
Klebsiella pneumonia in Sudan: Multidrug Resistance, Polyclonal Dissemination, and Virulence

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population Genomics of Klebsiella Pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella Pneumoniae: A Major Worldwide Source and Shuttle for Antibiotic Resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Silvester, R.; Madhavan, A.; Kokkat, A.; Parolla, A.; Adarsh, B.M.; Harikrishnan, M.; Abdulla, M.H. Global Surveillance of Antimicrobial Resistance and Hypervirulence in Klebsiella Pneumoniae from LMICs: An in-Silico Approach. Sci. Total Environ. 2022, 802, 149859. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- WHO. Geneva Global Action Plan on Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2017; pp. 1–28. [Google Scholar]
- WHO. Country Cooperation Strategy for WHO and Sudan; World Health Organization: Geneva, Switzerland, 2022; ISBN 9789290229698. [Google Scholar]
- WHO. Global Priority List of Antibiotic-Resistant Batceria to Guide Research, Discovery, and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017; p. 7. [Google Scholar]
- Alsammani, M.A.; Ahmed, M.I.; Abdelatif, N.F. Bacterial Uropathogens Isolates and Antibiograms in Children under 5 Years of Age. Med. Arch. 2014, 68, 239–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albasha, A.M.; Osman, E.H.; Abd-Alhalim, S.; Alshaib, E.F.; Al-Hassan, L.; Altayb, H.N. Detection of Several Carbapenems Resistant and Virulence Genes in Classical and Hyper-Virulent Strains of Klebsiella Pneumoniae Isolated from Hospitalized Neonates and Adults in Khartoum. BMC Res. Notes 2020, 13, 312. [Google Scholar] [CrossRef]
- Elbadawi, H.S.; Elhag, K.M.; Mahgoub, E.; Altayb, H.N.; Abdel Hamid, M.M. Antimicrobial Resistance Surveillance among Gram-Negative Bacterial Isolates from Patients in Hospitals in Khartoum State, Sudan [Version 1; Peer Review: Awaiting Peer Review]. F1000Research 2019, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Hamdan, H.Z.; Kubbara, E.; Adam, A.M.; Hassan, O.S.; Suliman, S.O.; Adam, I. Urinary Tract Infections and Antimicrobial Sensitivity among Diabetic Patients at Khartoum, Sudan. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 26. [Google Scholar] [CrossRef] [Green Version]
- Abd Alfadil, N.A.; Suliman Mohamed, M.; Ali, M.M.; El Nima, E.A.I. Characterization of Pathogenic Bacteria Isolated from Sudanese Banknotes and Determination of Their Resistance Profile. Int. J. Microbiol. 2018, 2018, 4375164. [Google Scholar] [CrossRef]
- Osman, E.A.; El-Amin, N.E.; Al-Hassan, L.L.; Mukhtar, M. Multiclonal Spread of Klebsiella Pneumoniae across Hospitals in Khartoum, Sudan. J. Glob. Antimicrob. Resist. 2021, 24, 241–245. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Wong, J.L.C.; Sanchez-Garrido, J.; Kwong, H.S.; Low, W.W.; Morecchiato, F.; Giani, T.; Rossolini, G.M.; Brett, S.J.; Clements, A.; et al. Widespread Emergence of OmpK36 Loop 3 Insertions among Multidrug-Resistant Clones of Klebsiella Pneumoniae. PLoS Pathog. 2022, 18, e1010334. [Google Scholar] [CrossRef] [PubMed]
- Wassef, M.; Abdelhaleim, M.; AbdulRahman, E.; Ghaith, D. The Role of OmpK35, OmpK36 Porins, and Production of β-Lactamases on Imipenem Susceptibility in Klebsiella Pneumoniae Clinical Isolates, Cairo, Egypt. Microb. Drug Resist. 2015, 21, 577–580. [Google Scholar] [CrossRef]
- Rodrigues, C.; Desai, S.; Passet, V.; Gajjar, D.; Brisse, S. Genomic Evolution of the Globally Disseminated Multidrug-Resistant Klebsiella Pneumoniae Clonal Group 147. Microb. Genomics 2022, 8, 000737. [Google Scholar] [CrossRef]
