

Emergence and Evolution of OXA-23-Producing ST46Pas-ST462Oxf-KL28-OCL1 Carbapenem-Resistant Acinetobacter baumannii Mediated by a Novel ISAba1-Based Tn7534 Transposon
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility Profiles, Resistance Determinants and Virulence Factors
2.2. Transfer of Carbapenemase Resistance Determinants
2.3. Multilocus Sequence Typing (MLST), Capsular Polysaccharide (KL) and Lipoolygosaccharide (OCL)
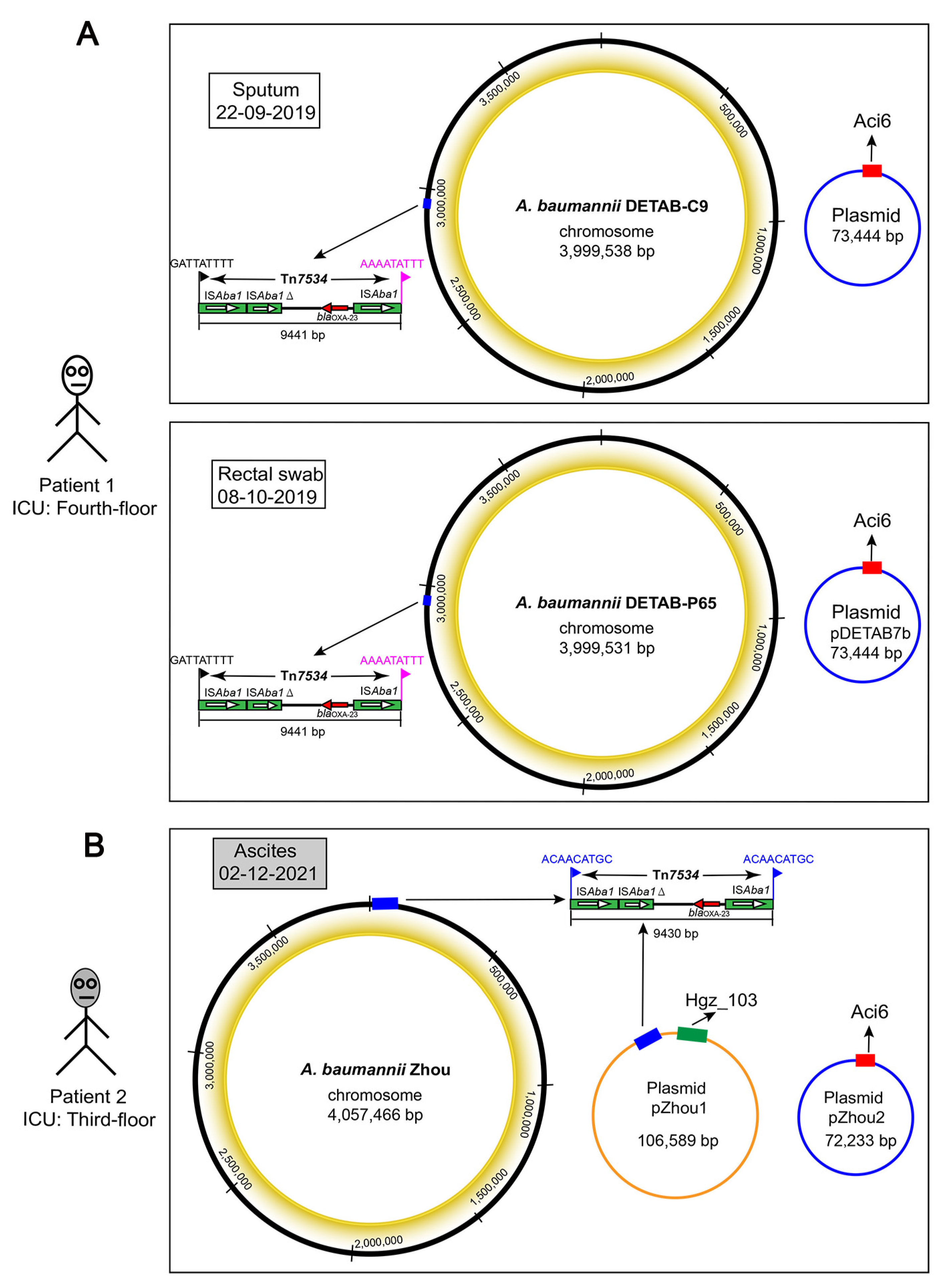
2.4. Chromosome Analysis of ST46Pas Strains
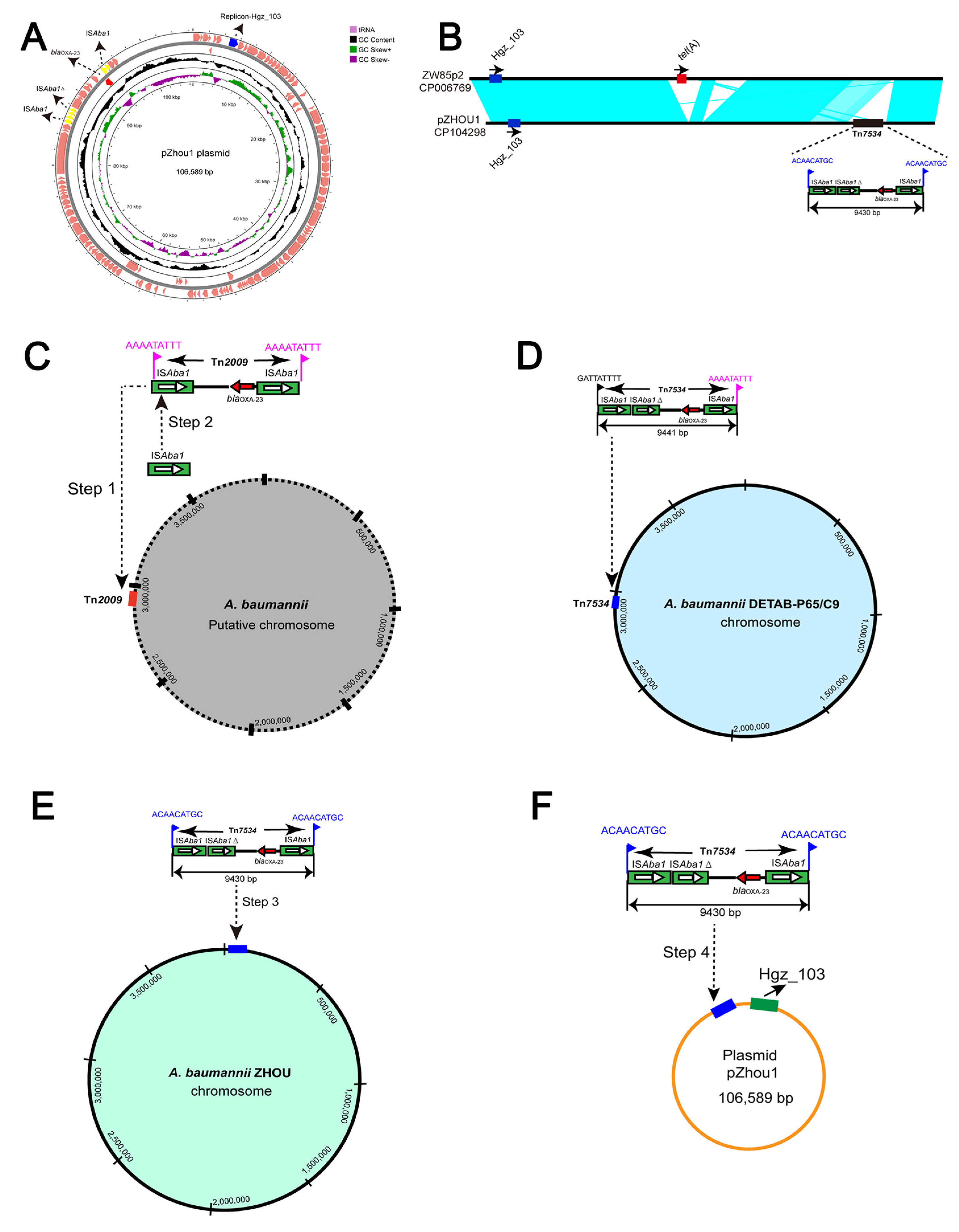
2.5. Genetic Analysis of Plasmids
2.6. Plasmid Comparison and the Evolution of ST46Pas CRAB Mediated by a Novel Tn7534 Transposon
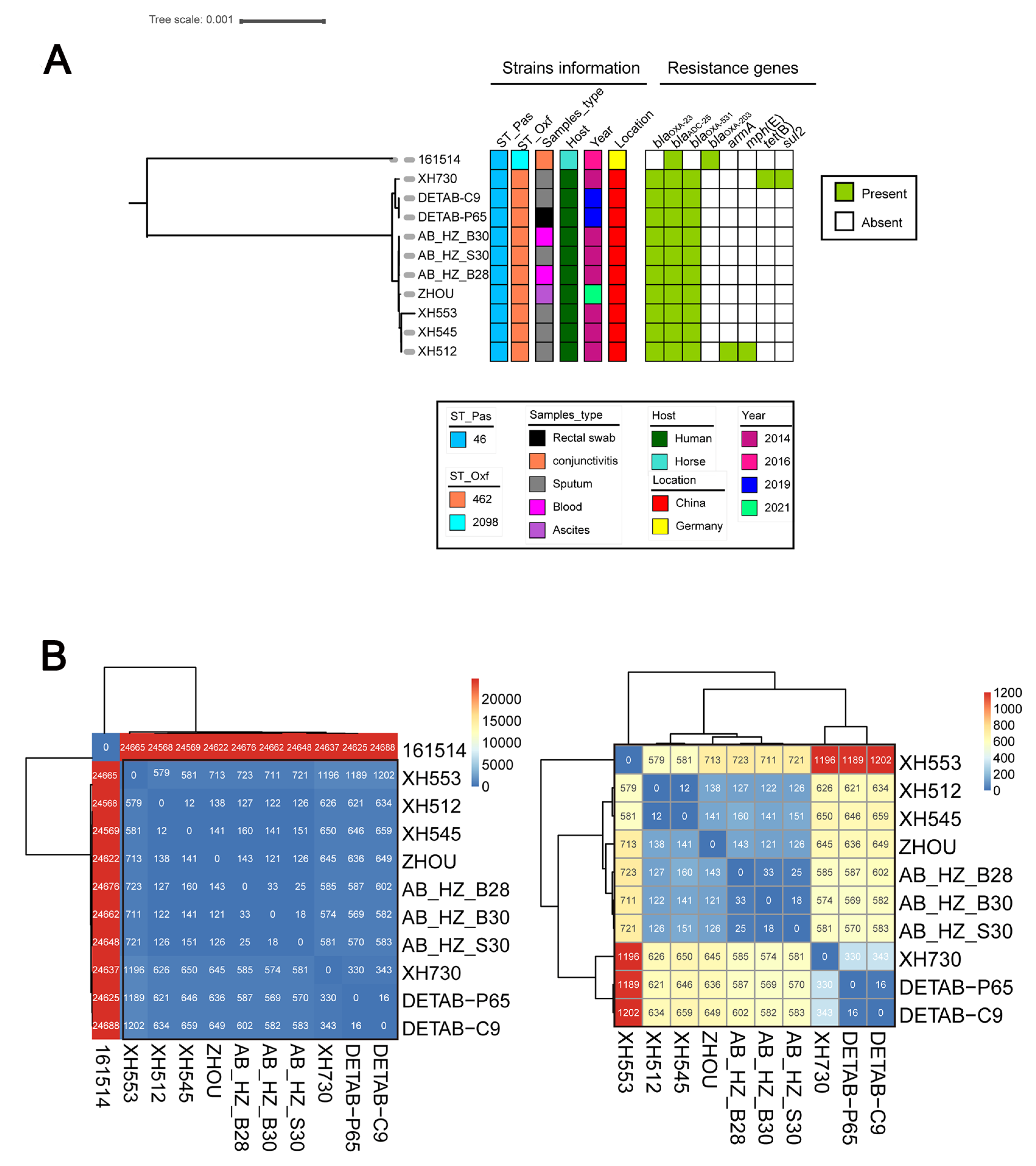
2.7. Phylogenetic Analysis of All ST46Pas A. baumannii Strains from NCBI Database
2.8. Pan-Genome and Single Nucleotide Polymorphisms (SNPs) Analysis
3. Discussion
4. Materials and Methods
4.1. Patient Information, Bacterial Isolation and Identification
4.2. Antimicrobial Agent Susceptibility Testing
4.3. Conjugation and Chemical Transformation Experiments
4.4. Whole-Genome Sequencing (WGS) and Phylogenetic Analysis
4.5. Pan-Genome Analysis
4.6. Nucleotide Sequence Accession Numbers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, H.; Moran, R.A.; Chen, Y.; Doughty, E.L.; Hua, X.; Jiang, Y.; Xu, Q.; Zhang, L.; Blair, J.M.A.; McNally, A.; et al. Transferable Acinetobacter baumannii plasmid pDETAB2 encodes OXA-58 and NDM-1 and represents a new class of antibiotic resistance plasmids. J. Antimicrob. Chemother. 2021, 76, 1130–1134. [Google Scholar] [CrossRef]
- Hamidian, M.; Nigro, S.J. Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii. Microb. Genom. 2019, 5, e000306. [Google Scholar] [CrossRef]
- Rando, E.; Segala, F.V.; Vargas, J.; Seguiti, C.; De Pascale, G.; Murri, R.; Fantoni, M. Cefiderocol for Severe Carbapenem-Resistant A. baumannii Pneumonia: Towards the Comprehension of Its Place in Therapy. Antibiotics 2021, 11, 3. [Google Scholar] [CrossRef]
