
Association of Antibacterial Susceptibility Profile with the Prevalence of Genes Encoding Efflux Proteins in the Bangladeshi Clinical Isolates of Staphylococcus aureus
 ,
,
Abstract
:1. Introduction
2. Materials & Methods
2.1. Chemicals
2.2. Bacterial Strain Collection
2.3. Bacterial Identification
2.4. Bacterial Cell Culture
2.5. Antibacterial Susceptibility Test
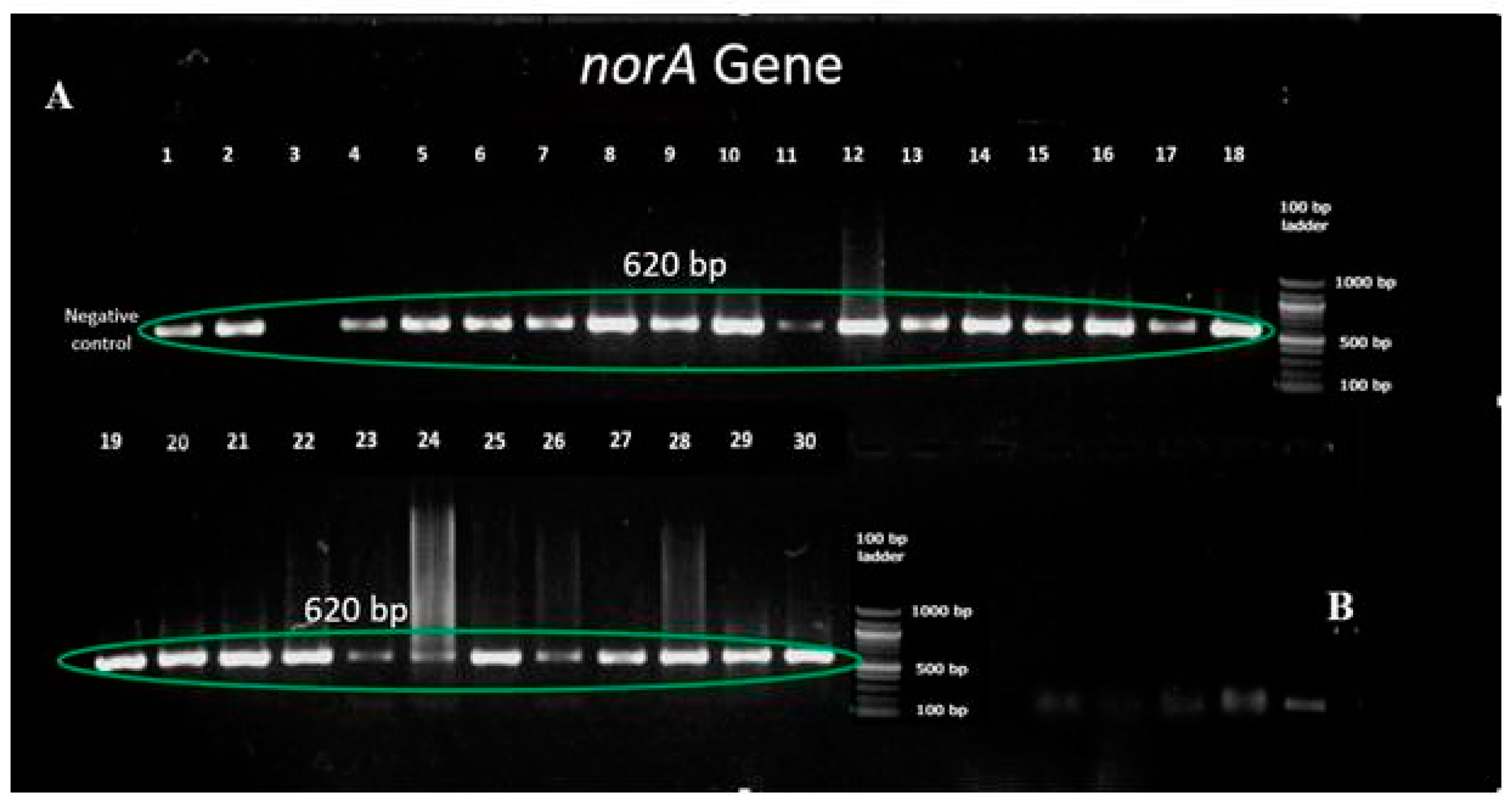
2.6. Amplification of Target Genes by Polymerase Chain Reaction