- Wong, J.L.C.; Romano, M.; Kerry, L.E.; Kwong, H.S.; Low, W.W.; Brett, S.J.; Clements, A.; Beis, K.; Frankel, G. OmpK36-Mediated Carbapenem Resistance Attenuates ST258 Klebsiella Pneumoniae in Vivo. Nat. Commun. 2019, 10, 3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papagiannitsis, C.C.; Loli, A.; Tzouvelekis, L.S.; Tzelepi, E.; Arlet, G.; Miriagou, V. SCO-1, a Novel Plasmid-Mediated Class A β-Lactamase with Carbenicillinase Characteristics from Escherichia Coli. Antimicrob. Agents Chemother. 2007, 51, 2185–2188. [Google Scholar] [CrossRef] [Green Version]
- Wyres, K.L.; Wick, R.R.; Judd, L.M.; Froumine, R.; Tokolyi, A.; Gorrie, C.L.; Lam, M.M.; Duchene, S.; Jenney, A.; Holt, E. Distinct Evolutionary Dynamics of Horizontal Gene Transfer in Drug Resistant and Virulent Clones of Klebsiella Pneumoniae. PLoS Genet. 2019, 15, e100811. [Google Scholar] [CrossRef] [Green Version]
- Turton, J.F.; Payne, Z.; Coward, A.; Hopkins, K.L.; Turton, J.A.; Doumith, M.; Woodford, N. Virulence Genes in Isolates of Klebsiella Pneumoniae from the UK during 2016, Including among Carbapenemase Gene-Positive Hypervirulent K1-St23 and ‘Non-Hypervirulent’ Types ST147, ST15 and ST383. J. Med. Microbiol. 2018, 67, 118–128. [Google Scholar] [CrossRef]
- Edward, E.A.; Mohamed, N.M.; Zakaria, A.S. Whole Genome Characterization of the High-Risk Clone ST383 Klebsiella Pneumoniae with a Simultaneous Carriage of BlaCTX-M-14 on IncLM Plasmid and BlaCTX-M-15 on Convergent IncHI1BIncFIB Plasmid from Egypt. Microorganisms 2022, 10, 1097. [Google Scholar] [CrossRef]
- Adam, M.A.; Elhag, W.I. Prevalence of Metallo-β-Lactamase Acquired Genes among Carbapenems Susceptible and Resistant Gram-Negative Clinical Isolates Using Multiplex PCR, Khartoum Hospitals, Khartoum Sudan. BMC Infect. Dis. 2018, 18, 4–9. [Google Scholar] [CrossRef]
- Al-Hassan, L.; Elbadawi, H.; Osman, E.; Ali, S.; Elhag, K.; Cantillon, D.; Wille, J.; Seifert, H.; Higgins, P.G. Molecular Epidemiology of Carbapenem-Resistant Acinetobacter Baumannii From Khartoum State, Sudan. Front. Microbiol. 2021, 12, 628736. [Google Scholar] [CrossRef]
- Afolayan, A.O.; Oaikhena, A.O.; Aboderin, A.O.; Olabisi, O.F.; Amupitan, A.A.; Abiri, O.V.; Ogunleye, V.O.; Odih, E.E.; Adeyemo, A.T.; Adeyemo, A.T.; et al. Clones and Clusters of Antimicrobial-Resistant Klebsiella from Southwestern Nigeria. Clin. Infect. Dis. 2021, 73, S308–S315. [Google Scholar] [CrossRef] [PubMed]
- Sherif, M.; Palmieri, M.; Mirande, C.; El-Mahallawy, H.; Rashed, H.G.; Abd-El-Reheem, F.; El-Manakhly, A.R.; Abdel-latif, R.A.R.; Aboulela, A.G.; Saeed, L.Y.; et al. Whole-Genome Sequencing of Egyptian Multidrug-Resistant Klebsiella Pneumoniae Isolates: A Multi-Center Pilot Study. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.B.; Kambal, S.; Munir, A.; Abdalla, N.; Hassan, M.; Hamad, A.; Mohammed, S.; Ahmed, F.; Hamid, O.; Ismail, A.; et al. First Whole-Genome Sequence of a Highly Resistant Klebsiella Pneumoniae Sequence Type 14 Strain Isolated from Sudan. Microbiol. Resour. Announc. 2019, 8, 14–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, E.A.; El-Amin, N.; Adrees, E.A.E.; Al-Hassan, L.; Mukhtar, M. Comparing Conventional, Biochemical and Genotypic Methods for Accurate Identification of Klebsiella Pneumoniae in Sudan. Access Microbiol. 2020, 2, e000096. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.G.; Prior, K.; Harmsen, D.; Seifert, H. Development and Evaluation of a Core Genome Multilocus Typing Scheme for Whole-Genome Sequence-Based Typing of Acinetobacter Baumannii. PLoS ONE 2017, 12, e0179228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, M.M.C.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.E. A Genomic Surveillance Framework and Genotyping Tool for Klebsiella pneumoniae and Its Related Species Complex. Nat. Commun. 2021, 12, 4188. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Performance Standards for Antimicrobial Susceptibility Testing An Informational Supplement for Global Application Developed through the Clinical and Laboratory Standards Institute, 26th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016; Volume M100S, ISBN 610.688.0700.