- Perez, S.; Innes, G.K.; Walters, M.S.; Mehr, J.; Arias, J.; Greeley, R.; Chew, D. Increase in Hospital-Acquired Carbapenem-Resistant Acinetobacter baumannii Infection and Colonization in an Acute Care Hospital During a Surge in COVID-19 Admissions—New Jersey, February-July 2020. Morb. Mortal. Wkly. Rep. 2020, 69, 1827–1831. [Google Scholar] [CrossRef]
- Liepa, R.; Mann, R.; Osman, M.; Hamze, M.; Gunawan, C.; Hamidian, M. Cl415, a carbapenem-resistant Acinetobacter baumannii isolate containing four AbaR4 and a new variant of AbGRI2, represents a novel global clone 2 strain. J. Antimicrob. Chemother. 2022, 77, 345–350. [Google Scholar] [CrossRef]
- Kim, D.H.; Choi, J.Y.; Kim, H.W.; Kim, S.H.; Chung, D.R.; Peck, K.R.; Thamlikitkul, V.; So, T.M.; Yasin, R.M.; Hsueh, P.R.; et al. Spread of carbapenem-resistant Acinetobacter baumannii global clone 2 in Asia and AbaR-type resistance islands. Antimicrob. Agents Chemother. 2013, 57, 5239–5246. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.L.; Ji, S.J.; Ruan, Z.; Fu, Y.; Fu, Y.Q.; Wang, Y.F.; Yu, Y.S. Dissemination of blaOXA-23 in Acinetobacter spp. in China: Main roles of conjugative plasmid pAZJ221 and transposon Tn2009. Antimicrob. Agents Chemother. 2015, 59, 1998–2005. [Google Scholar] [CrossRef] [Green Version]
- Fedrigo, N.H.; Xavier, D.E.; Cerdeira, L.; Fuga, B.; Marini, P.V.B.; Shinohara, D.R.; Carrara-Marroni, F.E.; Lincopan, N.; Tognim, M.C.B. Genomic insights of Acinetobacter baumannii ST374 reveal wide and increasing resistome and virulome. Infect. Genet. Evol. 2022, 97, 105148. [Google Scholar] [CrossRef]
- Lee, H.-Y.; Chang, R.-C.; Su, L.-H.; Liu, S.-Y.; Wu, S.-R.; Chuang, C.-H.; Chen, C.-L.; Chiu, C.-H. Wide spread of Tn2006 in an AbaR4-type resistance island among carbapenem-resistant Acinetobacter baumannii clinical isolates in Taiwan. Int. J. Antimicrob. Agents 2012, 40, 163–167. [Google Scholar] [CrossRef]
- Zhao, F.; Liu, H.; Yao, Y.; Zhang, L.; Zhou, Z.; Leptihn, S.; Yu, Y.; Hua, X.; Fu, Y. Description of a Rare Pyomelanin-Producing Carbapenem-Resistant Acinetobacter baumannii Strain Coharboring Chromosomal OXA-23 and NDM-1. Microbiol. Spectr. 2022, 10, e0214422. [Google Scholar] [CrossRef]