3. Result
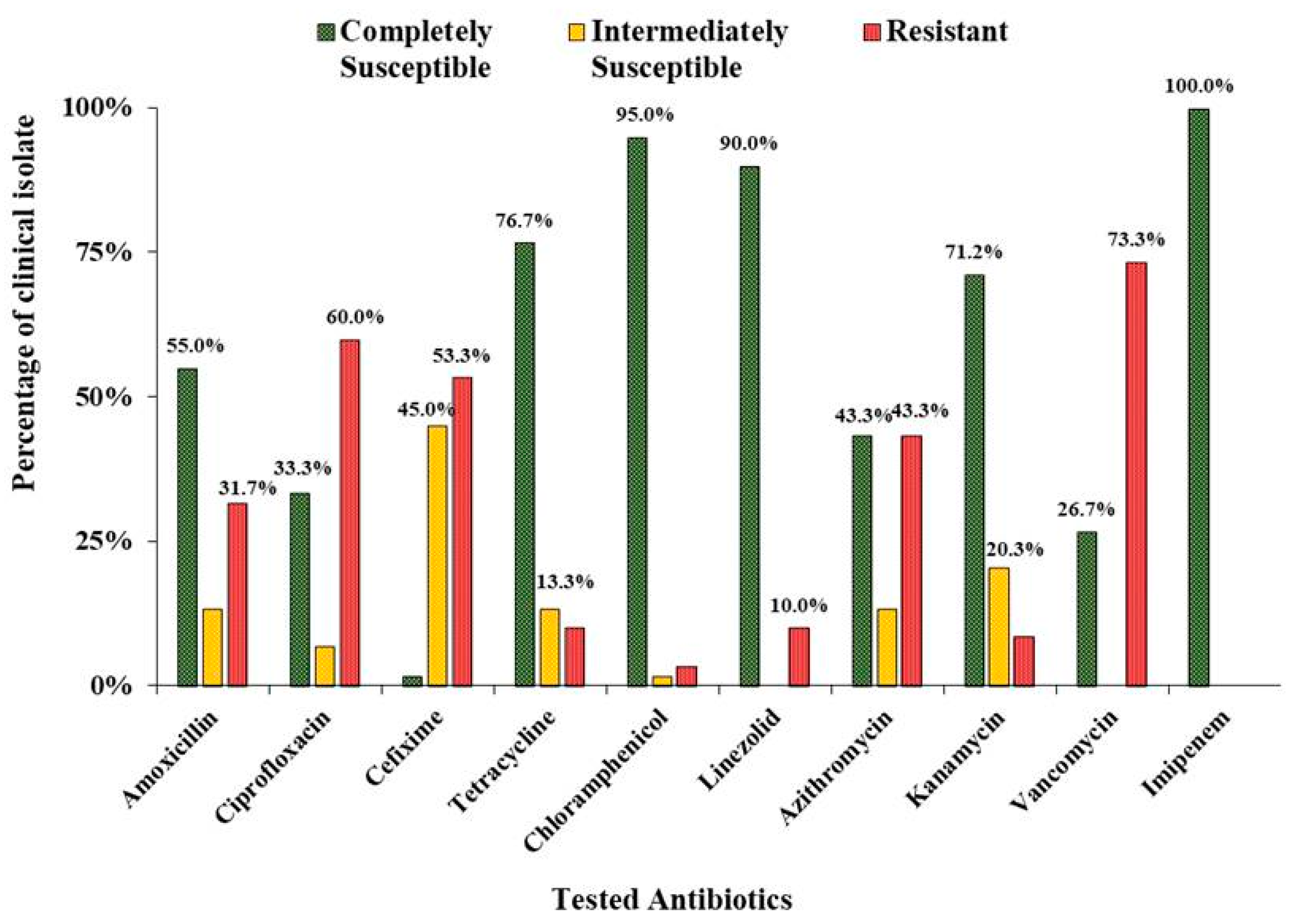
3.1. Antibacterial Susceptibility Pattern in Clinical Isolates of S. aureus
3.2. Characterizations of Different Types of S. aureus in Clinical Isolates
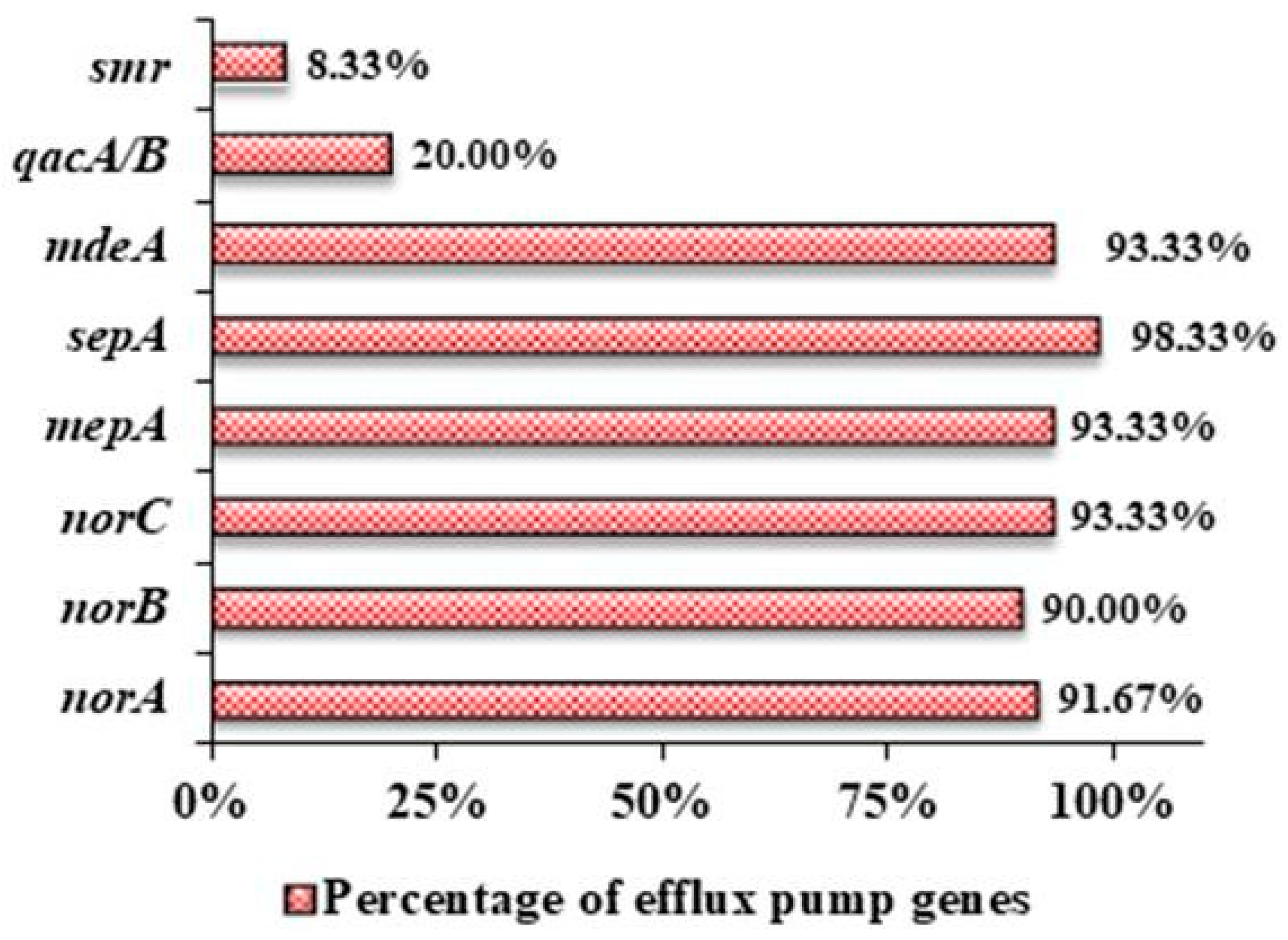
3.3. Prevalence of Different Efflux Pump Genes in the Clinical Isolates of S. aureus
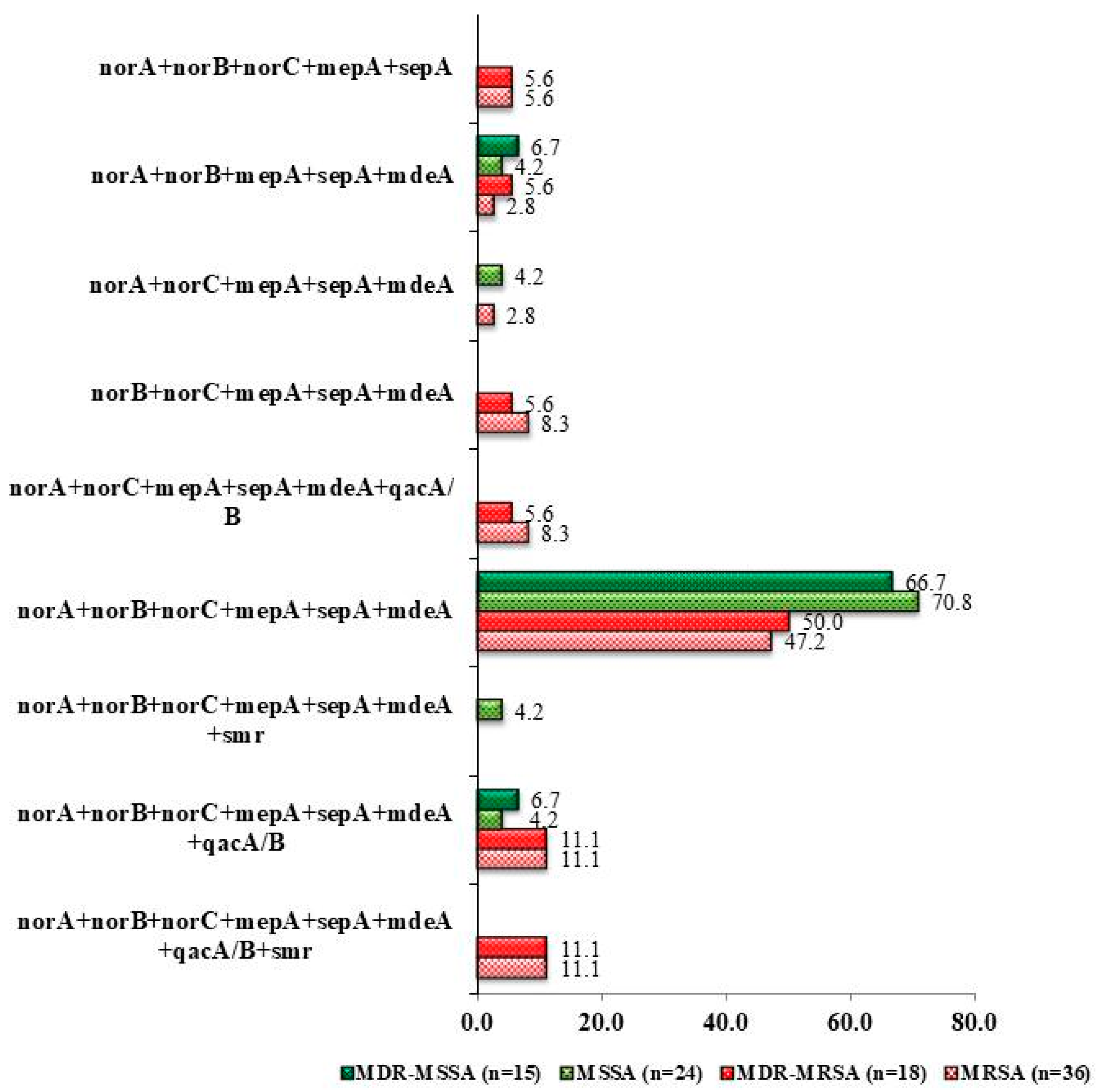
3.4. Antibiotic Resistant Profile Associated with Efflux Pump Genes Combination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okeke, I.N.; Lamikanra, A. Export of antimicrobial drugs by West African travelers. J. Travel Med. 2006, 10, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.P.; Pearson, A.; Duckworth, G. Surveillance and epidemiology of MRSA bacteraemia in the UK. J. Antimicrob. Chemother. 2005, 56, 455–462. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Point Prevalence Survey of Healthcare-Associated Infections and Antimicrobial Use in European Acute Care Hospitals: 2011–2012; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2013. [Google Scholar]
- World Health Organization. WHO’s First Global Report on Antibiotic Resistance Reveals Serious, Worldwide Threat to Public Health. World Health Organization: Geneva, Switzerland. Available online: https://www.who.int/news/item/30-04-2014-who-s-first-global-report-on-antibiotic-resistance-reveals-serious-worldwide-threat-to-public-health (accessed on 5 August 2022).