- EUCAST. EUCAST Reading Guide for Broth Microdilution; EUCAST: Växjö, Sweden, 2020. [Google Scholar]
- Aanensen, D.M.; Carlos, C.C.; Donado-Godoy, P.; Okeke, I.N.; Ravikumar, K.L.; Abudahab, K.; Abrudan, M.; Argimón, S.; Harste, H.; Kekre, M.; et al. Implementing Whole-Genome Sequencing for Ongoing Surveillance of Antimicrobial Resistance: Exemplifying Insights into Klebsiella Pneumoniae. Clin. Infect. Dis. 2021, 73, S255–S257. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
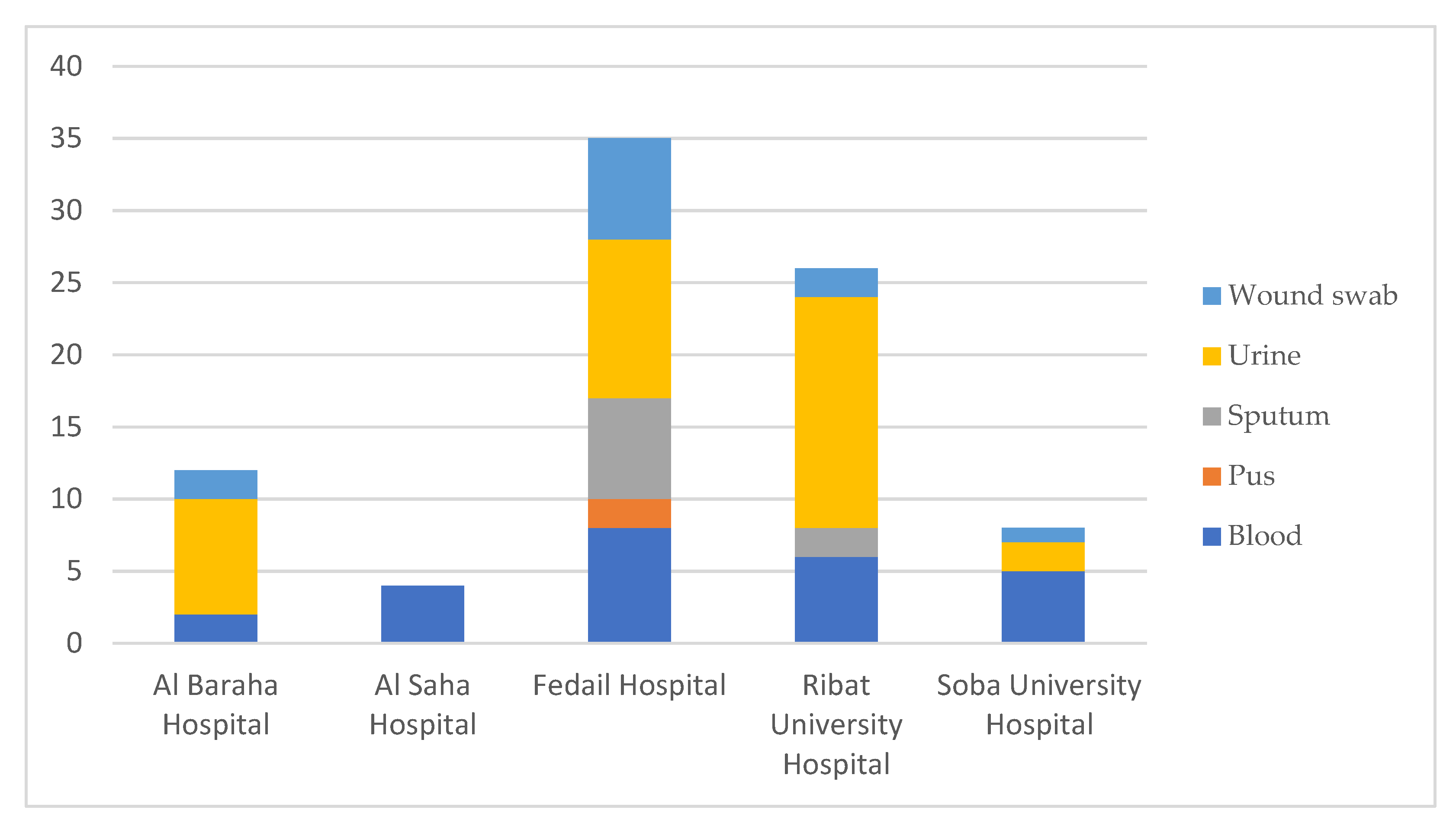
| Sample Name | Collection Date | Hospital | Isolation Source | Additional Information |
|---|---|---|---|---|
| HishK1 | 2016 | RUH | Urine | |
| HishK2 | 2016 | RUH | Blood | NICU |
| HishK10 | 2018 | SUH | Blood | NICU |
| HishK3 | 2018 | SUH | Urine | |
| HishK4 | 2018 | SUH | Urine | |
| HishK5 | 2018 | SUH | Blood | NICU |
| HishK7 | 2018 | SUH | Blood | NICU |
| HishK8 | 2018 | SUH | Blood | NICU |
| HishK9 | 2018 | SUH | Wound swab | |
| K10 | 2018 | ABH | Wound swab | |
| HishK11 | 2019 | ASH | Blood | |
| HishK12 | 2019 | ASH | Blood | |
| HishK13 | 2019 | ASH | Blood | |
| HishK14 | 2019 | ASH | Blood | |
| K11 | 2019 | ABH | Urine | |
| K12 | 2019 | ABH | Urine | |
| K13 | 2019 | ABH | Wound swab | |
| K15 | 2019 | ABH | Urine | |
| K2 | 2019 | ABH | Urine | |
| K3 | 2019 | ABH | Blood | NICU |
| K4 | 2019 | ABH | Urine | |
| K5 | 2019 | ABH | Urine | |
| K6 | 2019 | ABH | Blood | |
| K8 | 2019 | ABH | Urine | |
| K9 | 2019 | ABH | Urine | |
| LH_R146 | 08 February 2020 | RUH | Urine | |
| LH_S25 | 23 February 2020 | SUH | Blood | |
| LH_R100 | 26 February 2020 | RUH | Sputum | ICU |
| LH_R92 | 28 February 2020 | RUH | Urine | |
| LH_R107 | 01 March 2020 | RUH | Urine | NICU |
