
Staphylococcus aureus Adaptation to the Skin in Health and Persistent/Recurrent Infections
Abstract
:1. Introduction
2. S. aureus Metabolic Adaptation to the Skin
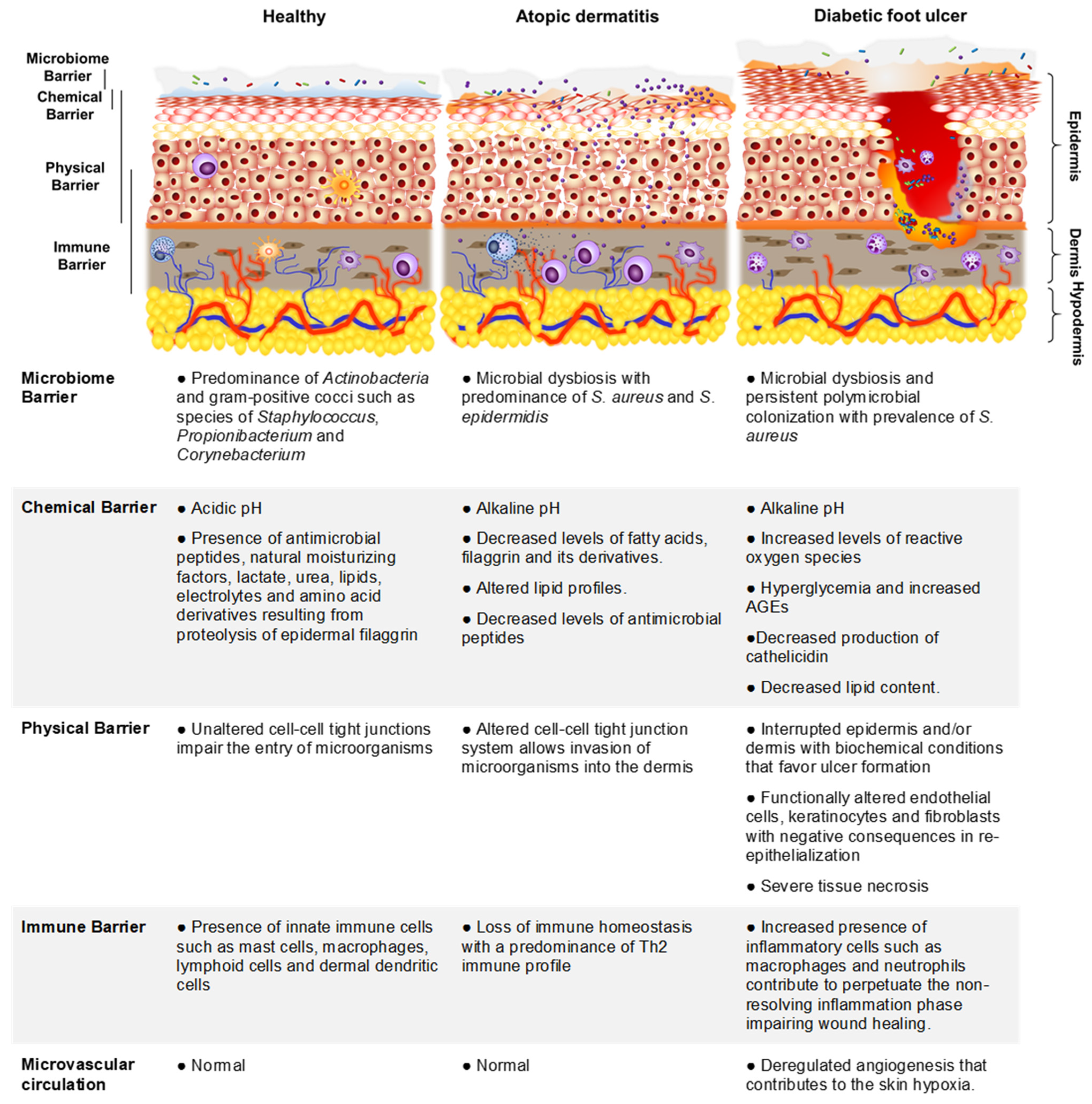
2.1. The Skin and Its Functions as a Cutaneous Barrier
2.2. S. aureus Colonization of Healthy Skin
2.3. S. aureus Colonization of the Skin in Individuals with Atopic Dermatitis
2.4. S. aureus Colonization of the Skin in Individuals with DFU
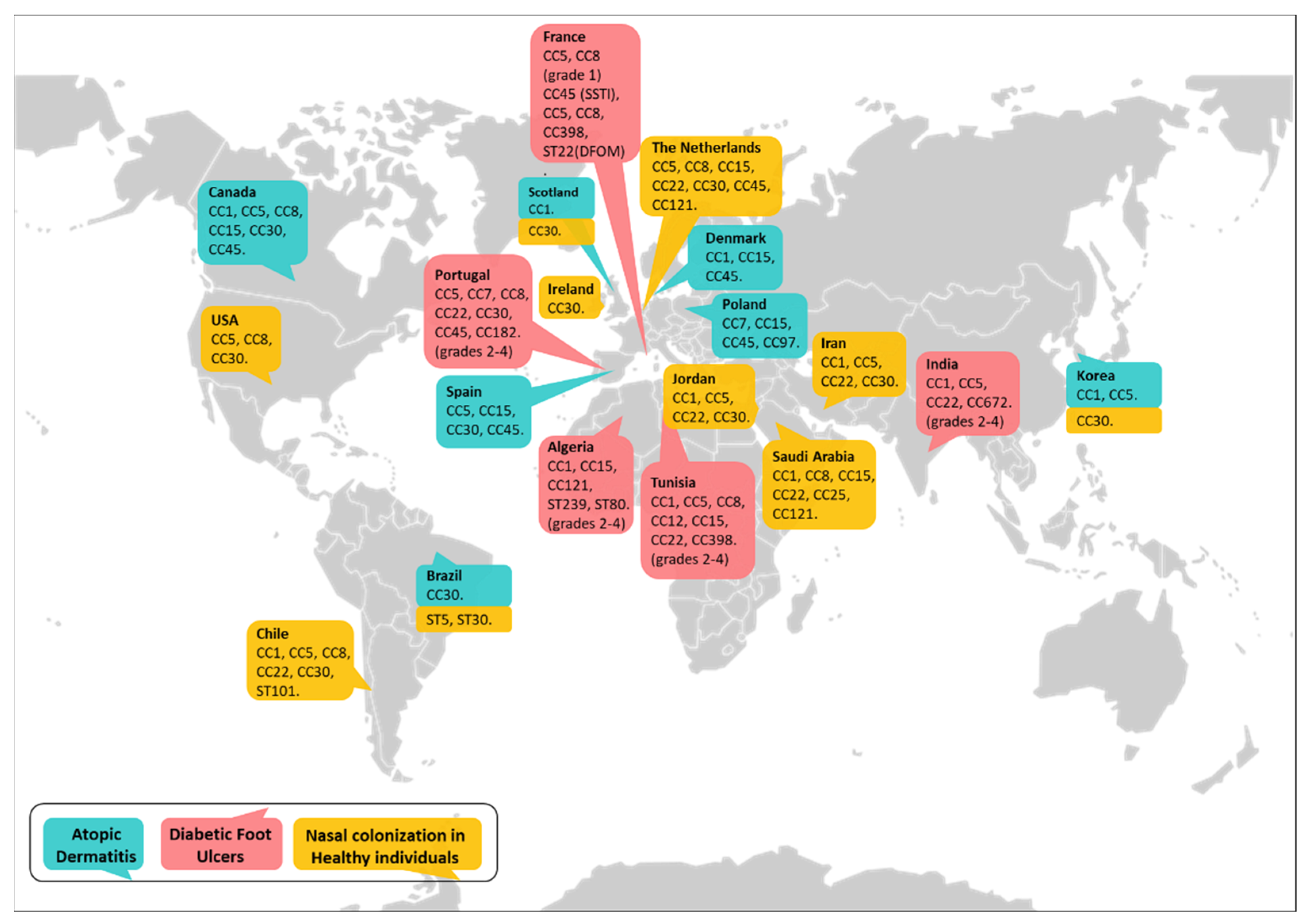
3. Genotypic Diversity among Nasal and Skin S. aureus Isolates in Health and Disease
3.1. S. aureus Nasal Carriage in Healthy Individuals
3.2. Lineages of S. aureus That Colonize and Infect Patients with Atopic Dermatitis
3.3. Major Clones of S. aureus That Colonize and Infect Skin Ulcers in Diabetic Patients
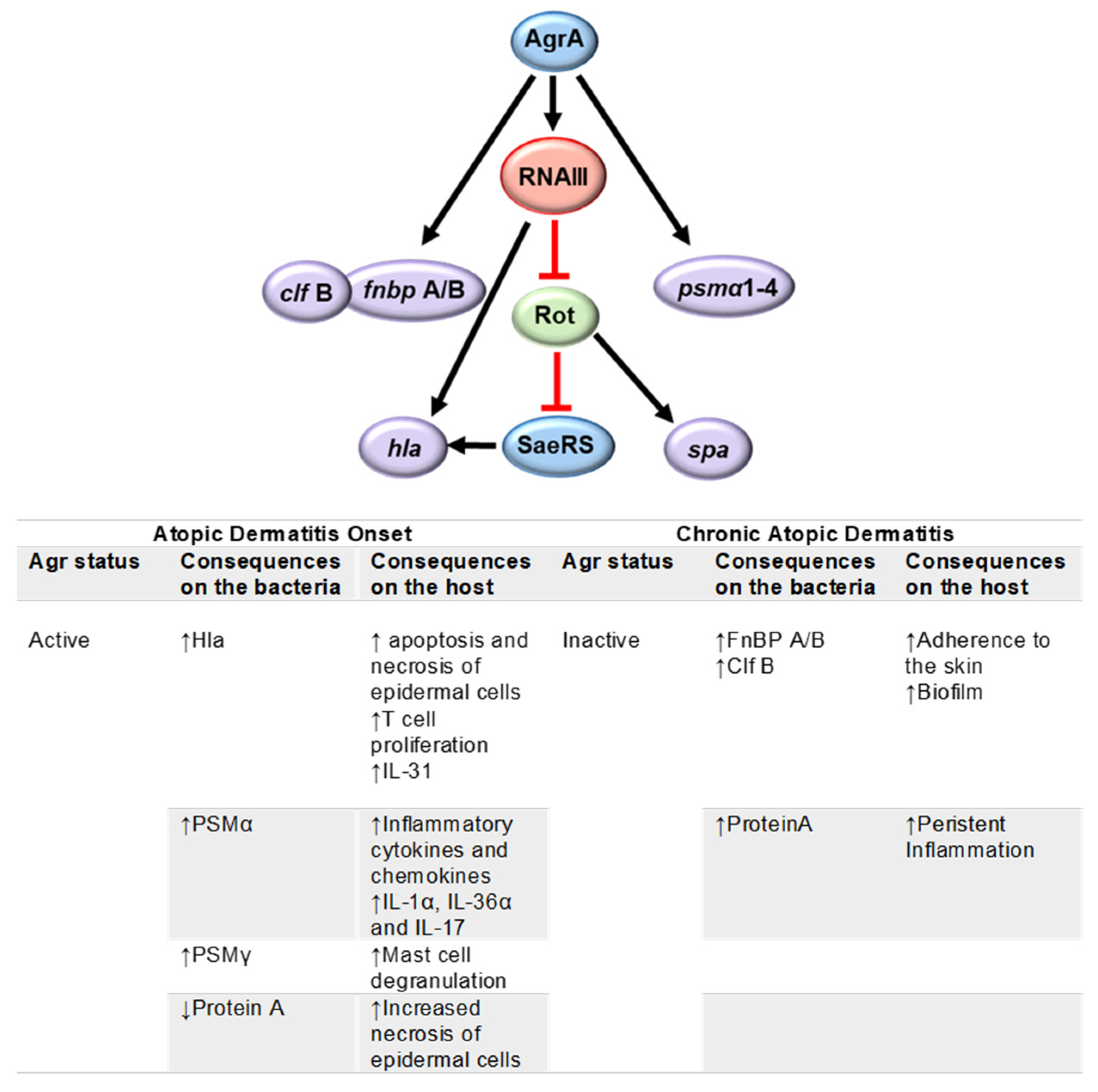
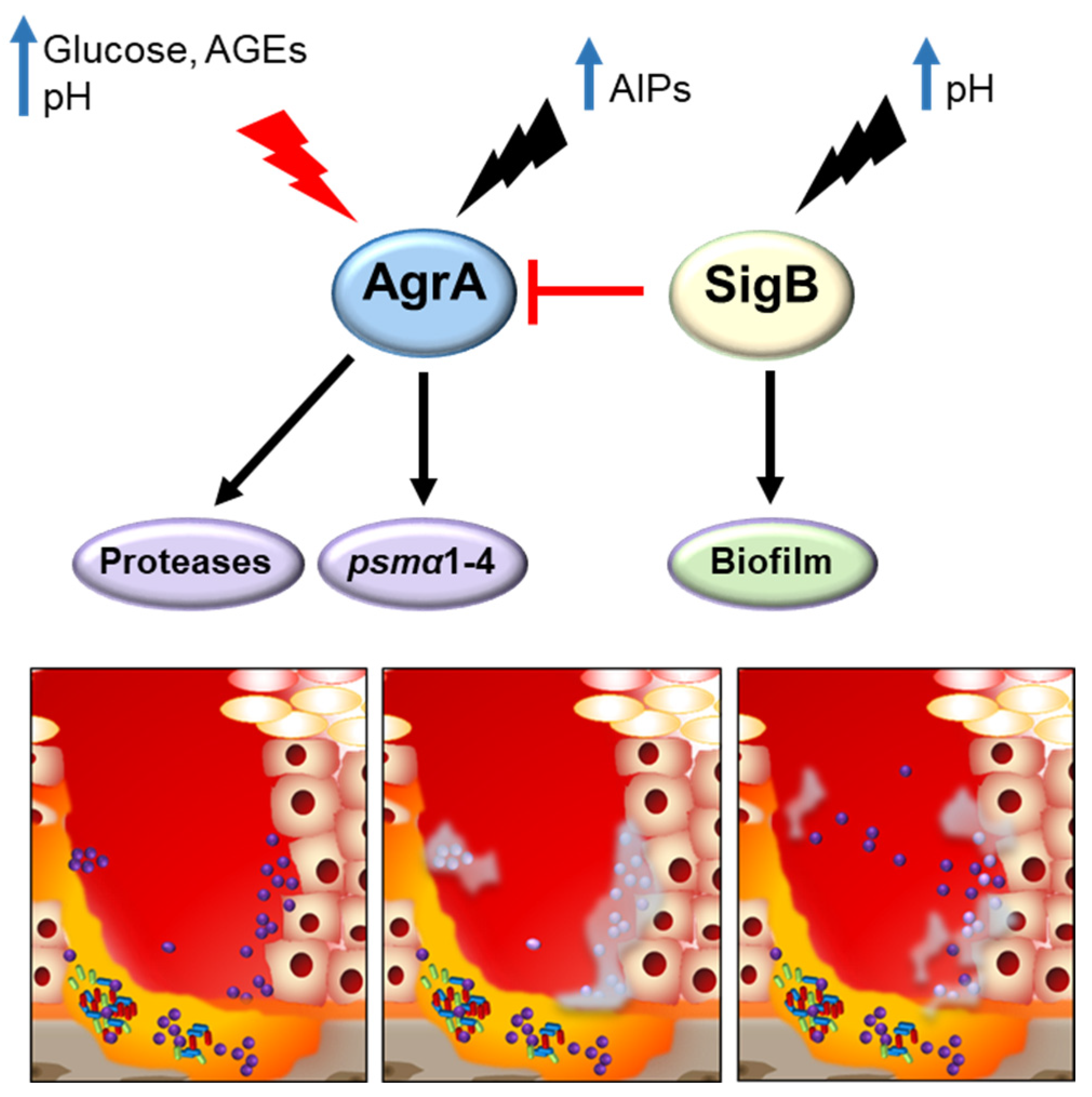
4. Regulation of Virulence Factor Expression and Its Impact on Disease Onset, Exacerbation and Chronicity
4.1. Atopic Dermatitis
4.2. Diabetic Foot Ulcer Infections
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Wertheim, H.F.L.; Melles, D.C.; Vos, M.C.; Van Leeuwen, W.; Van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The Role of Nasal Carriage in Staphylococcus aureus Infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- Bode, L.G.M.; Kluytmans, J.A.J.W.; Wertheim, H.F.L.; Bogaers, D.; Vandenbroucke-Grauls, C.M.J.E.; Roosendaal, R.; Troelstra, A.; Box, A.T.A.; Voss, A.; van der Tweel, I.; et al. Preventing Surgical-Site Infections in Nasal Carriers of Staphylococcus aureus. N. Engl. J. Med. 2010, 362, 9–17. [Google Scholar] [CrossRef]
- Nouwen, J.; Schouten, J.; Schneebergen, P.; Snijders, S.; Maaskant, J.; Koolen, M.; Van Belkum, A.; Verbrugh, H.A. Staphylococcus aureus Carriage Patterns and the Risk of Infections Associated with Continuous Peritoneal Dialysis. J. Clin. Microbiol. 2006, 44, 2233–2236. [Google Scholar] [CrossRef]
- Baker, J.L.; Bor, B.; Agnello, M.; Shi, W.; He, X. Ecology of the Oral Microbiome: Beyond Bacteria. Trends Microbiol. 2017, 25, 362–374. [Google Scholar] [CrossRef]
- Wertheim, H.F.L.; Vos, M.C.; Ott, A.; Van Belkum, A.; Voss, A.; Kluytmans, J.A.J.W.; Van Keulen, P.H.J.; Vandenbroucke-Grauls, C.M.J.E.; Meester, M.H.M.; Verbrugh, H.A. Risk and Outcome of Nosocomial Staphylococcus aureus Bacteraemia in Nasal Carriers versus Non-Carriers. Lancet 2004, 364, 703–705. [Google Scholar] [CrossRef]
- Eyerich, S.; Eyerich, K.; Traidl-Hoffmann, C.; Biedermann, T. Cutaneous Barriers and Skin Immunity: Differentiating A Connected Network. Trends Immunol. 2018, 39, 315–327. [Google Scholar] [CrossRef]
- Geoghegan, J.A.; Irvine, A.D.; Foster, T.J. Staphylococcus aureus and Atopic Dermatitis: A Complex and Evolving Relationship. Trends Microbiol. 2018, 26, 484–497. [Google Scholar] [CrossRef]
- Shettigar, K.; Murali, T.S. Virulence Factors and Clonal Diversity of Staphylococcus aureus in Colonization and Wound Infection with Emphasis on Diabetic Foot Infection. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2235–2246. [Google Scholar] [CrossRef]
- Krishna, S.; Miller, L.S. Host-Pathogen Interactions between the Skin and Staphylococcus aureus. Curr. Opin. Microbiol. 2012, 15, 28–35. [Google Scholar] [CrossRef]
- Ramsey, M.M.; Freire, M.O.; Gabrilska, R.A.; Rumbaugh, K.P.; Lemon, K.P. Staphylococcus aureus Shifts toward Commensalism in Response to Corynebacterium Species. Front. Microbiol. 2016, 7, 1230. [Google Scholar] [CrossRef]
- Gille, J.; Reisinger, K.; Westphal-Varghese, B.; Kaufmann, R. Decreased MRNA Stability as a Mechanism of Glucocorticoid-Mediated Inhibition of Vascular Endothelial Growth Factor Gene Expression by Cultured Keratinocytes. J. Investig. Dermatol. 2001, 117, 1581–1587. [Google Scholar] [CrossRef]
- McLean, W.H.I. Filaggrin Failure—From Ichthyosis Vulgaris to Atopic Eczema and Beyond. Br. J. Dermatol. 2016, 175, 4–7. [Google Scholar] [CrossRef]
- Brown, S.J.; McLean, W.H.I. One Remarkable Molecule: Filaggrin. J. Investig. Dermatol. 2012, 132, 751–762. [Google Scholar] [CrossRef]
- Lacey, K.; Geoghegan, J.; McLoughlin, R. The Role of Staphylococcus aureus Virulence Factors in Skin Infection and Their Potential as Vaccine Antigens. Pathogens 2016, 5, 22. [Google Scholar] [CrossRef]
- Kazuya, M.; Ohniwa, R.L.; Ohta, T.; Tanaka, Y.; Takeyasu, K.; Msadek, T. Adaptation beyond the Stress Response: Cell Structure Dynamics and Population Heterogeneity in Staphylococcus aureus. Microbes Environ. 2010, 25, 75–82. [Google Scholar] [CrossRef]
- Becker, K.; Bierbaum, G.; von Eiff, C.; Engelmann, S.; Götz, F.; Hacker, J.; Hecker, M.; Peters, G.; Rosenstein, R.; Ziebuhr, W. Understanding the Physiology and Adaptation of Staphylococci: A Post-Genomic Approach. Int. J. Med. Microbiol. 2007, 297, 483–501. [Google Scholar] [CrossRef]
- Proctor, R.A.; von Eiff, C.; Kahl, B.C.; Becker, K.; McNamara, P.; Herrmann, M.; Peters, G. Small Colony Variants: A Pathogenic Form of Bacteria That Facilitates Persistent and Recurrent Infections. Nat. Rev. Microbiol. 2006, 4, 295–305. [Google Scholar] [CrossRef]
- Deora, R.; Tseng, T.; Misra, T.K. Alternative Transcription Factor σ(SB) of Staphylococcus aureus: Characterization and Role in Transcription of the Global Regulatory Locus Sar. J. Bacteriol. 1997, 179, 6355–6359. [Google Scholar] [CrossRef]
- Inose, Y.; Takeshita, S.L.; Hidaka, T.; Higashide, M.; Maruyama, A.; Hayashi, H.; Morikawa, K.; Ohta, T. Genetic Characterization of the Natural SigB Variants Found in Clinical Isolates of Staphylococcus aureus. J. Gen. Appl. Microbiol. 2006, 52, 259–271. [Google Scholar] [CrossRef]
- Georgiou, G.; Segatori, L. Preparative Expression of Secreted Proteins in Bacteria: Status Report and Future Prospects. Curr. Opin. Biotechnol. 2005, 16, 538–545. [Google Scholar] [CrossRef]
- Costa, F.G.; Horswill, A.R. Overcoming PH Defenses on the Skin to Establish Infections. PLoS Pathog. 2022, 18, e1010512. [Google Scholar] [CrossRef]
- Rode, T.M.; Møretrø, T.; Langsrud, S.; Langsrud, Ø.; Vogt, G.; Holck, A. Responses of Staphylococcus aureus Exposed to HCL and Organic Acid Stress. Can. J. Microbiol. 2010, 56, 777–792. [Google Scholar] [CrossRef]