- Abdulgader, S.M.; Shittu, A.O.; Nicol, M.P.; Kaba, M. Molecular epidemiology of Methicillin-resistant Staphylococcus aureus in Africa: A systematic review. Front. Microbiol. 2015, 6, 348. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Mahmud, M.; Chowdhury, M.; Hakim, M. Prevalence of multidrug resistant Staphylococcus aureus isolates in clinical specimens collected from local patients of Chittagong, Bangladesh. Chittagong Univ. J. Sci. 2013, 6, 175–185. [Google Scholar] [CrossRef]
- Haq, J.A.; Rahman, M.M.; Asna, S.M.Z.H.; Hossain, M.A.; Ahmed, I.; Haq, T.; Morshed, M.A.H.G. Methicillin-resistant Staphylococcus aureus in Bangladesh—A multicentre study. Int. J. Antimicrob. Agents 2005, 25, 276–277. [Google Scholar] [CrossRef]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug efflux pumps in Staphylococcus aureus: An Update. Open Microbio. J. 2013, 7, 59–71. [Google Scholar] [CrossRef]
- Piddock, L.J.V. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef]
- Vali, L.; Davies, S.E.; Lai, L.L.G.; Dave, J.; Amyes, S.G.B. Frequency of biocide resistance genes, antibiotic resistance and the effect of chlorhexidine exposure on clinical methicillin-resistant Staphylococcus aureus isolates. J. Antimicrob. Chemother. 2008, 61, 524–532. [Google Scholar] [CrossRef]
- Hassanzadeh, S.; Mashhadi, R.; Yousefi, M.; Askari, E.; Saniei, M.; Pourmand, M.R. Frequency of efflux pump genes mediating ciprofloxacin and antiseptic resistance in methicillin-resistant Staphylococcus aureus isolates. Microb. Pathog. 2017, 111, 71–74. [Google Scholar] [CrossRef]
- Watkins, R.R.; David, M.Z.; Salata, R.A. Current concepts on the virulence mechanisms of meticillin-resistant Staphylococcus aureus. J. Med. Microbiol. 2012, 61, 1179–1193. [Google Scholar] [CrossRef] [PubMed]
- Uhlemann, A.C.; Otto, M.; Lowy, F.D.; DeLeo, F.R. Evolution of community- and healthcare-associated methicillin-resistant Staphylococcus aureus. Infect. Genet. Evol. 2014, 21, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Parvez MA, K.; Ferdous, R.N.; Rahman, M.S.; Islam, S. Healthcare-associated (HA) and community-associated (CA) methicillin resistant Staphylococcus aureus (MRSA) in Bangladesh—Source, diagnosis and treatment. J. Genet. Eng. Biotechnol. 2018, 16, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Islam, T.; Kubra, K.; Chowdhury, M.M.H. Prevalence of Methicillin-Resistant Staphylococcus aureus in hospitals in Chittagong, Bangladesh: A threat of nosocomial infection. J. Microsc. Ultrastruct. 2018, 6, 188–191. [Google Scholar]
- Dulon, M.; Haamann, F.; Peters, C.; Schablon, A.; Nienhaus, A. MRSA prevalence in European healthcare settings: A review. BMC Infect. Dis. 2011, 11, 138. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.Z.; Naher, A.; Hasan, P.; Mozazfia, K.T.; Tasnim, H.; Ferdush, Z.; Towhid, K.M.; Simran, M.A.A. Prevalent bacteria and their sensitivity and resistance pattern to antibiotics: A study in Dhaka Medical College Hospital. J. Dhaka Med. Coll. 2017, 26, 52–64. [Google Scholar] [CrossRef]
- McGuinness, W.A.; Malachowa, N.; Deleo, F.R. Vancomycin resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