| LH_R120 | 03 March 2020 | RUH | Urine | |
| LH_R154 | 11 March 2020 | RUH | Urine | Outpatient |
| LH_R167 | 18 March 2020 | RUH | Blood | NICU |
| LH_R182 | 19 March 2020 | RUH | Blood | Inpatient |
| LH_R195 | 22 March 2020 | RUH | Blood | ICU |
| LH_R164 | 23 March 2020 | RUH | Sputum | Inpatient |
| LH_R174 | 23 March 2020 | RUH | Wound swab | Inpatient |
| LH_R162 | 23 March 2020 | RUH | Urine | Outpatient |
| LH_R208 | 26 March 2020 | RUH | Urine | Outpatient |
| LH_R223 | 28 March 2020 | RUH | Wound swab | Outpatient |
| LH_R219 | 28 March 2020 | RUH | Urine | Outpatient |
| LH_R275 | 12 July 2020 | RUH | Urine | Inpatient |
| LH_R289 | 18 July 2020 | RUH | Blood | Outpatient |
| LH_R290 | 27 July 2020 | RUH | Urine | Urology unit |
| LH_R314 | 30 July 2020 | RUH | Blood | |
| LH_R313 | 06 August 2020 | RUH | Urine | |
| LH_R323 | 08 August 2020 | RUH | Urine | |
| LH_R344 | 12 August 2020 | RUH | Urine | |
| LH_F2 | 16 August 2020 | FH | Urine | |
| LH_F15 | 16 August 2020 | FH | Sputum | |
| LH_F18 | 17 August 2020 | FH | Sputum | |
| LH_F25 | 17 August 2020 | FH | Blood | |
| LH_F50_1 | 18 August 2020 | FH | Urine | |
| LH_F35 | 18 August 2020 | FH | Urine | |
| LH_F68 | 20 August 2020 | FH | Wound swab | |
| LH_F64 | 20 August 2020 | FH | Wound swab | |
| LH_F66 | 20 August 2020 | FH | Urine | |
| LH_F82 | 21 August 2020 | FH | Pus | |
| LH_F86 | 21 August 2020 | FH | Urine | |
| LH_F97 | 22 August 2020 | FH | Urine | |
| LH_F101 | 22 August 2020 | FH | Urine | |
| LH_R384 | 22 August 2020 | RUH | Urine | |
| LH_F104 | 23 August 2020 | FH | Urine | |
| LH_F102 | 23 August 2020 | FH | Blood | |
| LH_F122 | 25 August 2020 | FH | Blood | |
| LH_F126 | 25 August 2020 | FH | Wound swab | |
