
Mitochondria-Targeted Curcumin: A Potent Antibacterial Agent against Methicillin-Resistant Staphylococcus aureus with a Possible Intracellular ROS Accumulation as the Mechanism of Action

 ,
,
Abstract
:1. Introduction
2. Results
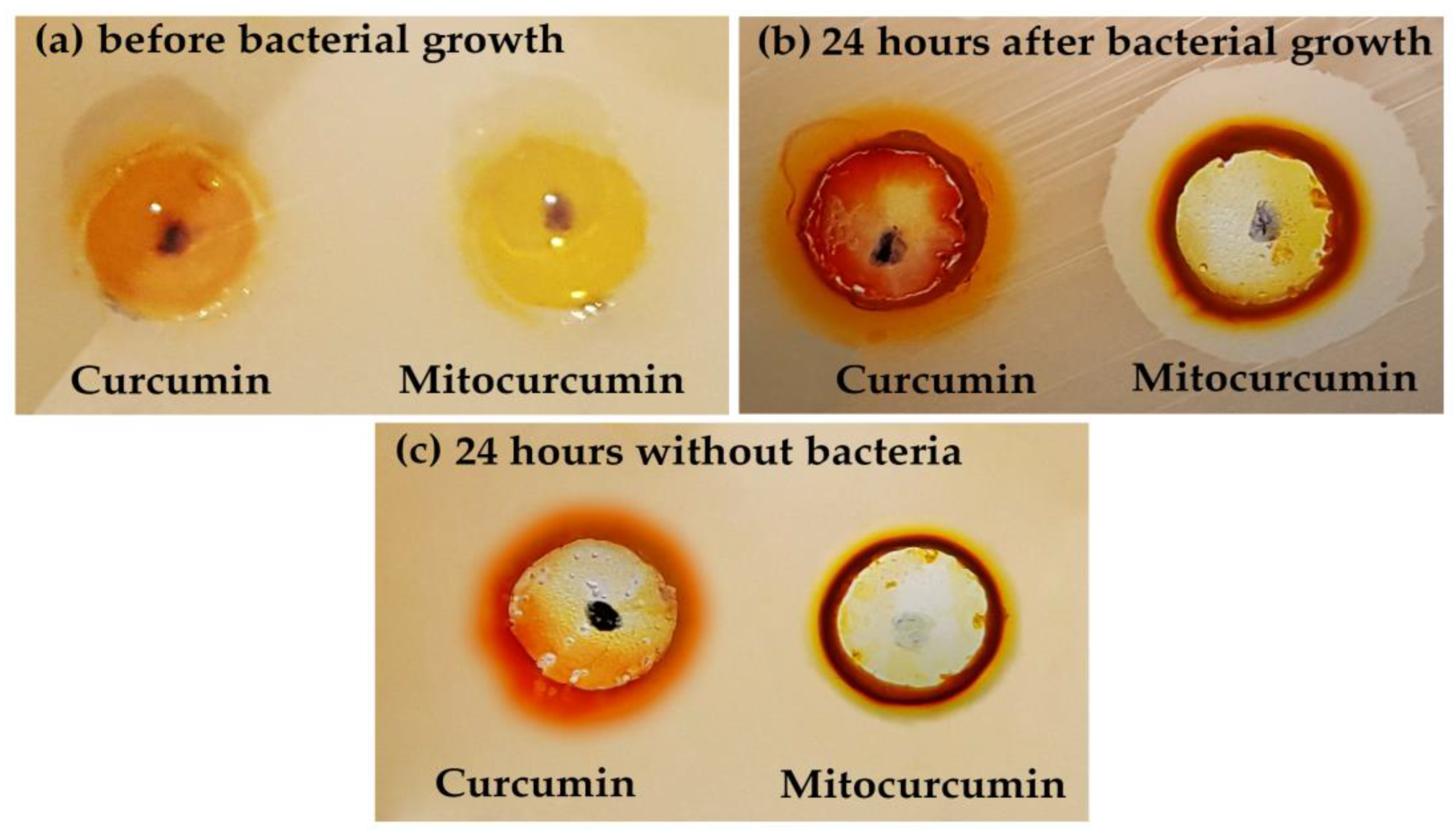
2.1. The Antibacterial Effect of Mitocurcumin Is Significantly Higher Than Curcumin
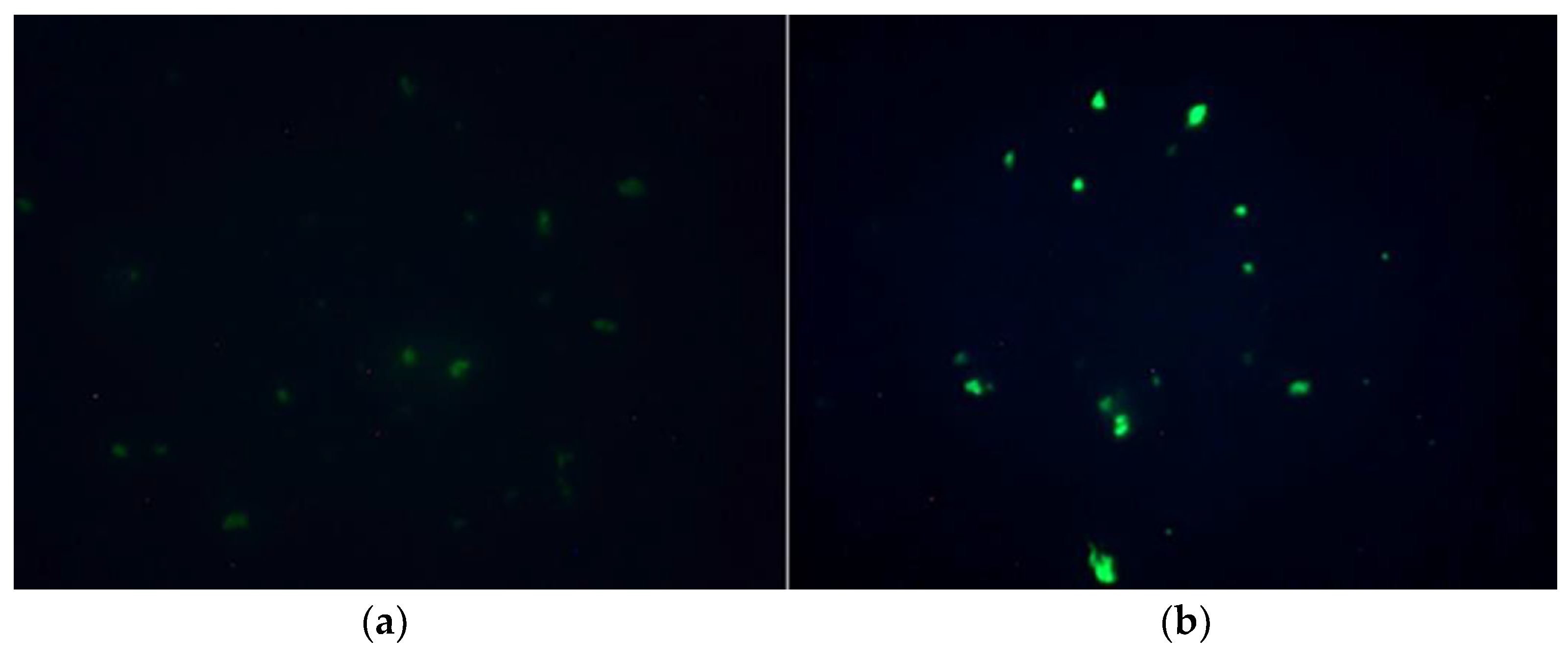
2.2. Both Compounds Generated Bacteria Fluorescence but Mitocurcumin Displayed a Stronger Effect
2.3. Fluorescence Microscopy Studies Showed Reactive Species Accumulation for a S. pombe Eukaryotic Model
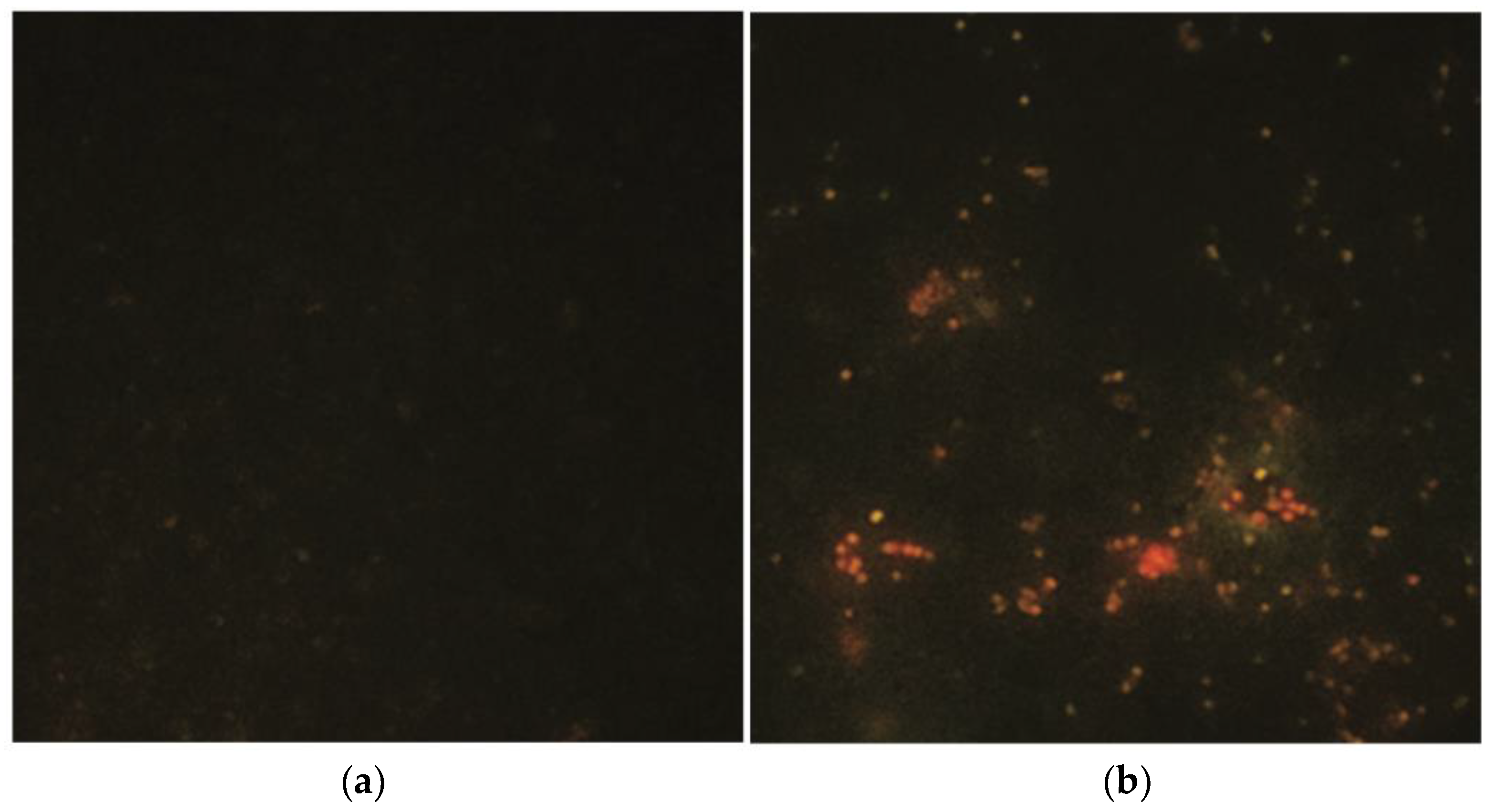
2.4. The Same Results Were Obtained with MitoTracker for the Prokaryotic Strains of MRSA (Figure 5)

2.5. The Antibacterial Activity of Mitocurcumin Is Modulated by Glutathione and Menadione
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Strains and Media for the S. pombe Model
4.3. Maintenance and Preservation of Microorganisms
4.4. Reagents
4.5. Preparation of Stock Solutions
4.6. Evaluation of Antimicrobial Activity
4.7. Fluorescence Microscopy Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khardori, N.; Stevaux, C.; Ripley, K. Antibiotics: From the Beginning to the Future: Part 1. Indian J. Pediatr. 2019, 87, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Khardori, N.; Stevaux, C.; Ripley, K. Antibiotics: From the Beginning to the Future: Part 2. Indian J. Pediatr. 2019, 87, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Arumugham, V.B.; Cascella, M. Third Generation Cephalosporins. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2019. [Google Scholar]