- Walker, K.A.; Miller, V.L. The Intersection of Capsule Gene Expression, Hypermucoviscosity and Hypervirulence in Klebsiella Pneumoniae. Curr. Opin. Microbiol. 2020, 54, 95–102. [Google Scholar] [CrossRef]
- Zhou, C.; Bhinderwala, F.; Lehman, M.K.; Thomas, V.C.; Chaudhari, S.S.; Yamada, K.J.; Foster, K.W.; Powers, R.; Kielian, T.; Fey, P.D. Urease Is an Essential Component of the Acid Response Network of Staphylococcus aureus and Is Required for a Persistent Murine Kidney Infection. PLoS Pathog. 2019, 15, e1007538. [Google Scholar] [CrossRef]
- Bäckhed, F.; Fraser, C.M.; Ringel, Y.; Sanders, M.E.; Sartor, R.B.; Sherman, P.M.; Versalovic, J.; Young, V.; Finlay, B.B. Defining a Healthy Human Gut Microbiome: Current Concepts, Future Directions, and Clinical Applications. Cell Host Microbe 2012, 12, 611–622. [Google Scholar] [CrossRef]
- Bore, E.; Langsrud, S.; Langsrud, Ø.; Rode, T.M.; Holck, A. Acid-Shock Responses in Staphylococcus aureus Investigated by Global Gene Expression Analysis. Microbiology 2007, 153, 2289–2303. [Google Scholar] [CrossRef]
- Maier, R.J.; Benoit, S.L. Role of Nickel in Microbial Pathogenesis. Inorganics 2019, 7, 80. [Google Scholar] [CrossRef]
- Joshi, G.S.; Spontak, J.S.; Klapper, D.G.; Richardson, A.R. Arginine Catabolic Mobile Element Encoded SpeG Abrogates the Unique Hypersensitivity of Staphylococcus aureuss to Exogenous Polyamines. Mol. Microbiol. 2011, 82, 9–20. [Google Scholar] [CrossRef]
- Li, L.; Li, Y.; Zhu, F.; Cheung, A.L.; Wang, G.; Bai, G.; Proctor, R.A.; Yeaman, M.R.; Bayer, A.S.; Xiong, Y.Q. New Mechanistic Insights into Purine Biosynthesis with Second Messenger C-Di-AMP in Relation to Biofilm-Related Persistent Methicillin-Resistant Staphylococcus aureus Infections. MBio 2021, 12, e02081-21. [Google Scholar] [CrossRef]
- Bieber, T. Atopic Dermatitis. Dermatol. Ther. 2008, 21, 96–100. [Google Scholar] [CrossRef]
- Weidinger, S.; Novak, N. Atopic Dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Boguniewicz, M.; Leung, D.Y.M. Atopic Dermatitis: A Disease of Altered Skin Barrier and Immune Dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef]
- Brandt, E.B.; Sivaprasad, U. Th2 Cytokines and Atopic Dermatitis Eric. J Clin. Cell Immunol. 2011, 2, 110–117. [Google Scholar] [CrossRef]
- Yeung, M.; Balma-Mena, A.; Shear, N.; Simor, A.; Pope, E.; Walsh, S.; McGavin, M.J. Identification of Major Clonal Complexes and Toxin Producing Strains among Staphylococcus aureus Associated with Atopic Dermatitis. Microbes Infect. 2011, 13, 189–197. [Google Scholar] [CrossRef]
- Kim, Y.; Lim, K.M. Skin Barrier Dysfunction and Filaggrin. Arch. Pharm. Res. 2021, 44, 36–48. [Google Scholar] [CrossRef]
- Kezic, S.; O’Regan, G.M.; Yau, N.; Sandilands, A.; Chen, H.; Campbell, L.E.; Kroboth, K.; Watson, R.; Rowland, M.; Irwin McLean, W.H.; et al. Levels of Filaggrin Degradation Products Are Influenced by Both Filaggrin Genotype and Atopic Dermatitis Severity. Allergy Eur. J. Allergy Clin. Immunol. 2011, 66, 934–940. [Google Scholar] [CrossRef]
- Howell, M.D.; Kim, B.E.; Gao, P.; Grant, A.V.; Boguniewicz, M.; DeBenedetto, A.; Schneider, L.; Beck, L.A.; Barnes, K.C.; Leung, D.Y.M. Cytokine Modulation of Atopic Dermatitis Filaggrin Skin Expression. J. Allergy Clin. Immunol. 2009, 124, 7–12. [Google Scholar] [CrossRef]
- Chieosilapatham, P.; Ogawa, H.; Niyonsaba, F. Current Insights into the Role of Human β-Defensins in Atopic Dermatitis. Clin. Exp. Immunol. 2017, 190, 155–166. [Google Scholar] [CrossRef]
- Cho, S.H.; Strickland, I.; Tomkinson, A.; Fehringer, A.P.; Gelfand, E.W.; Leung, D.Y.M. Preferential Binding of Staphylococcus aureus to Skin Sites of Th2-Mediated Inflammation in a Murine Model. J. Investig. Dermatol. 2001, 116, 658–663. [Google Scholar] [CrossRef]
- Jeong, C.W.; Ahnt, K.S.; Rho, N.K.; Park, Y.D.; Lee, D.Y.; Lee, J.H.; Lee, E.S.; Yang, J.M. Differential in Vivo Cytokine MRNA Expression in Lesional Skin of Intrinsic vs. Extrinsic Atopic Dermatitis Patients Using Semiquantitative RT-PCR. Clin. Exp. Allergy 2003, 33, 1717–1724. [Google Scholar] [CrossRef]
- Dillon, S.R.; Sprecher, C.; Hammond, A.; Bilsborough, J.; Rosenfeld-Franklin, M.; Presnell, S.R.; Haugen, H.S.; Maurer, M.; Harder, B.; Johnston, J.; et al. Interleukin 31, a Cytokine Produced by Activated T Cells, Induces Dermatitis in Mice. Nat. Immunol. 2004, 5, 752–760. [Google Scholar] [CrossRef]
- Sonkoly, E.; Muller, A.; Lauerma, A.I.; Pivarcsi, A.; Soto, H.; Kemeny, L.; Alenius, H.; Dieu-Nosjean, M.C.; Meller, S.; Rieker, J.; et al. IL-31: A New Link between T Cells and Pruritus in Atopic Skin Inflammation. J. Allergy Clin. Immunol. 2006, 117, 411–417. [Google Scholar] [CrossRef]
- Bonzano, L.; Borgia, F.; Casella, R.; Miniello, A.; Nettis, E.; Gangemi, S. Microbiota and IL-33/31 Axis Linkage: Implications and Therapeutic Perspectives in Atopic Dermatitis and Psoriasis. Biomolecules 2023, 13, 1100. [Google Scholar] [CrossRef]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; Mullikin, J.; et al. Temporal Shifts in the Skin Microbiome Associated with Disease Flares and Treatment in Children with Atopic Dermatitis. Genome Res. 2012, 22, 850–859. [Google Scholar] [CrossRef]
- Totté, J.E.E.; van der Feltz, W.T.; Hennekam, M.; van Belkum, A.; van Zuuren, E.J.; Pasmans, S.G.M.A. Prevalence and Odds of Staphylococcus aureus Carriage in Atopic Dermatitis: A Systematic Review and Meta-Analysis. Br. J. Dermatol. 2016, 175, 687–695. [Google Scholar] [CrossRef]
- Angulo, E.L.; Gern, J.E. Staphylococcus aureus and Staphylococcus epidermidis Strain Diversity Underlying Pediatric Atopic Dermatitis. Pediatrics 2018, 142, S229–S230. [Google Scholar] [CrossRef]
- Kim, J.; Kim, B.E.; Ahn, K.; Leung, D.Y.M. Interactions between Atopic Dermatitis and Staphylococcus aureus Infection: Clinical Implications. Allergy Asthma Immunol. Res. 2019, 11, 593–603. [Google Scholar] [CrossRef]
- Acker, K.P.; Wong Fok Lung, T.; West, E.; Craft, J.; Narechania, A.; Smith, H.; O’Brien, K.; Moustafa, A.M.; Lauren, C.; Planet, P.J.; et al. Strains of Staphylococcus aureus That Colonize and Infect Skin Harbor Mutations in Metabolic Genes. iScience 2019, 19, 281–290. [Google Scholar] [CrossRef]
