
Giving a Hand: Synthetic Peptides Boost the Antifungal Activity of Itraconazole against Cryptococcus neoformans
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material and Chemicals
2.2. Synthetic Peptides
2.3. Evaluation of Combined Antifungal Activity between SAMPs and ITR
2.4. Cell Membrane Integrity Assay
2.5. Overproduction of Reactive Oxygen Species (ROS)
2.6. 3/7 Caspase Activity
2.7. Counting Fluorescent Cells Using ImageJ
2.8. Statistical Analysis
3. Results
3.1. Combined Anticryptococcal Activity of Synthetic Peptides Combined with ITR
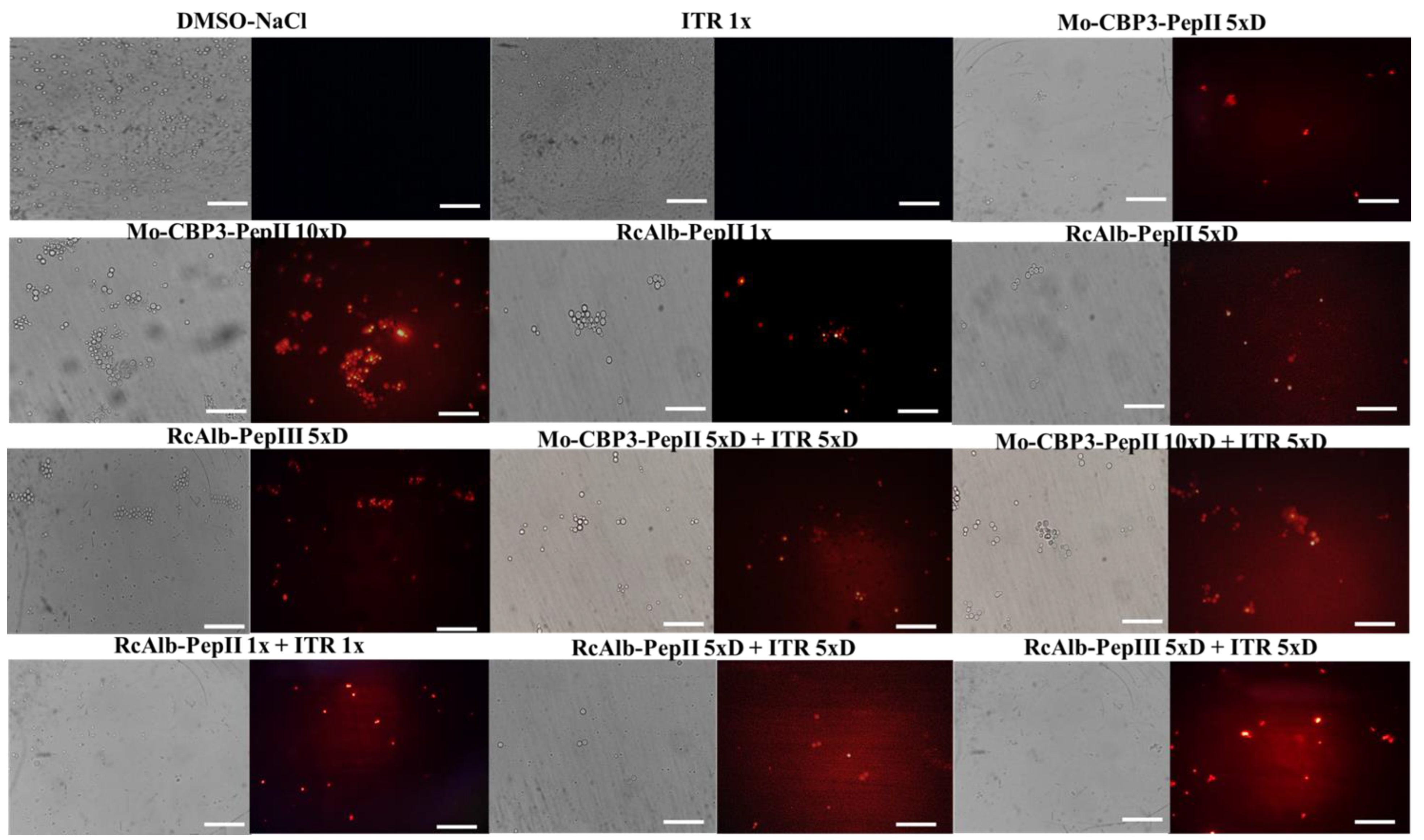
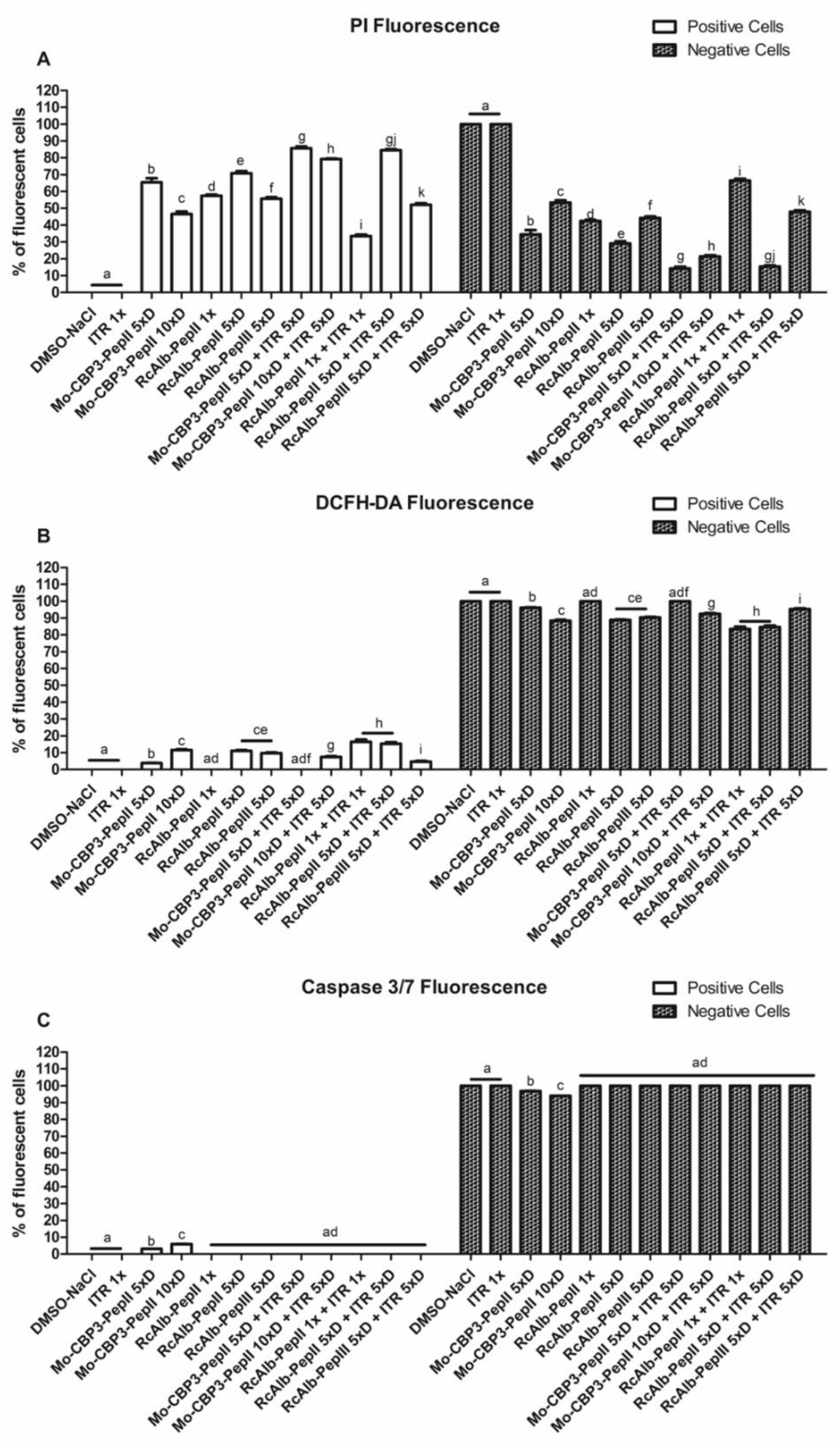
3.2. Membrane Pore Formation on C. neoformans Cells
3.3. ROS Overaccumulation and Apoptosis in C. neoformans Cells
3.4. Counting C. neoformans Fluorescent Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bermas, A.; Geddes-McAlister, J. Combatting the Evolution of Antifungal Resistance in Cryptococcus Neoformans. Mol. Microbiol. 2020, 114, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Robbins, N.; Caplan, T.; Cowen, L.E. Molecular Evolution of Antifungal Drug Resistance. Annu. Rev. Microbiol. 2017, 71, 753–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the Emerging Threat of Antifungal Resistance to Human Health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Zafar, H.; Altamirano, S.; Ballou, E.R.; Nielsen, K. A Titanic Drug Resistance Threat in Cryptococcus Neoformans. Curr. Opin. Microbiol. 2019, 52, 158–164. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic Antimicrobial Peptides: From Choice of the Best Sequences to Action Mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef]
- Mahindra, A.; Bagra, N.; Wangoo, N.; Khan, S.I.; Jacob, M.R.; Jain, R. Discovery of Short Peptides Exhibiting High Potency against Cryptococcus neoformans. ACS Med. Chem. Lett. 2014, 5, 315–320. [Google Scholar] [CrossRef]
- Aguiar, T.K.B.; Neto, N.A.S.; Freitas, C.D.T.; Silva, A.F.B.; Bezerra, L.P.; Malveira, E.A.; Branco, L.A.C.; Mesquita, F.P.; Goldman, G.H.; Alencar, L.M.R.; et al. Antifungal Potential of Synthetic Peptides against Cryptococcus Neoformans: Mechanism of Action Studies Reveal Synthetic Peptides Induce Membrane–Pore Formation, DNA Degradation, and Apoptosis. Pharmaceutics 2022, 14, 1678. [Google Scholar] [CrossRef]