- George, S.K.; Suseela, M.R.; Safi, S.E.; Elnagi, E.A.; Al-naam, Y.A.; Adam, A.A.M.; Jacob, A.M.; Al-Maqati, T.; KS, H.K. Molecular determination of van genes among clinical isolates of enterococci at a hospital setting. Saudi J. Biol. Sci. 2021, 28, 2895–2899. [Google Scholar] [CrossRef]
- Kaatz, G.W.; Seo, S.M. Inducible NorA-mediated multidrug resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1995, 39, 2650–2655. [Google Scholar] [CrossRef]
- Truong-Bolduc, Q.C.; Dunman, P.M.; Strahilevitz, J.; Projan, S.J.; Hooper, D.C. MgrA is a multiple regulator of two new efflux pumps in Staphylococcus aureus. J. Bacteriol. 2005, 187, 2395–2405. [Google Scholar] [CrossRef]
- Truong-Bolduc, Q.C.; Strahilevitz, J.; Hooper, D.C. NorC, a new efflux pump regulated by MgrA of Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 1104–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaatz, G.W.; McAleese, F.; Seo, S.M. Multidrug Resistance in Staphylococcus aureus due to overexpression of a novel multidrug and toxin extrusion (MATE) transport protein. Antimicrob. Agents Chemother. 2005, 49, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Narui, K.; Noguchi, N.; Wakasugi, K.; Sasatsu, M. Cloning and characterization of a novel chromosomal drug efflux gene in Staphylococcus aureus. Biol. Pharm. Bull. 2002, 25, 1533–1536. [Google Scholar] [CrossRef]
- Huang, J.; O’Toole, P.W.; Shen, W.; Amrine-Madsen, H.; Jiang, X.; Lobo, N.; Palmer, L.M.; Voelker, L.; Fan, F.; Gwynn, M.N.; et al. Novel chromosomally encoded multidrug efflux transporter MdeA in Staphylococcus aureus. Antimicrob. Agents Chemother. 2004, 48, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh, S.; Ganjloo, S.; Pourmand, M.R.; Mashhadi, R.; Ghazvini, K. Epidemiology of efflux pumps genes mediating resistance among Staphylococcus aureus: A systematic review. Microb. Pathog. 2020, 139, 103850. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, H.; Han, L.; Shu, W.; Wu, Q.; Ni, Y. Frequency of biocide-resistant genes and susceptibility to chlorhexidine in high-level mupirocin-resistant, methicillin-resistant Staphylococcus aureus (MuH MRSA). Diagn. Microbiol. Infect. Dis. 2015, 82, 278–283. [Google Scholar] [CrossRef]
- Shamkhi, G.J.; Saadedin, S.M.K.; Jassim, K.A. Detection the prevalence of some chromosomal efflux pump genes in Methicillin resistant Staphylococcus aureus isolated from Iraqi patients. Iraqi J. Biotechnol. 2019, 18, 33–42. [Google Scholar]
- Antiabong, J.F.; Kock, M.M.; Mbelle, N.M.; Ehlers, M.M. Diversity of multidrug efflux genes and phenotypic evaluation of the in vitro resistance dynamics of clinical Staphylococcus aureus isolates using methicillin; a model β-lactam. Open Microb. J. 2017, 11, 132–141. [Google Scholar] [CrossRef]
- Conceiçãoa, T.; Lencastrea, H.D.; Aires-de-Sousaa, M. Prevalence of biocide resistance genes and chlorhexidine and mupirocin non-susceptibility in Portuguese hospitals during a 31-year period (1985–2016). J. Glob. Antimicrob. Resist. 2021, 24, 169–174. [Google Scholar] [CrossRef]