- Hua, X.; Xu, Q.; Zhou, Z.; Ji, S.; Yu, Y. Relocation of Tn2009 and characterization of an ABGRI3-2 from re-sequenced genome sequence of Acinetobacter baumannii MDR-ZJ06. J. Antimicrob. Chemother. 2019, 74, 1153–1155. [Google Scholar] [CrossRef]
- Wareth, G.; Abdel-Glil, M.Y.; Schmoock, G.; Steinacker, U.; Kaspar, H.; Neubauer, H.; Sprague, L.D. Draft Genome Sequence of an Acinetobacter baumannii Isolate Recovered from a Horse with Conjunctivitis in Germany. Microbiol. Resour. Announc. 2019, 8, e01128-19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xie, J.; Li, H.; Wang, Z.; Yin, Y.; Wang, S.; Chen, H.; Wang, Q.; Wang, H. Genomic and Phenotypic Evolution of Tigecycline-Resistant Acinetobacter baumannii in Critically Ill Patients. Microbiol. Spectr. 2022, 10, e0159321. [Google Scholar] [CrossRef]
- Schultz, M.B.; Pham Thanh, D.; Tran Do Hoan, N.; Wick, R.R.; Ingle, D.J.; Hawkey, J.; Edwards, D.J.; Kenyon, J.J.; Phu Huong Lan, N.; Campbell, J.I.; et al. Repeated local emergence of carbapenem-resistant Acinetobacter baumannii in a single hospital ward. Microb. Genom. 2016, 2, e000050. [Google Scholar] [CrossRef] [Green Version]
- Douraghi, M.; Kenyon, J.J.; Aris, P.; Asadian, M.; Ghourchian, S.; Hamidian, M. Accumulation of Antibiotic Resistance Genes in Carbapenem-Resistant Acinetobacter baumannii Isolates Belonging to Lineage 2, Global Clone 1, from Outbreaks in 2012–2013 at a Tehran Burns Hospital. mSphere 2020, 5, e00164-20. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wu, Z.; Mao, C.; Guo, G.; Zeng, Z.; Fei, Y.; Wan, S.; Peng, J.; Wu, J. Antimicrobial Peptide Cec4 Eradicates the Bacteria of Clinical Carbapenem-Resistant Acinetobacter baumannii Biofilm. Front. Microbiol. 2020, 11, 1532. [Google Scholar] [CrossRef]
- Islam, M.M.; Kim, K.; Lee, J.C.; Shin, M. LeuO, a LysR-Type Transcriptional Regulator, Is Involved in Biofilm Formation and Virulence of Acinetobacter baumannii. Front. Cell. Infect. Microbiol. 2021, 11, 738706. [Google Scholar] [CrossRef]