- Bezerra, L.P.; Freitas, C.D.T.; Silva, A.F.B.; Amaral, J.L.; Neto, N.A.S.; Silva, R.G.G.; Parra, A.L.C.; Goldman, G.H.; Oliveira, J.T.A.; Mesquita, F.P.; et al. Synergistic Antifungal Activity of Synthetic Peptides and Antifungal Drugs against Candida Albicans and C. Parapsilosis Biofilms. Antibiotics 2022, 11, 553. [Google Scholar] [CrossRef]
- Bezerra, L.P.; Silva, A.F.; Santos-Oliveira, R.; Alencar, L.M.; Amaral, J.L.; Neto, N.A.; Silva, R.G.; Belém, M.O.; de Andrade, C.R.; Oliveira, J.T.; et al. Combined Antibiofilm Activity of Synthetic Peptides and Antifungal Drugs Against Candida spp. Future Microbiol. 2022, 17, 1133–1146. [Google Scholar] [CrossRef]
- Lima, P.G.; Souza, P.F.N.; Freitas, C.D.T.; Oliveira, J.T.A.; Dias, L.P.; Neto, J.X.S.; Vasconcelos, I.M.; Lopes, J.L.S.; Sousa, D.O.B. Anticandidal Activity of Synthetic Peptides: Mechanism of Action Revealed by Scanning Electron and Fluorescence Microscopies and Synergism Effect with Nystatin. J. Pept. Sci. 2020, 26, e3249. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Lima, P.G.; Freitas, C.D.T.; Sousa, D.O.B.; Neto, N.A.S.; Dias, L.P.; Vasconcelos, I.M.; Freitas, L.B.N.; Silva, R.G.G.; Sousa, J.S.; et al. Antidermatophytic Activity of Synthetic Peptides: Action Mechanisms and Clinical Application as Adjuvants to Enhance the Activity and Decrease the Toxicity of Griseofulvin. Mycoses 2020, 63, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.P.; Souza, P.F.N.; Oliveira, J.T.A.; Vasconcelos, I.M.; Araújo, N.M.S.; Tilburg, M.F.V.; Guedes, M.I.F.; Carneiro, R.F.; Lopes, J.L.S.; Sousa, D.O.B. RcAlb-PepII, a Synthetic Small Peptide Bioinspired in the 2S Albumin from the Seed Cake of Ricinus Communis, Is a Potent Antimicrobial Agent against Klebsiella Pneumoniae and Candida Parapsilosis. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183092. [Google Scholar] [CrossRef] [PubMed]