- Noureddin, S.A.; El-Shishtawy, R.M.; Al-Footy, K.O. Curcumin analogues and their hybrid molecules as multifunctional drugs. Eur. J. Med. Chem. 2019, 182, 111631. [Google Scholar] [CrossRef] [PubMed]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Campanini-Salinas, J.; Andrades-Lagos, J.; Mella-Raipan, J.; Vasquez-Velasquez, D. Novel Classes of Antibacterial Drugs in Clinical Development, a Hope in a Post-antibiotic Era. Curr. Top. Med. Chem. 2018, 18, 1188–1202. [Google Scholar] [CrossRef]
- Anand, P.; Thomas, S.G.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Sung, B.; Tharakan, S.T.; Misra, K.; Priyadarsini, I.K.; Rajasekharan, K.N.; et al. Biological activities of curcumin and its analogues (Congeners) made by man and Mother Nature. Biochem. Pharmacol. 2008, 76, 1590–1611. [Google Scholar] [CrossRef]
- Bagheri, H.; Ghasemi, F.; Barreto, G.E.; Rafiee, R.; Sathyapalan, T.; Sahebkar, A. Effects of curcumin on mitochondria in neurodegenerative diseases. BioFactors 2019, 46, 5–20. [Google Scholar] [CrossRef]
- Den Hartogh, D.J.; Gabriel, A.; Tsiani, E. Antidiabetic Properties of Curcumin II: Evidence from In Vivo Studies. Nutrients 2019, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Giordano, A.; Tommonaro, G. Curcumin and Cancer. Nutrients 2019, 11, 2367. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef] [Green Version]
- Moghadamtousi, S.Z.; Kadir, H.A.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. BioMed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef] [Green Version]
- Teow, S.Y.; Liew, K.; Ali, S.A.; Khoo, A.S.; Peh, S.C. Antibacterial Action of Curcumin against Staphylococcus aureus: A Brief Review. J. Trop. Med. 2016, 2016, 2853045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, A.; Ghodsi, R.; Kooshki, F.; Karimi, M.; Asghariazar, V.; Tarighat-Esfanjani, A. Therapeutic effects of curcumin on sepsis and mechanisms of action: A systematic review of preclinical studies. Phytother. Res. PTR 2019, 33, 2798–2820. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, P.; Singh, M.; Kumari, H.; Kumari, A.; Mukhopadhyay, K. Bactericidal activity of curcumin I is associated with damaging of bacterial membrane. PLoS ONE 2015, 10, e0121313. [Google Scholar] [CrossRef] [Green Version]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef] [Green Version]