- Moran, R.A.; Liu, H.; Doughty, E.L.; Hua, X.; Cummins, E.A.; Liveikis, T.; McNally, A.; Zhou, Z.; van Schaik, W.; Yu, Y. GR13-type plasmids in Acinetobacter potentiate the accumulation and horizontal transfer of diverse accessory genes. Microb. Genom. 2022, 8, 000840. [Google Scholar] [CrossRef]
- Wyres, K.L.; Cahill, S.M.; Holt, K.E.; Hall, R.M.; Kenyon, J.J. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 2020, 6, e000339. [Google Scholar] [CrossRef]
- Gorbunova, V.; Seluanov, A.; Mita, P.; McKerrow, W.; Fenyö, D.; Boeke, J.D.; Linker, S.B.; Gage, F.H.; Kreiling, J.A.; Petrashen, A.P.; et al. The role of retrotransposable elements in ageing and age-associated diseases. Nature 2021, 596, 43–53. [Google Scholar] [CrossRef]
- Hua, X.; Liang, Q.; Deng, M.; He, J.; Wang, M.; Hong, W.; Wu, J.; Lu, B.; Leptihn, S.; Yu, Y.; et al. BacAnt: A Combination Annotation Server for Bacterial DNA Sequences to Identify Antibiotic Resistance Genes, Integrons, and Transposable Elements. Front. Microbiol. 2021, 12, 649969. [Google Scholar] [CrossRef] [PubMed]
- Harmer, C.J.; Hall, R.M. IS 26 Family Members IS 257 and IS 1216 Also Form Cointegrates by Copy-In and Targeted Conservative Routes. mSphere 2020, 5, e00811-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmer, C.J.; Hall, R.M. IS 26-Mediated Formation of Transposons Carrying Antibiotic Resistance Genes. mSphere 2016, 1, e00038-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Chen, J.; Shen, H.; Chen, Z.; Yang, Q.W.; Zhu, J.; Li, X.; Yang, Q.; Zhao, F.; Ji, J.; et al. Emergence of Ceftazidime- and Avibactam-Resistant Klebsiella pneumoniae Carbapenemase-Producing Pseudomonas aeruginosa in China. mSystems 2021, 6, e0078721. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Liu, D.; Song, H.; Zhang, P.; Li, R.; Fu, Y.; Liu, X.; Jia, J.; Li, X.; Fanning, S.; et al. Novel IS26-mediated hybrid plasmid harbouring tet(X4) in Escherichia coli. J. Glob. Antimicrob. Resist. 