- Kornberg, M.D.; Bhargava, P.; Kim, P.M.; Putluri, V.; Snowman, A.M.; Putluri, N.; Calabresi, P.A.; Snyder, S.H. Dimethyl Fumarate Targets GAPDH and Aerobic Glycolysis to Modulate Immunity. Science 2018, 360, 449–453. [Google Scholar] [CrossRef]
- Wickersham, M.; Wachtel, S.; Wong, T.; Lung, F.; Richardson, A.; Parker, D.; Wickersham, M.; Wachtel, S.; Wong, T.; Lung, F.; et al. Metabolic Stress Drives Keratinocyte Defenses against Staphylococcus aureus Infection Article Metabolic Stress Drives Keratinocyte Defenses against Staphylococcus aureus Infection. Cell Rep. 2017, 18, 2742–2751. [Google Scholar] [CrossRef]
- Cho, S.H.; Strickland, I.; Boguniewicz, M.; Leung, D.Y.M. Fibronectin and Fibrinogen Contribute to the Enhanced Binding of Staphylococcus aureus to Atopic Skin. J. Allergy Clin. Immunol. 2001, 108, 269–274. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chiang, H.I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The Microbiome Extends to Subepidermal Compartments of Normal Skin. Nat. Commun. 2013, 4, 1431. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Two, A.M.; Chun, K.A.; Narala, S.; Geha, R.S.; Hata, T.R.; Gallo, R.L. Staphylococcus aureus Exploits Epidermal Barrier Defects in Atopic Dermatitis to Trigger Cytokine Expression. J. Investig. Dermatol. 2016, 136, 2192–2200. [Google Scholar] [CrossRef]
- Elias, P.M.; Steinhoff, M. “Outside-to-inside” (and Now Back to “Outside”) Pathogenic Mechanisms in Atopic Dermatitis. J. Investig. Dermatol. 2008, 128, 1067–1070. [Google Scholar] [CrossRef]
- Jinnestål, C.L.; Belfrage, E.; Bäck, O.; Schmidtchen, A.; Sonesson, A. Skin Barrier Impairment Correlates with Cutaneous Staphylococcus aureus Colonization and Sensitization to Skin-Associated Microbial Antigens in Adult Patients with Atopic Dermatitis. Int. J. Dermatol. 2014, 53, 27–33. [Google Scholar] [CrossRef]
- Burgess, J.L.; Wyant, W.A.; Abujamra, B.A.; Kirsner, R.S.; Jozic, I. Diabetic Wound-Healing Science. Medicina 2021, 57, 1072. [Google Scholar] [CrossRef]
- Greener, B.; Hughes, A.A.; Bannister, N.P.; Douglass, J. Proteases and PH in Chronic Wounds. J. Wound Care 2005, 14, 59–61. [Google Scholar] [CrossRef]
- McArdle, C.D.; Lagan, K.M.; McDowell, D.A. Effects of PH on the Antibiotic Resistance of Bacteria Recovered from Diabetic Foot Ulcer Fluid: An in Vitro Study. J. Am. Podiatr. Med. Assoc. 2018, 108, 6–11. [Google Scholar] [CrossRef]
- Rivas-Santiago, B.; Trujillo, V.; Montoya, A.; Gonzalez-Curiel, I.; Castañeda-Delgado, J.; Cardenas, A.; Rincon, K.; Hernandez, M.L.; Hernández-Pando, R. Expression of Antimicrobial Peptides in Diabetic Foot Ulcer. J. Dermatol. Sci. 2012, 65, 19–26. [Google Scholar] [CrossRef]
- Dunyach-Remy, C.; Essebe, C.N.; Sotto, A.; Lavigne, J.P. Staphylococcus aureus Toxins and Diabetic Foot Ulcers: Role in Pathogenesis and Interest in Diagnosis. Toxins 2016, 8, 209. [Google Scholar] [CrossRef]
- Prompers, L.; Huijberts, M.; Schaper, N.; Apelqvist, J.; Bakker, K.; Edmonds, M.; Holstein, P.; Jude, E.; Jirkovska, A.; Mauricio, D.; et al. Resource Utilisation and Costs Associated with the Treatment of Diabetic Foot Ulcers. Prospective Data from the Eurodiale Study. Diabetologia 2008, 51, 1826–1834. [Google Scholar] [CrossRef] [PubMed]
- Giurato, L.; Meloni, M.; Izzo, V.; Uccioli, L. Osteomyelitis in Diabetic Foot: A Comprehensive Overview. World J. Diabetes 2017, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Thimmappaiah Jagadeesh, A.; Prakash, P.Y.; Karthik Rao, N.; Ramya, V. Culture Characterization of the Skin Microbiome in Type 2 Diabetes Mellitus: A Focus on the Role of Innate Immunity. Diabetes Res. Clin. Pract. 2017, 134, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Smit, J.; Søgaard, M.; Schønheyder, H.C.; Nielsen, H.; Frøslev, T.; Thomsen, R.W. Diabetes and Risk of Community-Acquired Staphylococcus aureus Bacteremia: A Population-Based Case-Control Study. Eur. J. Endocrinol. 2016, 174, 631–639. [Google Scholar] [CrossRef]
- Kalan, L.R.; Meisel, J.S.; Loesche, M.A.; Horwinski, J.; Soaita, I.; Chen, X.; Uberoi, A.; Gardner, S.E.; Grice, E.A. Strain- and Species-Level Variation in the Microbiome of Diabetic Wounds Is Associated with Clinical Outcomes and Therapeutic Efficacy. Cell Host Microbe 2019, 25, 641–655. [Google Scholar] [CrossRef]
- Loesche, M.; Gardner, S.E.; Kalan, L.; Horwinski, J.; Zheng, Q.; Hodkinson, B.P.; Tyldsley, A.S.; Franciscus, C.L.; Hillis, S.L.; Mehta, S.; et al. Temporal Stability in Chronic Wound Microbiota Is Associated with Poor Healing. J. Investig. Dermatol. 2017, 137, 237–244. [Google Scholar] [CrossRef]
- Mamdoh, H.; Hassanein, K.M.; Eltoony, L.F.; Khalifa, W.A.; Hamed, E.; Alshammari, T.O.; El-Kareem, D.M.A.; El-Mokhtar, M.A. Clinical and Bacteriological Analyses of Biofilm-Forming Staphylococci Isolated from Diabetic Foot Ulcers. Infect. Drug Resist. 2023, 16, 1737–1750. [Google Scholar] [CrossRef]
- Taki, E.; Jabalameli, F.; Tehrani, M.R.M.; Feizabadi, M.M.; Halimi, S.; Sanjari, M.; Amini, M.R.; Beigverdi, R.; Emaneini, M. Molecular Characteristics of Staphylococcus aureus Strains Isolated from Nasal Cavity and Wound Infections among Diabetic Patients. Curr. Microbiol. 2023, 80, 147. [Google Scholar] [CrossRef]
- Monteiro-Soares, M.; Russell, D.; Boyko, E.J.; Jeffcoate, W.; Mills, J.L.; Morbach, S.; Game, F. Guidelines on the Classification of Diabetic Foot Ulcers (IWGDF 2019). Diabetes. Metab. Res. Rev. 2020, 36, e3273. [Google Scholar] [CrossRef]
- Shettigar, K.; Jain, S.; Bhat, D.V.; Acharya, R.; Ramachandra, L.; Satyamoorthy, K.; Murali, T.S. Virulence Determinants in Clinical Staphylococcus aureus from Monomicrobial and Polymicrobial Infections of Diabetic Foot Ulcers. J. Med. Microbiol. 2016, 65, 1392–1404. [Google Scholar] [CrossRef]
- Djahmi, N.; Messad, N.; Nedjai, S.; Moussaoui, A.; Mazouz, D.; Richard, J.L.; Sotto, A.; Lavigne, J.P. Molecular Epidemiology of Staphylococcus aureus Strains Isolated from Inpatients with Infected Diabetic Foot Ulcers in an Algerian University Hospital. Clin. Microbiol. Infect. 2013, 19, E398–E404. [Google Scholar] [CrossRef] [PubMed]
