRef]
- Lechner, D.; Gibbons, S.; Bucar, F. Plant phenolic compounds as ethidium bromide efflux inhibitors in Mycobacterium smegmatis. J. Antimicrob. Chemother. 2008, 62, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Rahman, F.I.; Hussain, F.; Rahman, S.M.A.; Rahman, M.M. Antimicrobial Diterpenes: Recent Development From Natural Sources. Front. Pharmacol. 2022, 12, 4141. [Google Scholar] [CrossRef]
- Singh, H. Potential medicinal pteridophytes of India and their chemical constituents. J. Econ. Tax. Bot. 1999, 23, 63–78. [Google Scholar]
- Nath, K.; Bhattacharya, M.K.; Kar, S. Antimicrobial Potential of Ethnomedicinal Ferns of Southern Assam, India. Indian J. Pharm. Sci. 2018, 80, 556–560. [Google Scholar] [CrossRef] [Green Version]
- Gibb, J.; Wong, D.W. Antimicrobial Treatment Strategies for Stenotrophomonas maltophilia: A Focus on Novel Therapies. Antibiotics 2021, 10, 1226. [Google Scholar] [CrossRef]
- Zając, O.M.; Tyski, S.; Laudy, A.E. Phenotypic and Molecular Characteristics of the MDR Efflux Pump Gene-Carrying Stenotrophomonas maltophilia Strains Isolated in Warsaw, Poland. Biology 2022, 11, 105. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.-Z.; Poole, K. SmeDEF Multidrug Efflux Pump Contributes to Intrinsic Multidrug Resistance in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2001, 45, 3497–3503. [Google Scholar] [CrossRef] [Green Version]
- Gould, V.C.; Okazaki, A.; Avison, M.B. Coordinate Hyperproduction of SmeZ and SmeJK Efflux Pumps Extends Drug Resistance in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2013, 57, 655–657. [Google Scholar] [CrossRef] [Green Version]
- Impey, R.E.; Hawkins, D.A.; Sutton, J.M.; Soares da Costa, T.P. Overcoming Intrinsic and Acquired Resistance Mechanisms Associated with the Cell Wall of Gram-Negative Bacteria. Antibiot. Basel Switz. 2020, 9, 623. [Google Scholar] [CrossRef]
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-W.; Liou, R.-S.; Lin, Y.-T.; Huang, H.-H.; Yang, T.-C. A Linkage between SmeIJK Efflux Pump, Cell Envelope Integrity, and σE-Mediated Envelope Stress Response in Stenotrophomonas maltophilia. PLoS ONE 2014, 9, e111784. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-T.; Huang, Y.-W.; Chen, S.-J.; Chang, C.-W.; Yang, T.-C. The SmeYZ Efflux Pump of Stenotrophomonas maltophilia Contributes to Drug Resistance, Virulence-Related Characteristics, and Virulence in Mice. Antimicrob. Agents Chemother. 2015, 59, 4067–4073. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-W.; Huang, Y.-W.; Hu, R.-M.; Yang, T.-C. SmeOP-TolCSm Efflux Pump Contributes to the Multidrug Resistance of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2014, 58, 2405–2408. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-T.; Huang, Y.-W.; Liou, R.-S.; Chang, Y.-C.; Yang, T.-C. MacABCsm, an ABC-type tripartite efflux pump of Stenotrophomonas maltophilia involved in drug resistance, oxidative and envelope stress tolerances and biofilm formation. J. Antimicrob. Chemother. 2014, 69, 3221–3226. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. vittata L. | F. japonica | |