- Qorri, B.; Harless, W.; Szewczuk, M.R. Novel Molecular Mechanism of Aspirin and Celecoxib Targeting Mammalian Neuraminidase-1 Impedes Epidermal Growth Factor Receptor Signaling Axis and Induces Apoptosis in Pancreatic Cancer Cells. Drug Des. Devel. Ther. 2020, 14, 4149–4167. [Google Scholar] [CrossRef] [PubMed]
- Iyer, K.R.; Revie, N.M.; Fu, C.; Robbins, N.; Cowen, L.E. Treatment Strategies for Cryptococcal Infection: Challenges, Advances and Future Outlook. Nat. Rev. Microbiol. 2021, 19, 454–466. [Google Scholar] [CrossRef]
- Teaford, H.R.; Saleh, O.M.A.; Villarraga, H.R.; Enzler, M.J.; Rivera, C.G. The Many Faces of Itraconazole Cardiac Toxicity. Mayo Clin. Proc. Innov. Qual. Outcomes 2020, 4, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.K.; Grilley, M.; Anderson, T.; Dhiman, C.; Oblad, J.; Chang, C.W.T.; Sorensen, K.N.; Takemoto, J.Y. In Vitro Antifungal Synergy between Amphiphilic Aminoglycoside K20 and Azoles against Candida Species and Cryptococcus Neoformans. Med. Mycol. 2015, 53, 837–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Rocha, L.F.; Pippi, B.; Fuentefria, A.M.; Mezzari, A. Synergistic Effect of Ibuprofen with Itraconazole and Fluconazole against Cryptococcus neoformans. Braz. J. Pharm. Sci. 2020, 56, 1–6. [Google Scholar] [CrossRef]
- Saag, M.S.; Graybill, R.J.; Larsen, R.A.; Pappas, P.G.; Perfect, J.R.; Powderly, W.G.; Sobel, J.D.; Dismukes, W.E. Practice Guidelines for the Management of Cryptococcal Disease. Clin. Infect. Dis. 2000, 30, 710–718. [Google Scholar] [CrossRef] [Green Version]
- Denning, D.W.; Tucker, R.M.; Hanson, L.H.; Hamilton, J.R.; Stevens, D.A. Itraconazole Therapy for Cryptococcal Meningitis and Cryptococcosis. Arch. Intern. Med. 1989, 149, 2301–2308. [Google Scholar] [CrossRef]
- Cauwenbergh, G. Cryptococcal Meningitis: The Place of Itraconazole. Mycoses 1993, 36, 221–228. [Google Scholar] [CrossRef]
- Tenforde, M.W.; Shapiro, A.E.; Rouse, B.; Jarvis, J.N.; Li, T.; Eshun-Wilson, I.; Ford, N. Treatment for HIV-Associated Cryptococcal Meningitis. Cochrane Database Syst. Rev. 2018, 2018, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.