- Mottola, C.; Semedo-Lemsaddek, T.; Mendes, J.J.; Melo-Cristino, J.; Tavares, L.; Cavaco-Silva, P.; Oliveira, M. Molecular Typing, Virulence Traits and Antimicrobial Resistance of Diabetic Foot Staphylococci. J. Biomed. Sci. 2016, 23, 33. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, S.; Pané-Farré, J.; Kohler, C.; Hecker, M.; Engelmann, S. Anaerobic Gene Expression in Staphylococcus aureus. J. Bacteriol. 2007, 189, 4275–4289. [Google Scholar] [CrossRef]
- Kinkel, T.L.; Roux, C.M.; Dunman, P.M.; Fang, F.C. The Staphylococcus aureus SrrAB Two-Component System Promotes Resistance to Nitrosative Stress and Hypoxia. MBio 2013, 4, e00696-13. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Ji, Q.; Jones, M.B.; Deng, X.; Liang, H.; Frank, B.; Telser, J.; Peterson, S.N.; Bae, T.; He, C. AirSR, a [2Fe-2S] Cluster-Containing Two-Component System, Mediates Global Oxygen Sensing and Redox Signaling in Staphylococcus aureus. J. Am. Chem. Soc. 2012, 134, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, D.; Harper, L.; Shopsin, B. Staphylococcus aureus pathogenesis in diverse host environments. Pathog. Dis. 2017, 75, ftx005. [Google Scholar] [CrossRef]
- Vitko, N.P.; Grosser, M.R.; Khatri, D.; Lance, T.R.; Richardson, A.R. Expanded Glucose Import Capability Affords Staphylococcus aureus Optimized Glycolytic Flux during Infection. MBio 2016, 7, e00296-16. [Google Scholar] [CrossRef]
- Dolivo, D.; Lanier, S.; Leung, K.; Mustoe, T.; Hong, S.J.; Galiano, R. Comparative Transcriptomic Adaptations of Staphylococcus aureus to the Wound Environment in Non-Diabetic and Diabetic Mice. Wound Repair Regen. 2022, 30, 541–545. [Google Scholar] [CrossRef]
- Seo, K.S.; Park, N.; Rutter, J.K.; Park, Y.; Baker, C.L.; Thornton, J.A.; Park, J.Y. Role of Glucose-6-Phosphate in Metabolic Adaptation of Staphylococcus aureus in Diabetes. Microbiol. Spectr. 2021, 9, e00857-21. [Google Scholar] [CrossRef]
- Totté, J.E.E.; van der Feltz, W.T.; Bode, L.G.M.; van Belkum, A.; van Zuuren, E.J.; Pasmans, S.G.M.A. A Systematic Review and Meta-Analysis on Staphylococcus aureus Carriage in Psoriasis, Acne and Rosacea. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1069–1077. [Google Scholar] [CrossRef]
- Goering, R.V.; Shawar, R.M.; Scangarella, N.E.; O’Hara, F.P.; Amrine-Madsen, H.; West, J.M.; Dalessandro, M.; Becker, J.A.; Walsh, S.L.; Miller, L.A.; et al. Molecular Epidemiology of Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus Isolates from Global Clinical Trials. J. Clin. Microbiol. 2008, 46, 2842–2847. [Google Scholar] [CrossRef]
- Chroboczek, T.; Boisset, S.; Rasigade, J.P.; Tristan, A.; Bes, M.; Meugnier, H.; Vandenesch, F.; Etienne, J.; Laurent, F. Clonal Complex 398 Methicillin Susceptible Staphylococcus aureus: A Frequent Unspecialized Human Pathogen with Specific Phenotypic and Genotypic Characteristics. PLoS ONE 2013, 8, e68462. [Google Scholar] [CrossRef] [PubMed]
- Kuehnert, M.J.; Kruszon-Moran, D.; Hill, H.A.; McQuillan, G.; McAllister, S.K.; Fosheim, G.; McDougal, L.K.; Chaitram, J.; Jensen, B.; Fridkin, S.K.; et al. Prevalence of Staphylococcus aureus Nasal Colonization in the United States, 2001-2002. J. Infect. Dis. 2006, 193, 172–179. [Google Scholar] [CrossRef]
- Hanis, C.L.; Garrett, K.E.; Essigmann, H.T.; Robinson, D.A.; Gunter, S.M.; Nyitray, A.G.; Brown, E.L. Household Aggregation of Staphylococcus aureus by Clonal Complex and Methicillin Resistance Profiles in Starr County, Texas. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1787–1793. [Google Scholar] [CrossRef] [PubMed]
- Melles, D.C.; Gorkink, R.F.J.; Boelens, H.A.M.; Snijders, S.V.; Peeters, J.K.; Moorhouse, M.J.; van der Spek, P.J.; van Leeuwen, W.B.; Simons, G.; Verbrugh, H.A.; et al. Natural Population Dynamics and Expansion of Pathogenic Clones of Staphylococcus aureus. J. Clin. Investig. 2004, 114, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Fleury, O.M.; McAleer, M.A.; Feuillie, C.; Formosa-Dague, C.; Sansevere, E.; Bennett, D.E.; Towell, A.M.; McLean, W.H.I.; Kezic, S.; Robinson, D.A.; et al. Clumping Factor B Promotes Adherence of Staphylococcus aureus to Corneocytes in Atopic Dermatitis. Infect. Immun. 2017, 85, e00994-16. [Google Scholar] [CrossRef] [PubMed]
- Harkins, C.P.; Pettigrew, K.A.; Oravcová, K.; Gardner, J.; Hearn, R.M.R.; Rice, D.; Mather, A.E.; Parkhill, J.; Brown, S.J.; Proby, C.M.; et al. The Microevolution and Epidemiology of Staphylococcus aureus Colonization during Atopic Eczema Disease Flare. J. Investig. Dermatol. 2018, 138, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.S.; Lee, J.Y.; Baek, J.Y.; Peck, K.R.; Rhee, J.Y.; Kwon, K.T.; Heo, S.T.; Ahn, K.M.; Song, J.H. Characterization of Staphylococcus aureus Nasal Carriage from Children Attending an Outpatient Clinic in Seoul, Korea. Microb. Drug Resist. 2008, 14, 37–44. [Google Scholar] [CrossRef]
- Sarkar, A.; Raji, A.; Garaween, G.; Soge, O.; Rey-Ladino, J.; Al-Kattan, W.; Shibl, A.; Senok, A. Antimicrobial Resistance and Virulence Markers in Methicillin Sensitive Staphylococcus aureus Isolates Associated with Nasal Colonization. Microb. Pathog. 2016, 93, 8–12. [Google Scholar] [CrossRef]
- Mobasherizadeh, S.; Shojaei, H.; Azadi, D.; Havaei, S.A.; Rostami, S. Molecular Characterization and Genotyping of Methicillinresistant Staphylococcus aureus in Nasal Carriage of Healthy Iranian Children. J. Med. Microbiol. 2019, 68, 374–378. [Google Scholar] [CrossRef]
- Aqel, A.A.; Alzoubi, H.M.; Vickers, A.; Pichon, B.; Kearns, A.M. Molecular Epidemiology of Nasal Isolates of Methicillin-Resistant Staphylococcus aureus from Jordan. J. Infect. Public Health 2015, 8, 90–97. [Google Scholar] [CrossRef]
- Aravena Molló, C.; Cáceres, J.; Adolfo Bastías, A.; Opazo, J.F.; Magna, Y.; Saralegui, C.; Quintana, C.; Del Campo, R. Nasal Carriage, Antibiotype and Genotype of Isolated Staphylococcus aureus from Medicine and Nursing Students of Campus San Felipe, University of Valparaiso, Chile, during 2017. Rev. Chil. Infectol. 2021, 38, 774–782. [Google Scholar] [CrossRef]