|---|---|---|
| Aqueous phase | Procyanidin trimer A-type a Procyanidin dimer digallate A-type a Procyanidin tetramer A-type a 4- caffeoylquinic acid 3- caffeoylquinicacid p-coumaroyl pentoside acid | trans-Piceid and its isomer a trans-Resveratrol a Oxyresveratrolglucoside a Proanthocyanidin tetramer B-type Proanthocyanidin trimer B-type Proanthocyanidin dimer B-type Taxifolin |
| DCM phase | Unidentified diterpene a | Emodin a Emodin dianthrone and its isomer a Torosachrysone a Physcion a Fallopion and its isomer a Resveratrol derivative |
| Samples | Clinical Strains | Environmental Strains | |||||
|---|---|---|---|---|---|---|---|
| Plant | Solvent | 502 | 0366 | K279a | PierC1 | BurA1 | BurE1 |
| P. vittata L. | H2O | NA * | >2000 | >2000 | >2000 | >2000 | >2000 |
| DCM | >2000 | >2000 | >2000 | >2000 | >2000 | >2000 | |
| F. japonica | H2O | >2000 | >2000 | >2000 | >2000 | >2000 | >2000 |
| DCM | >1000 | 31.25 | >1000 | NA | >1000 | NA | |
| S. maltophillia Strains | 502 | 0366 | K279a | PierC1 | BurA1 | BurE1 |
|---|---|---|---|---|---|---|
| GEN | 20 | 5 | 80 | 10 | 40 | 320 |
| GEN+PAβN | >80 | 20 | 40 | 10 | 20 | 80 |
| Concentration (μg/mL) | P. vittata L. | F. japonica | ||
|---|---|---|---|---|
| H2O | DCM | H2O | DCM | |
| 250 | (X) | 2-fold for BurE1 | (X) | 4-fold for BurA1 8-fold for BurE1 |
| 125 | 4-fold for K279a, BurA1 4-fold for BurE1 | |||
| 62.5 | 2-fold for K279a, BurE1 | |||
| K279a | BurA1 | BurE1 | |
|---|---|---|---|
| Emodin | 6.28 ± 0.20 | −2.05 ± 1.47 | 1.37 ± 2.06 |
| Physcion | −1.75 ± 0.88 | −1.22 ± 0.70 | −1.34 ± 1.75 |
| Sennidine A | −8.56 ± 0.95 | 2.07 ± 0.75 | 1.50 ± 0.28 |
| Sennidine B | −7.88 ± 2.05 | 4.02 ± 1.41 | 0.81 ± 0.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.H.T.; Nguyen, N.A.T.; Nguyen, H.D.; Nguyen, T.T.H.; Le, M.H.; Pham, M.Q.; Do, H.N.; Hoang, K.C.; Michalet, S.; Dijoux-Franca, M.-G.; et al. Plant Secondary Metabolites on Efflux-Mediated Antibiotic Resistant Stenotrophomonas Maltophilia: Potential of Herbal-Derived Efflux Pump Inhibitors. Antibiotics 2023, 12, 421. https://doi.org/10.3390/antibiotics12020421
Nguyen THT, Nguyen NAT, Nguyen HD, Nguyen TTH, Le MH, Pham MQ, Do HN, Hoang KC, Michalet S, Dijoux-Franca M-G, et al. Plant Secondary Metabolites on Efflux-Mediated Antibiotic Resistant Stenotrophomonas Maltophilia: Potential of Herbal-Derived Efflux Pump Inhibitors. Antibiotics. 2023; 12(2):421. https://doi.org/10.3390/antibiotics12020421
Chicago/Turabian StyleNguyen, Thi Huyen Thu, Ngoc Anh Thơ Nguyen, Hai Dang Nguyen, Thi Thu Hien Nguyen, Mai Huong Le, Minh Quan Pham, Huu Nghi Do, Kim Chi Hoang, Serge Michalet, Marie-Geneviève Dijoux-Franca, and et al. 2023. "Plant Secondary Metabolites on Efflux-Mediated Antibiotic Resistant Stenotrophomonas Maltophilia: Potential of Herbal-Derived Efflux Pump Inhibitors" Antibiotics 12, no. 2: 421. https://doi.org/10.3390/antibiotics12020421