2020, 21, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, G.A.; Hall, R.M. The tet39 Determinant and the msrE-mphE Genes in Acinetobacter Plasmids Are Each Part of Discrete Modules Flanked by Inversely Oriented p dif (XerC-XerD) Sites. Antimicrob. Agents Chemother. 2017, 61, e00780-17. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, M.M.; Giani, T.; D’Arezzo, S.; Capone, A.; Petrosillo, N.; Visca, P.; Luzzaro, F.; Rossolini, G.M. Characterization of pABVA01, a plasmid encoding the OXA-24 carbapenemase from Italian isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 3528–3533. [Google Scholar] [CrossRef] [Green Version]
- Loh, B.; Chen, J.; Manohar, P.; Yu, Y.; Hua, X.; Leptihn, S. A Biological Inventory of Prophages in A. baumannii Genomes Reveal Distinct Distributions in Classes, Length, and Genomic Positions. Front. Microbiol. 2020, 11, 579802. [Google Scholar] [CrossRef]
- Krahn, T.; Wibberg, D.; Maus, I.; Winkler, A.; Bontron, S.; Sczyrba, A.; Nordmann, P.; Pühler, A.; Poirel, L.; Schlüter, A. Intraspecies Transfer of the Chromosomal Acinetobacter baumannii bla NDM-1 Carbapenemase Gene. Antimicrob. Agents Chemother. 2016, 60, 3032–3040. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.-C.; Chen, C.-C.; Lu, Y.-C.; Chuang, Y.-C.; Tang, H.-J. In vitro activity of cefoperazone and cefoperazone-sulbactam against carbapenem-resistant Acinetobacter baumannii and Pseudomonas aeruginosa. Infect. Drug Resist. 2019, 12, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Weng, X.; Shi, Q.; Wang, S.; Shi, Y.; Sun, D.; Yu, Y. The Characterization of OXA-232 Carbapenemase-Producing ST437 Klebsiella pneumoniae in China. Can. J. Infect. Dis. Med. Microbiol. 2020, 2020, 5626503. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Diancourt, L.; Passet, V.; Nemec, A.; Dijkshoorn, L.; Brisse, S. The population structure of Acinetobacter baumannii: Expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS ONE 2010, 5, e10034. [Google Scholar] [CrossRef] [Green Version]