- Neto, E.D.A.; Guerrero, J.; Snyder, R.E.; Pereira, R.F.A.; de Fátima Nogueira de Freitas, M.; Silva-Santana, G.; Riley, L.W.; Aguiar-Alves, F. Genotypic Distribution of Staphylococcus aureus Colonizing Children and Adolescents in Daycare Centers, an Outpatient Clinic, and Hospitals in a Major Brazilian Urban Setting. Diagn. Microbiol. Infect. Dis. 2020, 97, 115058. [Google Scholar] [CrossRef]
- Clausen, M.L.; Edslev, S.M.; Nørreslet, L.B.; Sørensen, J.A.; Andersen, P.S.; Agner, T. Temporal Variation of Staphylococcus aureus Clonal Complexes in Atopic Dermatitis: A Follow-up Study. Br. J. Dermatol. 2019, 180, 181–186. [Google Scholar] [CrossRef]
- Rojo, A.; Aguinaga, A.; Monecke, S.; Yuste, J.R.; Gastaminza, G.; España, A. Staphylococcus aureus Genomic Pattern and Atopic Dermatitis: May Factors Other than Superantigens Be Involved? Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 651–658. [Google Scholar] [CrossRef]
- Kim, D.W.; Park, J.Y.; Park, K.D.; Kim, T.H.; Lee, W.J.; Lee, S.J.; Kim, J. Are There Predominant Strains and Toxins of Staphylococcus aureus in Atopic Dermatitis Patients? Genotypic Characterization and Toxin Determination of S. aureus Isolated in Adolescent and Adult Patients with Atopic Dermatitis. J. Dermatol. 2009, 36, 75–81. [Google Scholar] [CrossRef]
- Cavalcante, F.S.; Saintive, S.; Carvalho Ferreira, D.; Rocha Silva, A.B.; Guimarães, L.C.; Braga, B.S.; Dios Abad, E.d.; Ribeiro, M.; Netto dos Santos, K.R. Methicillin-Resistant Staphylococcus aureus from Infected Skin Lesions Present Several Virulence Genes and Are Associated with the CC30 in Brazilian Children with Atopic Dermatitis. Virulence 2021, 12, 260–269. [Google Scholar] [CrossRef]
- Benito, D.; Aspiroz, C.; Gilaberte, Y.; Sanmartín, R.; Hernández-Martin, Á.; Alonso, M.; Gómez, P.; Lozano, C.; Torres, C. Genetic Lineages and Antimicrobial Resistance Genotypes in Staphylococcus aureus from Children with Atopic Dermatitis: Detection of Clonal Complexes CC1, CC97 and CC398. J. Chemother. 2016, 28, 359–366. [Google Scholar] [CrossRef]
- Ogonowska, P.; Szymczak, K.; Empel, J.; Urbaś, M.; Woźniak-Pawlikowska, A.; Barańska-Rybak, W.; Świetlik, D.; Nakonieczna, J. Staphylococcus aureus from Atopic Dermatitis Patients: Its Genetic Structure and Susceptibility to Phototreatment. Microbiol. Spectr. 2023, 11, e04598-22. [Google Scholar] [CrossRef]
- Clausen, M.L.; Edslev, S.M.; Andersen, P.S.; Clemmensen, K.; Krogfelt, K.A.; Agner, T. Staphylococcus aureus Colonization in Atopic Eczema and Its Association with Filaggrin Gene Mutations. Br. J. Dermatol. 2017, 177, 1394–1400. [Google Scholar] [CrossRef]
- Van Mierlo, M.M.F.; Pardo, L.M.; Fieten, K.B.; Van Den Broek, T.J.; Schuren, F.H.J.; Van Geel, M.; Pasmans, S.G.M.A. The Skin and Nose Microbiome and Its Association with Filaggrin Gene Mutations in Pediatric Atopic Dermatitis. Dermatology 2022, 238, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.L.; Villarreal, M.; Jepson, B.; Rafaels, N.; David, G.; Hanifin, J.; Taylor, P.; Boguniewicz, M.; Yoshida, T.; De Benedetto, A.; et al. Patients with Atopic Dermatitis Colonized with Staphylococcus aureus Have a Distinct Phenotype and Endotype. J. Investig. Dermatol. 2018, 138, 2224–2233. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.; Rocha, L.; Sokhatska, O.; Soares, J.; Tavaria, F.; Correia, O.; Pintado, M.; Fernandes, S.; Delgado, L.; Moreira, A. Filaggrin Polymorphism Pro478Ser Is Associated with the Severity of Atopic Dermatitis and Colonization by Staphylococcal Aureus. J. Investig. Allergol. Clin. Immunol. 2016, 26, 70–72. [Google Scholar] [CrossRef]
- Van Mierlo, M.M.F.; Pasmans, S.G.M.A.; Totté, J.E.E.; De Wit, J.; Herpers, B.L.; Vos, M.C.; Klaassen, C.H.W.; Pardo, L.M. Temporal Variation in Staphylococcus aureus Protein a Genotypes from Nose and Skin in Atopic Dermatitis Patients. Dermatology 2021, 237, 506–512. [Google Scholar] [CrossRef]
- Lomholt, H.; Andersen, K.E.; Kilian, M. Staphylococcus aureus Clonal Dynamics and Virulence Factors in Children with Atopic Dermatitis. J. Investig. Dermatol. 2005, 125, 977–982. [Google Scholar] [CrossRef]
- Stalder, J.F.; Taïeb, A.; Atherton, D.J.; Bieber, P.; Bonifazi, E.; Broberg, A.; Calza, A.; Coleman, R.; De Prost, Y.; Stalder, J.F.; et al. Severity Scoring of Atopic Dermatitis: The SCORAD Index: Consensus Report of the European Task Force on Atopic Dermatitis. Dermatology 1993, 186, 23–31. [Google Scholar] [CrossRef]
- Arfaoui, A.; Sallem, R.B.; Fernández-Fernández, R.; Eguizábal, P.; Dziri, R.; Abdullahi, I.N.; Sayem, N.; Ben Khelifa Melki, S.; Ouzari, H.I.; Torres, C.; et al. Methicillin-Resistant Staphylococcus aureus from Diabetic Foot Infections in a Tunisian Hospital with the First Detection of MSSA CC398-T571. Antibiotics 2022, 11, 1755. [Google Scholar] [CrossRef]
- Senneville, E.; Brière, M.; Neut, C.; Messad, N.; Lina, G.; Richard, J.L.; Sotto, A.; Lavigne, J.P.; Bouziges, N.; Jourdan, N.; et al. First Report of the Predominance of Clonal Complex 398 Staphylococcus aureus Strains in Osteomyelitis Complicating Diabetic Foot Ulcers: A National French Study. Clin. Microbiol. Infect. 2014, 20, O274–O277. [Google Scholar] [CrossRef]
- Wehrhahn, M.C.; Robinson, J.O.; Pascoe, E.M.; Coombs, G.W.; Pearson, J.C.; O’Brien, F.G.; Tan, H.L.; New, D.; Salvaris, P.; Salvaris, R.; et al. Illness Severity in Community-Onset Invasive Staphylococcus aureus Infection and the Presence of Virulence Genes. J. Infect. Dis. 2012, 205, 1840–1848. [Google Scholar] [CrossRef]
- Bouiller, K.; Hocquet, D.; Sauget, M.; Bertrand, X.; Chirouze, C. Epidemiology and Risk Factors of Staphylococcus aureus CC398 Bone and Joint Infections. BMC Infect. Dis. 2020, 20, 384. [Google Scholar] [CrossRef]
- Bouiller, K.; Bertrand, X.; Hocquet, D.; Chirouze, C. Human Infection of Methicillin-susceptible Staphylococcus aureus Cc398: A Review. Microorganisms 2020, 8, 1737. [Google Scholar] [CrossRef]
- Sotto, A.; Richard, J.L.; Messad, N.; Molinari, N.; Jourdan, N.; Schuldiner, S.; Sultan, A.; Carrière, C.; Canivet, B.; Landraud, L.; et al. Distinguishing Colonization from Infection with Staphylococcus aureus in Diabetic Foot Ulcers with Miniaturized Oligonucleotide Arrays: A French Multicenter Study. Diabetes Care 2012, 35, 617–623. [Google Scholar] [CrossRef]