- Burgos-Moron, E.; Calderon-Montano, J.M.; Salvador, J.; Robles, A.; Lopez-Lazaro, M. The dark side of curcumin. Int. J. Cancer 2010, 126, 1771–1775. [Google Scholar] [CrossRef]
- Mun, S.H.; Joung, D.K.; Kim, Y.S.; Kang, O.H.; Kim, S.B.; Seo, Y.S.; Kim, Y.C.; Lee, D.S.; Shin, D.W.; Kweon, K.T.; et al. Synergistic antibacterial effect of curcumin against methicillin-resistant Staphylococcus aureus. Phytomed. Int. J. Phytother. Phytopharm. 2013, 20, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.H.; Kim, S.B.; Kong, R.; Choi, J.G.; Kim, Y.C.; Shin, D.W.; Kang, O.H.; Kwon, D.Y. Curcumin reverse methicillin resistance in Staphylococcus aureus. Molecules 2014, 19, 18283–18295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielonka, J.; Joseph, J.; Sikora, A.; Hardy, M.; Ouari, O.; Vasquez-Vivar, J.; Cheng, G.; Lopez, M.; Kalyanaraman, B. Mitochondria-Targeted Triphenylphosphonium-Based Compounds: Syntheses, Mechanisms of Action, and Therapeutic and Diagnostic Applications. Chem. Rev. 2017, 117, 10043–10120. [Google Scholar] [CrossRef]
- Oyewole, A.O.; Birch-Machin, M.A. Mitochondria-targeted antioxidants. FASEB J 2015, 29, 4766–4771. [Google Scholar] [CrossRef] [Green Version]
- Sassi, N.; Mattarei, A.; Azzolini, M.; Bernardi, P.; Szabo; Ildiko; Paradisi, C.; Zoratti, M.; Biasutto, L. Mitochondria-targeted Resveratrol Derivatives Act as Cytotoxic Pro-oxidants. Curr. Pharm. Des. 2014, 20, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, S.; Jayakumar, S.; Gupta, G.D.; Bihani, S.C.; Sharma, D.; Kutala, V.K.; Sandur, S.K.; Kumar, V. Antibacterial activity of new structural class of semisynthetic molecule, triphenyl-phosphonium conjugated diarylheptanoid. Free Radic. Biol. Med. 2019, 143, 140–145. [Google Scholar] [CrossRef]