| LH_F139 | 26 August 2020 | FH | Wound swab | |
| LH_F134 | 26 August 2020 | FH | Blood | |
| LH_F137 | 27 August 2020 | FH | Sputum | |
| LH_R387 | 27 August 2020 | RUH | Urine | |
| LH_F143 | 27 August 2020 | FH | Blood | |
| LH_F146 | 28 August 2020 | FH | Urine | |
| LH_F149 | 29 August 2020 | FH | Pus | |
| LH_F164 | 01 September 2020 | FH | Wound swab | |
| LH_F158 | 01 September 2020 | FH | Blood | |
| LH_F159 | 01 September 2020 | FH | Blood | |
| LH_F169 | 02 September 2020 | FH | Blood | |
| LH_F174 | 02 September 2020 | FH | Wound swab | |
| LH_F175 | 03 September 2020 | FH | Wound swab | |
| LH_F176 | 03 September 2020 | FH | Sputum | |
| LH_F190 | 05 September 2020 | FH | Sputum | |
| LH_F192 | 05 September 2020 | FH | Sputum | |
| LH_F281 | 20 September 2020 | FH | Urine |
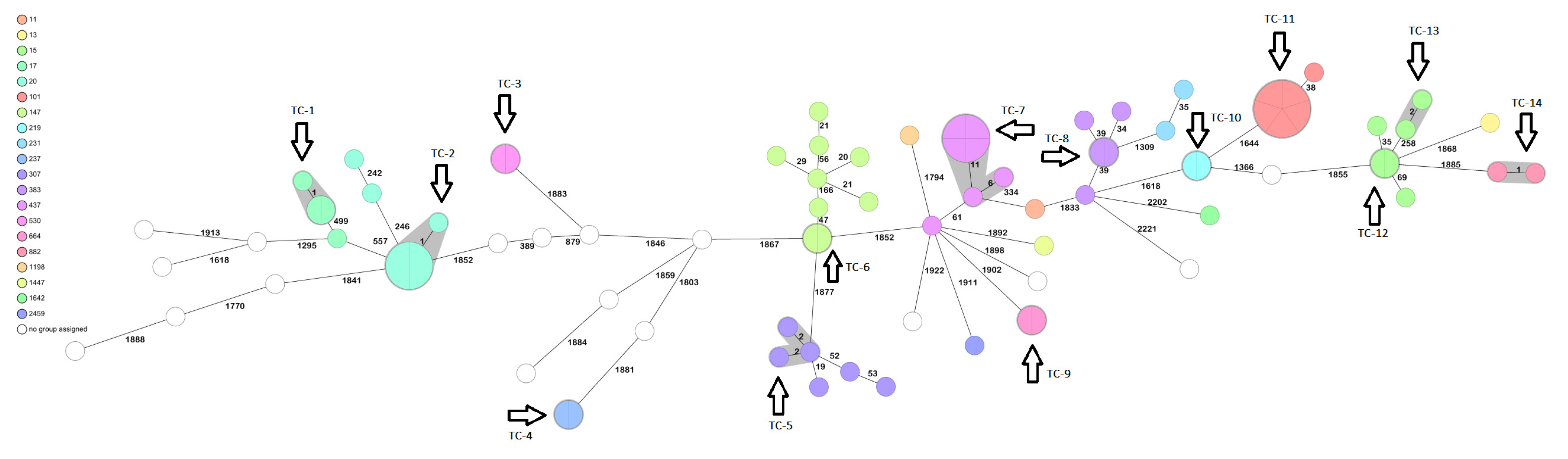
| ST | ESBL bla | Carb R bla | Intrinsic β-Lactamase bla | Virulence |
|---|---|---|---|---|
| 101 (6) | CTX-M-15 (6) | NDM-1 (1); ompK substitution (1) | SHV-1 | ybt9 (6) |
| 11 (1) | CTX-M-15 | NDM-4 | SHV-11 | ybt9 |
| 1198 (1) | - | - | SHV-11 | |
| 13 (1) | CTX-M-15 | - | SHV-1 | |
| 1147 (1) | - | - | SHV-27 | |
| 147 (9) | CTX-M-15 (9) | NDM-1 (5); NDM-5 (1) | SHV-11 | ybt9 (6) |
| ST15 (6) | CTX-M-15 (5) | OmpK35 substitution (1) | SHV-28 | ybt9 (1) |
| ST17 (4) | CTX-M-15 (2) | NDM-1 | SHV-11 | |
| ST20 (7) | CTX-M-15 (7) | NDM-1 (5) | SHV-187 | |
| ST218-3LV | - | - | SHV-93 | |
| ST219 (2) | CTX-M-15 (2) | - | SHV-1 | |
| ST231 (2) | CTX-M-15 (1) | OXA-232 (1) | SHV-1 | ybt14 (1); iuc (1) |
| ST237 (2) | - | - | SHV-11 | |
| ST24-1LV | - | - | SHV-11 | |
| ST2459 (1) | CTX-M-15 | - | SHV-1 | |
| ST2674 (1) | CTX-M-15 | NDM-1 | SHV-11 | |
| ST2735 (1) | - | - | SHV-11 | |
| ST29-1LV (1) | CTX-M-15 | - | - | |