- Haleem, A.; Schultz, J.S.; Heilmann, K.P.; Dohrn, C.L.; Diekema, D.J.; Gardner, S.E. Concordance of Nasal and Diabetic Foot Ulcer Staphylococcal Colonization. Diagn. Microbiol. Infect. Dis. 2014, 79, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.; Hamilton, E.J.; Makepeace, A.; Davis, W.A.; Latkovic, E.; Lim, E.M.; Dyer, J.R.; Davis, T.M.E. Prevalence, Risk Factors and Sequelae of Staphylococcus aureus Carriage in Diabetes: The Fremantle Diabetes Study Phase II. J. Diabetes Complicat. 2015, 29, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Planet, P.J.; Parker, D.; Ruff, N.L.; Shinefield, H.R. Revisiting Bacterial Interference in the Age of Methicillin-Resistant Staphylococcus aureus. Pediatr. Infect. Dis. J. 2019, 38, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Dunyach-Remy, C.; Courtais-Coulon, C.; DeMattei, C.; Jourdan, N.; Schuldiner, S.; Sultan, A.; Carrière, C.; Alonso, S.; Sotto, A.; Lavigne, J.P. Link between Nasal Carriage of Staphylococcus aureus and Infected Diabetic Foot Ulcers. Diabetes Metab. 2017, 43, 167–171. [Google Scholar] [CrossRef]
- Lavigne, J.P.; Hosny, M.; Dunyach-Remy, C.; Boutet-Dubois, A.; Schuldiner, S.; Cellier, N.; Yahiaoui-Martinez, A.; Molle, V.; La Scola, B.; Marchandin, H.; et al. Long-Term Intrahost Evolution of Staphylococcus aureus among Diabetic Patients with Foot Infections. Front. Microbiol. 2021, 12, 741406. [Google Scholar] [CrossRef] [PubMed]
- Kahl, B.C.; Becker, K.; Friedrich, A.W.; Clasen, J.; Sinha, B.; Von Eiff, C.; Peters, G. Agr-Dependent Bacterial Interference Has No Impact on Long-Term Colonization of Staphylococcus aureus during Persistent Airway Infection of Cystic Fibrosis Patients. J. Clin. Microbiol. 2003, 41, 5199–5201. [Google Scholar] [CrossRef]
- Garofalo, A.; Giai, C.; Lattar, S.; Gardella, N.; Mollerach, M.; Kahl, B.C.; Becker, K.; Prince, A.S.; Sordelli, D.O.; Gómez, M.I. The Length of the Staphylococcus aureus Protein a Polymorphic Region Regulates Inflammation: Impact on Acute and Chronic Infection. J. Infect. Dis. 2012, 206, 81–90. [Google Scholar] [CrossRef]
- Casino, P.; Rubio, V.; Marina, A. The Mechanism of Signal Transduction by Two-Component Systems. Curr. Opin. Struct. Biol. 2010, 20, 763–771. [Google Scholar] [CrossRef]
- Haag, A.F.; Bagnoli, F. The Role of Two-Component Signal Transduction Systems in Staphylococcus aureus Virulence Regulation. Curr. Top. Microbiol. Immunol. 2017, 409, 145–198. [Google Scholar] [CrossRef]
- Giraudo, A.T.; Calzolari, A.; Cataldi, A.A.; Bogni, C.; Nagel, R. The Sae Locus of Staphylococcus aureus Encodes a Two-Component Regulatory System. FEMS Microbiol. Lett. 1999, 177, 15–22. [Google Scholar] [CrossRef]
- Norvick, R.P.; Jiang, D. The Staphylococcal SaeRS System Coordinates Environmental Signals Agr Quorum Sensing. Microbiology 2003, 149, 2709–2717. [Google Scholar] [CrossRef]
- Cheung, A.L.; Bayer, A.S.; Zhang, G.; Gresham, H.; Xiong, Y.-Q. Regulation of Virulence Determinants in Vitro and in Vivo in Staphylococcus aureus. FEMS Immunol. Med. Microbiol. 2004, 40, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.; Geisinger, E. Quorum Sensing in Staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef] [PubMed]
- Peschel, A.; Otto, M. Phenol-Soluble Modulins and Staphylococcal Infection. Nat. Rev. Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Queck, S.Y.; Jameson-Lee, M.; Villaruz, A.E.; Bach, T.H.L.; Khan, B.A.; Sturdevant, D.E.; Ricklefs, S.M.; Li, M.; Otto, M. RNAIII-Independent Target Gene Control by the Agr Quorum-Sensing System: Insight into the Evolution of Virulence Regulation in Staphylococcus aureus. Mol. Cell 2008, 32, 150–158. [Google Scholar] [CrossRef]
- Regassa, L.B.; Novick, R.P.; Betley, M.J. Glucose and Nonmaintained PH Decrease Expression of the Accessory Gene Regulator (Agr) in Staphylococcus aureus. Infect. Immun. 1992, 60, 3381–3388. [Google Scholar] [CrossRef]
- Jeong, D.W.; Cho, H.; Jones, M.B.; Shatzkes, K.; Sun, F.; Ji, Q.; Liu, Q.; Peterson, S.N.; He, C.; Bae, T. The Auxiliary Protein Complex SaePQ Activates the Phosphatase Activity of Sensor Kinase SaeS in the SaeRS Two-Component System of Staphylococcus aureus. Mol. Microbiol. 2012, 86, 331–348. [Google Scholar] [CrossRef]
- DuMont, A.L.; Nygaard, T.K.; Watkins, R.L.; Smith, A.; Kozhaya, L.; Kreiswirth, B.N.; Shopsin, B.; Unutmaz, D.; Voyich, J.M.; Torres, V.J. Characterization of a New Cytotoxin That Contributes to Staphylococcus aureus Pathogenesis. Mol. Microbiol. 2011, 79, 814–825. [Google Scholar] [CrossRef]
- Voyich, J.M.; Braughton, K.R.; Sturdevant, D.E.; Whitney, A.R.; Saïd-Salim, B.; Porcella, S.F.; Long, R.D.; Dorward, D.W.; Gardner, D.J.; Kreiswirth, B.N.; et al. Insights into Mechanisms Used by Staphylococcus aureus to Avoid Destruction by Human Neutrophils. J. Immunol. 2005, 175, 3907–3919. [Google Scholar] [CrossRef] [PubMed]
- Voyich, J.M.; Vuong, C.; DeWald, M.; Nygaard, T.K.; Kocianova, S.; Griffith, S.; Jones, J.; Iverson, C.; Sturdevant, D.E.; Braughton, K.R.; et al. The SaeR/S Gene Regulatory System Is Essential for Innate Immune Evasion by Staphylococcus aureus. J. Infect. Dis. 2009, 199, 1698–1706. [Google Scholar] [CrossRef] [PubMed]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus Virulence. Microbiol. Spectr. 2019, 6, GPP3-0031-2018. [Google Scholar] [CrossRef]
- Ogonowska, P.; Gilaberte, Y.; Barańska-Rybak, W.; Nakonieczna, J. Colonization with Staphylococcus aureus in Atopic Dermatitis Patients: Attempts to Reveal the Unknown. Front. Microbiol. 2021, 11, 567090. [Google Scholar] [CrossRef]
- Breuer, K.; Wittmann, M.; Kempe, K.; Kapp, A.; Mai, U.; Dittrich-Breiholz, O.; Kracht, M.; Mrabet-Dahbi, S.; Werfel, T. Alpha-Toxin Is Produced by Skin Colonizing Staphylococcus aureus and Induces a T Helper Type 1 Response in Atopic Dermatitis. Clin. Exp. Allergy 2005, 35, 1088–1095. [Google Scholar] [CrossRef]
- Conte, A.L.; Brunetti, F.; Marazzato, M.; Longhi, C.; Maurizi, L.; Raponi, G.; Palamara, A.T.; Grassi, S.; Conte, M.P. Atopic Dermatitis-Derived Staphylococcus aureus Strains: What Makes Them Special in the Interplay with the Host. Front. Cell. Infect. Microbiol. 2023, 13, 1194254. [Google Scholar] [CrossRef]