- Shamsudin, M.N.; Alreshidi, M.A.; Hamat, R.A.; Alshrari, A.S.; Atshan, S.S.; Neela, V. High prevalence of qacA/B carriage among clinical isolates of meticillin-resistant Staphylococcus aureus in Malaysia. J. Hosp. Infect. 2012, 81, 206–208. [Google Scholar] [CrossRef]
- Longtin, J.; Seah, S.; Siebert, K.; McGeer, A.; Simor, A.; Longtin, Y.; Low, D.E.; Melano, R.G. Distribution of antiseptic resistance genes qacA, qacB, and smr in methicillin-resistant Staphylococcus aureus isolated in Toronto, Canada, from 2005 to 2009. Antimicrob. Agents Chemother. 2011, 55, 2999–3001. [Google Scholar] [CrossRef] [PubMed]
- Conceição, T.; Coelho, C.; Lencastre, H.D.; Aires-de-Sousa, M. High prevalence of biocide resistance determinants in Staphylococcus aureus isolates from three African countries. Antimicrob. Agents Chemother. 2016, 60, 678–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, W.D.; Berger-Bächi, B.; Kayser, F.H. Additional DNA in methicillin-resistant Staphylococcus aureus and molecular cloning of mec-specific DNA. J. Bacteriol. 1986, 165, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Oligonucleotide Sequence | %GC Content | Tm (°C) | Amplicon Size | |
|---|---|---|---|---|---|
| norA | Forward | 5′-TTCACCAAGCCATCAAAAAG-3′ | 40 | 54.3 | 620 bp |
| Reverse | 5′-CTTGCCTTTCTCCAGCAATA-3′ | 45 | 56.4 | ||
| norB | Forward | 5′-AGCGCGTTGTCTATCTTTCC-3′ | 50 | 58.4 | 213 bp |
| Reverse | 5′-GCAGGTGGTCTTGCTGATAA-3′ | 50 | 58.4 | ||
| norC | Forward | 5′-AATGGGTTCTAAGCGACCAA-3′ | 45 | 56.4 | 216 bp |
| Reverse | 5′-ATACCTGAAGCAACGCCAAC-3′ | 50 | 58.4 | ||
| mepA | Forward | 5′-ATGTTGCTGCTGCTCTGTTC-3′ | 50 | 58.4 | 718 bp |
| Reverse | 5′-TCAACTGTCAAACGATCACG-3′ | 45 | 56.4 | ||
| sepA | Forward | 5′-GCAGTCGAGCATTTAATGGA-3′ | 45 | 56.4 | 103 bp |
| Reverse | 5′-ACGTTGTTGCAACTGTGTAAGA-3′ | 41 | 58.4 | ||
| mdeA | Forward | 5′-AACGCGATACCAACCATTC-3′ | 47 | 55.2 | 677 bp |
| Reverse | 5′-TTAGCACCAGCTATTGGACCT-3′ | 48 | 59.4 | ||
| qacA/B | Forward | 5′-GCTGCATTTATGACAATGTTTG-3′ | 36 | 56.6 | 628 bp |
| Reverse | 5′-AATCCCACCTACTAAAGCAG-3′ | 45 | 56.4 | ||
| smr | Forward | 5′-ATAAGTACTGAAGTTATTGGAAGT-3′ | 29 | 56.7 | 285 bp |
| Reverse | 5′-TTCCGAAAATGTTTAACGAAACTA-3′ | 29 | 56.7 | ||
| mecA | Forward | 5′-GTTGTAGTTGTCGGGTTTGG-3′ | 50 | 58.4 | 331 bp |
| Reverse | 5′-CTTCCACATACCATCTTCTTTAAC-3′ | 38 | 60.1 | ||
| Characterizations of Isolates | No. of Isolates | Percentage (%) | Characterizations of Isolates | No. of Isolates | Percentage (%) |
|---|---|---|---|---|---|
| Methicillin resistant S. aureus (MRSA) | 36 | 60% | MDR-MRSA | 18 | 30% |
| Regular-MRSA | 18 | 30% | |||
| Methicillin sensitive S. aureus(MSSA) | 24 | 40% | MDR-MSSA | 15 | 25% |
| Regular-MSSA | 9 | 15% | |||
| Multi-drug resistant S. aureus | 33 | 55% | Regular S. aureus | 27 | 45% |
| Status of Isolates | No. of Isolates | 4 Genes | 5 Genes | 6 Genes | 7 Genes | 8 Genes |
|---|---|---|---|---|---|---|
| MRSA | 36 | 36 | 35 | 27 | 7 | 3 |
| MDR MRSA | 18 | - | 18 | 14 | 4 | 2 |
| MSSA | 24 | 24 | 22 | 19 | 2 | - |
| MDR MSSA | 15 | 15 | 13 | 11 | 1 | - |
| Gene Combinations | AMX | CIP | CFM | AZM | VAN |
|---|---|---|---|---|---|
| MRSA (n = 36) | |||||
| Resistant isolates (n) | 11 | 21 | 22 | 13 | 25 |