- Bartual, S.G.; Seifert, H.; Hippler, C.; Luzon, M.A.D.; Wisplinghoff, H.; Rodriguez-Valera, F. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4382–4390. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Liang, Q.; Liu, C.; Xu, R.; Song, M.; Zhou, Z.; Li, H.; Dai, W.; Yang, M.; Yu, Y.; Chen, H. fIDBAC: A Platform for Fast Bacterial Genome Identification and Typing. Front. Microbiol. 2021, 12, 723577. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [Green Version]
- Tansirichaiya, S.; Rahman, M.A.; Roberts, A.P. The Transposon Registry. Mob. DNA 2019, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, Y.; Liu, M.; Tai, C.; Sun, J.; Deng, Z.; Ou, H.-Y. oriTfinder: A web-based tool for the identification of origin of transfers in DNA sequences of bacterial mobile genetic elements. Nucleic Acids Res. 2018, 46, W229–W234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, X.; Liang, Q.; Fang, L.; He, J.; Wang, M.; Hong, W.; Leptihn, S.; Wang, H.; Yu, Y.; Chen, H. Bautype: Capsule and Lipopolysaccharide Serotype Prediction for Acinetobacter baumannii Genome. Infectious Microbes & Diseases. 2020, 2, 18–25. [Google Scholar] [CrossRef]
- Shao, M.; Ying, N.; Liang, Q.; Ma, N.; Leptihn, S.; Yu, Y.; Chen, H.; Liu, C.; Hua, X. Pdif-mediated antibiotic resistance genes transfer in bacteria identified by pdifFinder. Brief Bioinform 2023, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A fast phage search tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Bush, S.J. Generalizable characteristics of false-positive bacterial variant calls. Microb. Genom. 2021, 7, 000615. [Google Scholar] [CrossRef]
- Feng, Y.; Zou, S.M.; Chen, H.F.; Yu, Y.S.; Ruan, Z. BacWGSTdb 2.0: A one-stop repository for bacterial whole-genome sequence typing and source tracking. Nucleic Acids Res. 2021, 49, D644–D650. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpi, F.M.; Coman, M.M.; Silvi, S.; Picciolini, M.; Verdenelli, M.C.; Napolioni, V. Comprehensive pan-genome analysis of Lactiplantibacillus plantarum complete genomes. J. Appl. Microbiol. 2022, 132, 592–604. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Isolates | Antibiotics 1 Minimum Inhibitory Concentration (mg/L) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| IMP | MEM | CAZ | SCF (1:1) | SCF (2:1) | AMI | CIP | COL | TGC | |