- Nakamura, Y.; Takahashi, H.; Takaya, A.; Inoue, Y.; Katayama, Y.; Kusuya, Y.; Shoji, T.; Takada, S.; Nakagawa, S.; Oguma, R.; et al. Staphylococcus Agr Virulence Is Critical for Epidermal Colonization and Associates with Atopic Dermatitis Development. Sci. Transl. Med. 2020, 12, eaay4068. [Google Scholar] [CrossRef]
- Tamai, M.; Yamazaki, Y.; Ito, T.; Nakagawa, S.; Nakamura, Y. Pathogenic Role of the Staphylococcal Accessory Gene Regulator Quorum Sensing System in Atopic Dermatitis. Front. Cell. Infect. Microbiol. 2023, 13, 1178650. [Google Scholar] [CrossRef]
- Le, K.Y.; Otto, M. Quorum-Sensing Regulation in Staphylococci—An Overview. Front. Microbiol. 2015, 6, 1174. [Google Scholar] [CrossRef]
- Nakagawa, S.; Matsumoto, M.; Katayama, Y.; Oguma, R.; Wakabayashi, S.; Nygaard, T.; Saijo, S.; Inohara, N.; Otto, M.; Matsue, H.; et al. Staphylococcus aureus Virulent PSMα Peptides Induce Keratinocyte Alarmin Release to Orchestrate IL-17-Dependent Skin Inflammation. Cell Host Microbe 2017, 22, 667–677.e5. [Google Scholar] [CrossRef] [PubMed]
- Niebuhr, M.; Mamerow, D.; Heratizadeh, A.; Satzger, I.; Werfel, T. Staphylococcal α-Toxin Induces a Higher T Cell Proliferation and Interleukin-31 in Atopic Dermatitis. Int. Arch. Allergy Immunol. 2011, 156, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Damour, A.; Robin, B.; Deroche, L.; Broutin, L.; Bellin, N.; Verdon, J.; Lina, G.; Leclère, F.M.; Garcia, M.; Cremniter, J.; et al. Phenol-Soluble Modulins α Are Major Virulence Factors of Staphylococcus aureus Secretome Promoting Inflammatory Response in Human Epidermis. Virulence 2021, 12, 2474–2492. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.L.; Eberhardt, K.; Heinrichs, J.H. Regulation of Protein a Synthesis by the Sar and Agr Loci of Staphylococcus aureus. Infect. Immun. 1997, 65, 2243–2249. [Google Scholar] [CrossRef]
- Ledo, C.; Gonzalez, C.D.; Garofalo, A.; Sabbione, F.; Keitelman, I.A.; Giai, C.; Stella, I.; Trevani, A.S.; Gómez, M.I. Protein A Modulates Neutrophil and Keratinocyte Signaling and Survival in Response to Staphylococcus aureus. Front. Immunol. 2021, 11, 524180. [Google Scholar] [CrossRef]
- Nakamura, Y.; Oscherwitz, J.; Cease, K.B.; Chan, S.M.; Muñoz-Planillo, R.; Hasegawa, M.; Villaruz, A.E.; Cheung, G.Y.C.; McGavin, M.J.; Travers, J.B.; et al. Staphylococcus δ-Toxin Induces Allergic Skin Disease by Activating Mast Cells. Nature 2013, 503, 397–401. [Google Scholar] [CrossRef]
- Miedzobrodzki, J.; Kaszycki, P.; Bialecka, A.; Kasprowicz, A. Proteolytic Activity of Staphylococcus aureus Strains Isolated from the Colonized Skin of Patients with Acute-Phase Atopic Dermatitis. Eur. J. Clin. Microbiol. Infect. Dis. 2002, 21, 269–276. [Google Scholar] [CrossRef]
- Williams, M.R.; Nakatsuji, T.; Sanford, J.A.; Vrbanac, A.F.; Gallo, R.L. Staphylococcus aureus Induces Increased Serine Protease Activity in Keratinocytes. J. Investig. Dermatol. 2017, 137, 377–384. [Google Scholar] [CrossRef]
- Geiger, T.; Goerke, C.; Mainiero, M.; Kraus, D.; Wolz, C. The Virulence Regulator Sae of Staphylococcus aureus: Promoter Activities and Response to Phagocytosis-Related Signals. J. Bacteriol. 2008, 190, 3419–3428. [Google Scholar] [CrossRef]
- Geiger, T.; Francois, P.; Liebeke, M.; Fraunholz, M.; Goerke, C.; Krismer, B.; Schrenzel, J.; Lalk, M.; Wolz, C. The Stringent Response of Staphylococcus aureus and Its Impact on Survival after Phagocytosis through the Induction of Intracellular PSMs Expression. PLoS Pathog. 2012, 8, e1003016. [Google Scholar] [CrossRef]
- Soong, G.; Paulino, F.; Wachtel, S.; Parker, D.; Wickersham, M.; Zhang, D.; Brown, A.; Lauren, C.; Dowd, M.; West, E.; et al. Methicillin-Resistant Staphylococcus aureus Adaptation to Human. mBio 2015, 6, e00289-15. [Google Scholar] [CrossRef]
- Classen, A.; Kalali, B.N.; Schnopp, C.; Andres, C.; Aguilar-Pimentel, J.a.; Ring, J.; Ollert, M.; Mempel, M. TNF Receptor I on Human Keratinocytes Is a Binding Partner for Staphylococcal Protein A Resulting in the Activation of NF Kappa B, AP-1, and Downstream Gene Transcription. Exp. Dermatol. 2011, 20, 48–52. [Google Scholar] [CrossRef]
- Ezepchuk, Y.V.; Leung, D.Y.M.; Middleton, M.H.; Bina, P.; Reiser, R.; Norris, D.A. Staphylococcal Toxins and Protein A Differentially Induce Cytotoxicity and Release of Tumor Necrosis Factor-α from Human Keratinocytes. J. Investig. Dermatol. 1996, 107, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.H.; Lee, J.H.; Kim, S.I.; Choi, C.W.; Park, T.I.; Jung, H.R.; Cho, J.W.; Kim, S.H.; Lee, J.C. Staphylococcus aureus-Derived Membrane Vesicles Exacerbate Skin Inflammation in Atopic Dermatitis. Clin. Exp. Allergy 2017, 47, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.I.; Lee, A.; Reddy, B.; Muir, A.; Soong, G.; Pitt, A.; Cheung, A.; Prince, A. Staphylococcus aureus Protein A Induces Airway Epithelial Inflammatory Responses by Activating TNFR1. Nat. Med. 2004, 10, 842–848. [Google Scholar] [CrossRef]
- Gómez, M.I.; O’Seaghdha, M.; Magargee, M.; Foster, T.J.; Prince, A.S. Staphylococcus aureus Protein A Activates TNFR1 Signaling through Conserved IgG Binding Domains. J. Biol. Chem. 2006, 281, 20190–20196. [Google Scholar] [CrossRef]
- Lopatnikova, J.A.; Alshevskaya, A.A.; Krugleeva, O.L.; Nepomnyschih, V.M.; Gladkikh, V.S.; Lukinov, V.L.; Karaulov, A.V.; Sennikov, S.V. Expression of TNFα Receptors on Immunocompetent Cells Is Increased in Atopic Dermatitis. Int. Arch. Allergy Immunol. 2017, 174, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Merino, N.; Toledo-Arana, A.; Vergara-Irigaray, M.; Valle, J.; Solano, C.; Calvo, E.; Lopez, J.A.; Foster, T.J.; Penadés, J.R.; Lasa, I. Protein A-Mediated Multicellular Behavior in Staphylococcus aureus. J. Bacteriol. 2009, 191, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Sonesson, A.; Przybyszewska, K.; Eriksson, S.; Mörgelin, M.; Kjellström, S.; Davies, J.; Potempa, J.; Schmidtchen, A. Identification of Bacterial Biofilm and the Staphylococcus aureus Derived Protease, Staphopain, on the Skin Surface of Patients with Atopic Dermatitis. Sci. Rep. 2017, 7, 8689. [Google Scholar] [CrossRef]