| norA + norB + norC + mepA + sepA + mdeA | 3 | 9 | 8 | 6 | 13 |
| norA + norB + norC + mepA + sepA + mdeA + qacA/B | 2 | 2 | 4 * | 1 | 2 |
| norA + norB + norC + mepA + sepA + mdeA + qacA/B + smr | - | 2 | 3 | 2 | 2 |
| norA + norC + mepA + sepA + mdeA + qacA/B | 2 | 3 * | 2 | 2 | 2 |
| norB + norC + mepA + sepA + mdeA | 2 | 3 | 2 | 1 | 2 |
| norA + norB + norC + mepA + sepA | - | 2 * | 1 | - | 3 * |
| MDR-MRSA (n = 18) | |||||
| Resistant isolates (n) | 10 | 14 | 15 | 9 | 17 |
| norA + norB + norC + mepA + sepA + mdeA | 5 * | 9 * | 7 | 4 | 9 * |
| norA + norB + norC + mepA + sepA + mdeA + qacA/B | 1 | 2 * | 2 * | 2 * | 2 * |
| norA + norB + norC + mepA + sepA + mdeA + qacA/B + smr | - | 2 * | 2 * | 1 | 2 * |
| MSSA (n = 24) | |||||
| Resistant isolates (n) | 8 | 15 | 13 | 10 | 19 |
| norA + norB + norC + mepA + sepA + mdeA | 5 | 11 | 6 | 7 | 13 |
| norA + norB + norC + mepA + sepA + mdeA + qacA/B | 1 | - | 1 * | 1 * | 1 * |
| norB + norC + mepA + sepA + mdeA | - | 1 * | - | 1 * | - |
| norA + norB + norC + mepA + sepA | - | 2 * | 1 | 2 * | 2 * |
| MDR-MSSA (n = 15) | |||||
| Resistant isolates (n) | 8 | 11 | 9 | 11 | 13 |
| norA + norB + norC + mepA + sepA + mdeA | 5 | 8 | 5 | 7 | 8 |
| norA + norB + norC + mepA + sepA + mdeA + qacA/B | 1 | - | 1 * | 1 * | 1 * |
| norA + norB + norC + mepA + sepA | - | 2 * | 1 | 2 * | 2 * |
| Countries | Efflux Pump Genes (%) | References | |||||||
|---|---|---|---|---|---|---|---|---|---|
| norA | norB | norC | mepA | sepA | mdeA | qacA/B | smr | ||
| China | 96.2 | 98.2 | 92.5 | 90.6 | 96.2 | 94.3 | 83.0 | 77.4 | [28] |
| Iraq | 80.2 | 56.2 | 17.8 | 92.7 | - | 100 | - | - | [29] |
| South Africa | 98.9 | 98.9 | 79.4 | 97.9 | 96.9 | 95.9 | - | - | [30] |
| Iran | 41.7 | 41.7 | 41.7 | 60.0 | 35.0 | 61.7 | 3.3 | 30.0 | [12] |
| Portugal | - | - | - | 100 | 100 | - | 22.4 | 1.0 | [31] |
| Malaysia | - | - | - | - | - | - | 83.3 | 1.6 | [32] |
| Canada | - | - | - | - | - | - | 2.0 | 7.0 | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suma, T.A.; Alam, N.; Raihan, S.Z.; Zahid, M.A.; Mandal, S.C.; Suchana, F.J.; Kundu, R.; Hossain, A.; Muhit, M.A. Association of Antibacterial Susceptibility Profile with the Prevalence of Genes Encoding Efflux Proteins in the Bangladeshi Clinical Isolates of Staphylococcus aureus. Antibiotics 2023, 12, 305. https://doi.org/10.3390/antibiotics12020305
Suma TA, Alam N, Raihan SZ, Zahid MA, Mandal SC, Suchana FJ, Kundu R, Hossain A, Muhit MA. Association of Antibacterial Susceptibility Profile with the Prevalence of Genes Encoding Efflux Proteins in the Bangladeshi Clinical Isolates of Staphylococcus aureus. Antibiotics. 2023; 12(2):305. https://doi.org/10.3390/antibiotics12020305
Chicago/Turabian StyleSuma, Tanjina Akter, Nushrat Alam, Sheikh Zahir Raihan, Md. Al Zahid, Shankar Chandra Mandal, Fahrin Jahan Suchana, Ripa Kundu, Anwar Hossain, and Md. Abdul Muhit. 2023. "Association of Antibacterial Susceptibility Profile with the Prevalence of Genes Encoding Efflux Proteins in the Bangladeshi Clinical Isolates of Staphylococcus aureus" Antibiotics 12, no. 2: 305. https://doi.org/10.3390/antibiotics12020305