-H.; et al. Clinical Practice Guidelines for the Management of Cryptococcal Disease: 2010 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, M.R.; Coote, P.J. Combination of Caspofungin or Anidulafungin with Antimicrobial Peptides Results in Potent Synergistic Killing of Candida Albicans and Candida Glabrata in Vitro. Int. J. Antimicrob. Agents 2010, 35, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Maurya, I.K.; Pathak, S.; Sharma, M.; Sanwal, H.; Chaudhary, P.; Tupe, S.; Deshpande, M.; Chauhan, V.S.; Prasad, R. Antifungal Activity of Novel Synthetic Peptides by Accumulation of Reactive Oxygen Species (ROS) and Disruption of Cell Wall against Candida Albicans. Peptides 2011, 32, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Queiroz-Telles, F.; Silva, N.; Carvalho, M.M.; Alcântara, A.P.; da Matta, D.; Barberino, M.G.; Bartczak, S.; Colombo, A.L. Evaluation of Efficacy and Safety of Itraconazole Oral Solution for the Treatment of Oropharyngeal Candidiasis in Aids Patients. Braz. J. Infect. Dis. 2001, 5, 60–66. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments | % of Inhibition of C. neoformans Growth |
|---|---|
| DMSO-NaCl | 0 |
| ITR 1× | 45.3 ± 0.021 |
| ITR 5×D | 12.2 ± 0.003 |
| ITR 10×D | 0 |
| Mo-CBP3-PepII 1× (MIC50) | 50.0 ± 0.004 |
| Mo-CBP3-PepII 5×D | 12.4 ± 0.001 |
| Mo-CBP3-PepII 10×D | 0 |
| RcAlb-PepII 1× (MIC50) | 50.0 ± 0.001 |
| RcAlb-PepII 5×D | 2.6 ± 0.005 |
| RcAlb-PepII 10×D | 0 |
| RcAlb-PepIII 1× (MIC50) | 50.0 ± 0.004 |
| RcAlb-PepIII 5×D | 10.6 ± 0.003 |
| RcAlb-PepIII 10×D | 0 |
| PepGAT 1× (MIC50) | 50.0 ± 0.009 |
| PepGAT 5×D | 17.1 ± 0.003 |
| PepGAT 10×D | 0 |
| PepGAT 1× (MIC50) | 50.0 ± 0.005 |
| PepGAT 5×D | 20.4 ± 0.009 |
| PepGAT 10×D | 0 |
| Mo-CBP3-PepII 1× + ITR 1× | 78.8 ± 0.004 |
| Mo-CBP3-PepII 5×D + ITR 1× | 74.5 ± 0.008 |
| Mo-CBP3-PepII 10×D + ITR 1× | 73.8 ± 0.009 |
| Mo-CBP3-PepII 5×D + ITR 5×D | 84.1 ± 0.001 |
| Mo-CBP3-PepII 10×D + ITR 5×D | 87.2 ± 0.002 |
| Mo-CBP3-PepII 5×D + ITR 10×D | 71.7 ± 0.005 |
| Mo-CBP3-PepII 10×D + ITR 10×D | 70.8 ± 0.006 |
| Mo-CBP3-PepII 1× + ITR 1× | 78.8 ± 0.002 |
| RcAlb-PepII 1× + ITR 1× | 83.9 ± 0.001 |
| RcAlb-PepII 5×D + ITR 1× | 74.8 ± 0.006 |
| RcAlb-PepII 10×D + ITR 1× | 65.6 ± 0.007 |