- Zinovkin, R.A.; Zamyatnin, A.A. Mitochondria-Targeted Drugs. Curr. Mol. Pharmacol. 2019, 12, 202–214. [Google Scholar] [CrossRef]
- Lopresti, A.L. The Problem of Curcumin and Its Bioavailability: Could Its Gastrointestinal Influence Contribute to Its Overall Health-Enhancing Effects? Adv. Nutr. 2018, 9, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Hay, E.; Lucariello, A.; Contieri, M.; Esposito, T.; De Luca, A.; Guerra, G.; Perna, A. Therapeutic effects of turmeric in several diseases: An overview. Chem. Biol. Interact. 2019, 310, 108729. [Google Scholar] [CrossRef]
- Kumari, S.; Jayakumar, S.; Bihani, S.C.; Shetake, N.; Naidu, R.; Kutala, V.K.; Sarma, H.D.; Gupta, G.D.; Sandur, S.K.; Kumar, V. Pharmacological characterization of a structurally new class of antibacterial compound, triphenyl-phosphonium conjugated diarylheptanoid: Antibacterial activity and molecular mechanism. J. Biosci. 2020, 45, 147. [Google Scholar] [CrossRef] [PubMed]
- Nazarov, P.A.; Osterman, I.A.; Tokarchuk, A.V.; Karakozova, M.V.; Korshunova, G.A.; Lyamzaev, K.G.; Skulachev, M.V.; Kotova, E.A.; Skulachev, V.P.; Antonenko, Y.N. Mitochondria-targeted antioxidants as highly effective antibiotics. Sci. Rep. 2017, 7, 1394. [Google Scholar] [CrossRef] [PubMed]
- Khailova, L.S.; Nazarov, P.A.; Sumbatyan, N.V.; Korshunova, G.A.; Rokitskaya, T.I.; Dedukhova, V.I.; Antonenko, Y.N.; Skulachev, V.P. Uncoupling and Toxic Action of Alkyltriphenylphosphonium Cations on Mitochondria and the Bacterium Bacillus subtilis as a Function of Alkyl Chain Length. Biochemistry 2015, 80, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Severin, F.F.; Severina, I.I.; Antonenko, Y.N.; Rokitskaya, T.I.; Cherepanov, D.A.; Mokhova, E.N.; Vyssokikh, M.Y.; Pustovidko, A.V.; Markova, O.V.; Yaguzhinsky, L.S.; et al. Penetrating cation/fatty acid anion pair as a mitochondria-targeted protonophore. Proc. Natl. Acad. Sci. USA 2010, 107, 663–668. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Koh, J.J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane Active Antimicrobial Peptides: Translating Mechanistic Insights to Design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Van Acker, H.; Coenye, T. The Role of Reactive Oxygen Species in Antibiotic-Mediated Killing of Bacteria. Trends Microbiol. 2017, 25, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, S.; Patwardhan, R.S.; Pal, D.; Singh, B.; Sharma, D.; Kutala, V.K.; Sandur, S.K. Mitochondrial targeted curcumin exhibits anticancer effects through disruption of mitochondrial redox and modulation of TrxR2 activity. Free Radic. Biol. Med. 2017, 113, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Silver, L.L. Multi-targeting by monotherapeutic antibacterials. Nat. Rev. Drug Discov. 2007, 6, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Besnard, J.; Ruda, G.F.; Setola, V.; Abecassis, K.; Rodriguiz, R.M.; Huang, X.P.; Norval, S.; Sassano, M.F.; Shin, A.I.; Webster, L.A.; et al. Automated design of ligands to polypharmacological profiles. Nature 2012, 492, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, N.; Zhang, Z.; Liao, F.; Jiang, T.; Tu, Y. The birth of artemisinin. Pharmacol. Ther. 2020, 216, 107658. [Google Scholar] [CrossRef]
- Jiang, Y.; Leung, A.W.; Hua, H.; Rao, X.; Xu, C. Photodynamic Action of LED-Activated Curcumin against Staphylococcus aureus Involving Intracellular ROS Increase and Membrane Damage. Int. J. Photoenergy 2014, 2014, 637601. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 18 January 2023).