| DETAB-C9 | 16 | 32 | 4 | 32 | 64 | 8 | >32 | 1 | 1 |
| DETAB-P65 | 8 | 16 | 4 | 32 | 64 | 8 | >32 | 1 | 0.5 |
| ZHOU | 32 | 64 | 4 | 32 | 64 | 8 | >32 | 1 | 0.5 |
| Isolates | Element | Replicon | Size (bp) | GC (%) | Antibiotic Resistance Genes |
|---|---|---|---|---|---|
| DETAB-C9 | chromosome | ND | 3,999,538 | 38.98% | blaOXA-23, blaOXA-67, blaADC-26, ant(3″)-IIa |
| pDETAB-C9-1 | Aci6 | 73,444 | 33.40% | Not detected | |
| DETAB-P65 | chromosome | ND | 3,999,531 | 38.98% | blaOXA-23, blaOXA-67, blaADC-26, ant(3″)-IIa |
| pDETAB7b | Aci6 | 73,444 | 33.40% | Not detected | |
| ZHOU | chromosome | ND | 4,057,466 | 39.02% | blaOXA-23, blaOXA-67, blaADC-26, ant(3″)-IIa |
| pZHOU-1 | Hgz_103 | 106,589 | 41.38% | blaOXA-23 | |
| pZHOU-2 | Aci6 | 72,233 | 33.50% | Not detected |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Liu, X.; He, J.; Zhang, L.; Zhao, F.; Zhou, Z.; Hua, X.; Yu, Y. Emergence and Evolution of OXA-23-Producing ST46Pas-ST462Oxf-KL28-OCL1 Carbapenem-Resistant Acinetobacter baumannii Mediated by a Novel ISAba1-Based Tn7534 Transposon. Antibiotics 2023, 12, 396. https://doi.org/10.3390/antibiotics12020396
Liu H, Liu X, He J, Zhang L, Zhao F, Zhou Z, Hua X, Yu Y. Emergence and Evolution of OXA-23-Producing ST46Pas-ST462Oxf-KL28-OCL1 Carbapenem-Resistant Acinetobacter baumannii Mediated by a Novel ISAba1-Based Tn7534 Transposon. Antibiotics. 2023; 12(2):396. https://doi.org/10.3390/antibiotics12020396
Chicago/Turabian StyleLiu, Haiyang, Xiaochen Liu, Jintao He, Linghong Zhang, Feng Zhao, Zhihui Zhou, Xiaoting Hua, and Yunsong Yu. 2023. "Emergence and Evolution of OXA-23-Producing ST46Pas-ST462Oxf-KL28-OCL1 Carbapenem-Resistant Acinetobacter baumannii Mediated by a Novel ISAba1-Based Tn7534 Transposon" Antibiotics 12, no. 2: 396. https://doi.org/10.3390/antibiotics12020396