| ST292 (1) | CTX-M-15 | - | SHV-11 | |
| ST307 (6) | CTX-M-15 (6) | NDM-1 (4) | SHV-28 | ybt10 (3) |
| ST3161 (1) | - | - | SHV-11 | |
| ST3430 (1) | CTX-M-15 | - | SHV-77 | |
| ST38 (1) | CTX-M-15 | NDM-1 | SHV-11 | |
| ST383 (5) | CTX-M-14 (4);CTX-M-15 (4) | NDM-5 (4); OXA-48 (4) | SHV-1 | iuc1 (4); rmp1 (2) |
| ST39 (1) | - | - | SHV-1 | ybt4 |
| ST437 (7) | CTX-M-15 (7) | NDM-1 (1); NDM-5 (6) | SHV-11 | ybt9 (7) |
| ST45 (1) | CTX-M-15 | OmpK36 variant | SHV-1 | ybt10 |
| ST469 (1) | CTX-M-15 | - | SHV-11 | |
| ST474 (1) | CTX-M-15 | - | SHV-11 | |
| ST501 (1) | CTX-M-15 | - | SHV-11 | |
| ST514 (1) | CTX-M-15 | - | SHV-63 | |
| ST530 (2) | CTX-M-15 (2) | NDM-1 (2) | - | ybt10 (2) |
| ST664 (2) | CTM-X-15 (2) | NDM-1 (2) | SHV-11 | |
| ST882 (2) | - | - | - | ybt (2) |
| ST901 (1) | CTX-M-15 | - | SHV-1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osman, E.A.; Yokoyama, M.; Altayb, H.N.; Cantillon, D.; Wille, J.; Seifert, H.; Higgins, P.G.; Al-Hassan, L. Klebsiella pneumonia in Sudan: Multidrug Resistance, Polyclonal Dissemination, and Virulence. Antibiotics 2023, 12, 233. https://doi.org/10.3390/antibiotics12020233
Osman EA, Yokoyama M, Altayb HN, Cantillon D, Wille J, Seifert H, Higgins PG, Al-Hassan L. Klebsiella pneumonia in Sudan: Multidrug Resistance, Polyclonal Dissemination, and Virulence. Antibiotics. 2023; 12(2):233. https://doi.org/10.3390/antibiotics12020233
Chicago/Turabian StyleOsman, Einas A., Maho Yokoyama, Hisham N. Altayb, Daire Cantillon, Julia Wille, Harald Seifert, Paul G. Higgins, and Leena Al-Hassan. 2023. "Klebsiella pneumonia in Sudan: Multidrug Resistance, Polyclonal Dissemination, and Virulence" Antibiotics 12, no. 2: 233. https://doi.org/10.3390/antibiotics12020233