| RcAlb-PepII 5×D + ITR 5×D | 82.3 ± 0.002 |
| RcAlb-PepII10×D + ITR 5×D | 63.5 ± 0.005 |
| RcAlb-PepII 5×D + ITR 10×D | 71.7 ± 0.004 |
| RcAlb-PepII 10×D + ITR 10×D | 70.8 ± 0.003 |
| RcAlb-PepIII 1× + ITR 1× | 73.8 ± 0.004 |
| RcAlb-PepIII 5×D + ITR 1× | 74.5 ± 0.001 |
| RcAlb-PepIII 10×D + ITR 1× | 69.8 ± 0.01 |
| RcAlb-PepIII 5×D + ITR 5×D | 84.1 ± 0.02 |
| RcAlb-PepIII 10×D + ITR 5×D | 73.9 ± 0.009 |
| RcAlb-PepIII 5×D + ITR 10×D | 66.3 ± 0.001 |
| RcAlb-PepIII 10×D + ITR 10×D | 49.0 ± 0.008 |
| PepGAT 1× + ITR 1× | 73.4 ± 0.02 |
| PepGAT 5×D + ITR 1× | 69.4 ± 0.005 |
| PepGAT 10×D + ITR 1× | 66.3 ± 0.011 |
| PepGAT 5×D + ITR 5×D | 59.0 ± 0.02 |
| PepGAT 10×D + ITR 5×D | 54.2 ± 0.001 |
| PepGAT 5×D + ITR 10×D | 52.8 ± 0.008 |
| PepGAT 10×D + ITR 10×D | 47.4 ± 0.005 |
| PepKAA 1× + ITR 1× | 68.6 ± 0.003 |
| PepKAA 5×D + ITR 1× | 71.4 ± 0.004 |
| PepKAA 10×D + ITR 1× | 69.6 ± 0.05 |
| PepKAA 5×D + ITR 5×D | 45.6 ± 0.002 |
| PepKAA 10×D + ITR 5×D | 50.3 ± 0.015 |
| PepKAA 5×D + ITR 10×D | 50.3 ± 0.025 |
| PepKAA 10×D + ITR 10×D | 50.6 ± 0.009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguiar, T.K.B.; Feitosa, R.M.; Neto, N.A.S.; Malveira, E.A.; Gomes, F.I.R.; Costa, A.C.M.; Freitas, C.D.T.; Mesquita, F.P.; Souza, P.F.N. Giving a Hand: Synthetic Peptides Boost the Antifungal Activity of Itraconazole against Cryptococcus neoformans. Antibiotics 2023, 12, 256. https://doi.org/10.3390/antibiotics12020256
Aguiar TKB, Feitosa RM, Neto NAS, Malveira EA, Gomes FIR, Costa ACM, Freitas CDT, Mesquita FP, Souza PFN. Giving a Hand: Synthetic Peptides Boost the Antifungal Activity of Itraconazole against Cryptococcus neoformans. Antibiotics. 2023; 12(2):256. https://doi.org/10.3390/antibiotics12020256
Chicago/Turabian StyleAguiar, Tawanny K. B., Ricardo M. Feitosa, Nilton A. S. Neto, Ellen A. Malveira, Francisco I. R. Gomes, Ana C. M. Costa, Cleverson D. T. Freitas, Felipe P. Mesquita, and Pedro F. N. Souza. 2023. "Giving a Hand: Synthetic Peptides Boost the Antifungal Activity of Itraconazole against Cryptococcus neoformans" Antibiotics 12, no. 2: 256. https://doi.org/10.3390/antibiotics12020256