- Bhalodi, A.A.; Oppermann, N.; Campeau, S.A.; Humphries, R.M. Variability of Beta-Lactam Broth Microdilution for Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2021, 65, e0064021. [Google Scholar] [CrossRef]
- Stoica, B.A.; Rusu, M.; Petreus, T.; Nechifor, M. Manganese SOD Mimics Are Effective Against Heat Stress in a Mutant Fission Yeast Deficient in Mitochondrial Superoxide Dismutase. Biol. Trace Elem. Res. 2011, 144, 1344–1350. [Google Scholar] [CrossRef]
- Bonev, B.; Hooper, J.; Parisot, J. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [Google Scholar] [CrossRef] [Green Version]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K.; CLSI Methods Development and Standardization Working Group of the Subcommittee on Antimicrobial Susceptibility Testing. CLSI Methods Development and Standardization Working Group Best Practices for Evaluation of Antimicrobial Susceptibility Tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Maslov, I.; Bogorodskiy, A.; Mishin, A.; Okhrimenko, I.; Gushchin, I.; Kalenov, S.; Dencher, N.A.; Fahlke, C.; Büldt, G.; Gordeliy, V.; et al. Efficient non-cytotoxic fluorescent staining of halophiles. Sci. Rep. 2018, 8, 2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osório, N.S.; Carvalho, A.; Almeida, A.J.; Padilla-Lopez, S.; Leão, C.; Laranjinha, J.; Ludovico, P.; Pearce, D.A.; Rodrigues, F. Nitric Oxide Signaling Is Disrupted in the Yeast Model for Batten Disease. Mol. Biol. Cell 2007, 18, 2755–2767. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Inhibition Diameter mm (Curcumin) | Inhibition Diameter mm (Mitocurcumin) |
|---|---|---|
| MRSA strain (ATCC 700698) | 0 | 17 ± 1 |
| Strain | MIC—µM (Curcumin) | MIC—µM (Mitocurcumin) |
|---|---|---|
| MRSA strain (ATCC 700698) | >250 | 1.95 |
| HS1 * | >250 | 1.95 |
| HS2 * | >250 | 1.95 |
| HS3 * | >250 | 1.95 |
| ATCC 29213 | >250 | 1.95 |
| Strain | MIC—µM (mitocurcumin) | MIC—µM (mitocurcumin + glutathione) | MIC—µM (mitocurcumin + menadione) |
|---|---|---|---|
| MRSA strain (ATCC 700698) | 1.95 | 3.9 | 0.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leferman, C.-E.; Stoica, L.; Stoica, B.A.; Ciubotaru, A.D.; Badescu, A.C.; Bogdanici, C.-M.; Neagu, T.P.; Ghiciuc, C.-M. Mitochondria-Targeted Curcumin: A Potent Antibacterial Agent against Methicillin-Resistant Staphylococcus aureus with a Possible Intracellular ROS Accumulation as the Mechanism of Action. Antibiotics 2023, 12, 401. https://doi.org/10.3390/antibiotics12020401
Leferman C-E, Stoica L, Stoica BA, Ciubotaru AD, Badescu AC, Bogdanici C-M, Neagu TP, Ghiciuc C-M. Mitochondria-Targeted Curcumin: A Potent Antibacterial Agent against Methicillin-Resistant Staphylococcus aureus with a Possible Intracellular ROS Accumulation as the Mechanism of Action. Antibiotics. 2023; 12(2):401. https://doi.org/10.3390/antibiotics12020401
Chicago/Turabian StyleLeferman, Carmen-Ecaterina, Laura Stoica, Bogdan Alexandru Stoica, Alin Dumitru Ciubotaru, Aida Corina Badescu, Camelia-Margareta Bogdanici, Tiberiu Paul Neagu, and Cristina-Mihaela Ghiciuc. 2023. "Mitochondria-Targeted Curcumin: A Potent Antibacterial Agent against Methicillin-Resistant Staphylococcus aureus with a Possible Intracellular ROS Accumulation as the Mechanism of Action" Antibiotics 12, no. 2: 401. https://doi.org/10.3390/antibiotics12020401