- Williams, M.R.; Costa, S.K.; Zaramela, L.S.; Khalil, S.; Todd, D.A.; Winter, H.L.; Sanford, J.A.; O’Neill, A.M.; Liggins, M.C.; Nakatsuji, T.; et al. Quorum Sensing between Bacterial Species on the Skin Protects against Epidermal Injury in Atopic Dermatitis. Sci. Transl. Med. 2019, 11, eaat8329. [Google Scholar] [CrossRef]
- Ji, G.; Beavis, R.; Novick, R.P. Bacterial Interference Caused by Autoinducing Peptide Variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P.; Muir, T.W. Virulence Gene Regulation by Peptides in Staphylococci and Other Gram- Positive Bacteria. Curr. Opin. Microbiol. 1999, 2, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Liu, X.; Li, Y.; Luo, L.; Yuan, W.; Chen, B.; Liang, G.; Shen, R.; Li, H.; Huang, S.; et al. Advanced Glycation End Products Enhance Biofilm Formation by Promoting Extracellular DNA Release Through SigB Upregulation in Staphylococcus aureus. Front. Microbiol. 2020, 11, 1479. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, L.D.; Lopes, A.C.P.; Teixeira, M.M.; da Silva, J.M.A.; Silva, L.O.; de Almeida, J.B.; Campos, G.B.; Teodósio, R.; Marques, L.M. Clinical and Microbiological Profile of Diabetic Foot Ulcers Infected with Staphylococcus aureus in a Regional General Hospital in Bahia, Brazil. Int. J. Low. Extrem. Wounds 2021, 34747264. [Google Scholar] [CrossRef]
- Mitsuhashi, T.; Nakayama, H.; Itoh, T.; Kuwajima, S.; Aoki, S.; Atsumi, T.; Koike, T. Immunochemical Detection of Advanced Glycation End Products in Renal Cortex from STZ-Induced Diabetic Rat. Diabetes 1993, 42, 826–832. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Shu, J.C.; Lin, M.H.; Chong, K.Y.; Chen, C.C.; Wen, S.M.; Hsieh, Y.T.; Liao, W.T. High Glucose Concentration Promotes Vancomycin-Enhanced Biofilm Formation of Vancomycin-Non-Susceptible Staphylococcus aureus in Diabetic Mice. PLoS ONE 2015, 10, e0134852. [Google Scholar] [CrossRef]
- Seidl, K.; Goerke, C.; Wolz, C.; Mack, D.; Berger-Bächi, B.; Bischoff, M. Staphylococcus aureus CcpA Affects Biofilm Formation. Infect. Immun. 2008, 76, 2044–2050. [Google Scholar] [CrossRef]
- Vitko, N.P.; Spahich, N.A.; Richardson, A.R. Glycolytic Dependency of High-Level Nitric Oxide Resistance and Virulence in Staphylococcus aureus. MBio 2015, 6, 2–11. [Google Scholar] [CrossRef]
- Vlassara, H.; Uribarri, J. Advanced Glycation End Products (AGE) and Diabetes: Cause, Effect, or Both? Curr. Diab. Rep. 2014, 14, 453. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, C.; Wang, C.; Liu, D.; Lao, G.; Liang, Y.; Sun, K.; Luo, H.; Tan, Q.; Ren, M.; et al. AGE-Induced Keratinocyte MMP-9 Expression Is Linked to TET2-Mediated CpG Demethylation. Wound Repair Regen. 2016, 24, 489–500. [Google Scholar] [CrossRef]
- Haasbroek, K.; Yagi, M.; Yonei, Y. Staphylococcus aureus Biofilm Inhibiting Activity of Advanced Glycation Endproduct Crosslink Breaking and Glycation Inhibiting Compounds. Antibiotics 2022, 11, 1412. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Grajera, M.; Pacha-Olivenza, M.A.; Gallardo-Moreno, A.M.; González-Martín, M.L.; Pérez-Giraldo, C.; Fernández-Calderón, M.C. Modification of Physico-Chemical Surface Properties and Growth of Staphylococcus aureus under Hyperglycemia and Ketoacidosis Conditions. Colloids Surfaces B Biointerfaces 2022, 209, 112137. [Google Scholar] [CrossRef] [PubMed]
- Tankersley, A.; Frank, M.B.; Bebak, M.; Brennan, R. Early Effects of Staphylococcus aureus Biofilm Secreted Products on Inflammatory Responses of Human Epithelial Keratinocytes. J. Inflamm. 2014, 11, 17. [Google Scholar] [CrossRef]
- Schierle, C.F.; De La Garza, M.; Mustoe, T.A.; Galiano, R.D. Staphylococcal Biofilms Impair Wound Healing by Delaying Reepithelialization in a Murine Cutaneous Wound Model. Wound Repair Regen. 2009, 17, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Aswathanarayan, J.B.; Rao, P.; Hm, S.; Gs, S.; Rai, R.V. Biofilm-Associated Infections in Chronic Wounds and Their Management. Adv. Exp. Med. Biol. 2023, 1370, 55–75. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Usui, M.L.; Underwood, R.A.; Singh, P.K.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Delayed Healing in Biofilm-Colonized Db/Db Mouse Wounds. Wound Repair Regen. 2012, 20, 342–352. [Google Scholar] [CrossRef]
- Mauriello, C.T.; Hair, P.S.; Rohn, R.D.; Rister, N.S.; Krishna, N.K.; Cunnion, K.M. Hyperglycemia Inhibits Complement-Mediated Immunological Control of S. Aureus in a Rat Model of Peritonitis. J. Diabetes Res. 2014, 2014, 762051. [Google Scholar] [CrossRef]
- Pouget, C.; Gustave, C.A.; Ngba-Essebe, C.; Laurent, F.; Lemichez, E.; Tristan, A.; Sotto, A.; Dunyach-Rémy, C.; Lavigne, J.P. Adaptation of Staphylococcus aureus in a Medium Mimicking a Diabetic Foot Environment. Toxins 2021, 13, 230. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Löffler, B.; Proctor, R.A. Persistence of Staphylococcus aureus: Multiple Metabolic Pathways Impact the Expression of Virulence Factors in Small-Colony Variants (SCVs). Front. Microbiol. 2020, 11, 1028. [Google Scholar] [CrossRef]
- Lee, J.; Zilm, P.S.; Kidd, S.P. Novel Research Models for Staphylococcus aureus Small Colony Variants (SCV) Development: Co-Pathogenesis and Growth Rate. Front. Microbiol. 2020, 11, 321. [Google Scholar] [CrossRef]
- Keren, I.; Kaldalu, N.; Spoering, A.; Wang, Y.; Lewis, K. Persister Cells and Tolerance to Antimicrobials. FEMS Microbiol. Lett. 2004, 230, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Conlon, B.P. Staphylococcus aureus Chronic and Relapsing Infections: Evidence of a Role for Persister Cells: An Investigation of Persister Cells, Their Formation and Their Role in S. Aureus Disease. Bioessays 2014, 36, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Huitema, L.; Phillips, T.; Alexeev, V.; Tomic-Canic, M.; Pastar, I.; Igoucheva, O. Intracellular Escape Strategies of Staphylococcus aureus in Persistent Cutaneous Infections. Exp. Dermatol. 2021, 30, 1428–1439. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Mashayamombe, M.; Walsh, T.P.; Kuang, B.K.P.; Pena, G.N.; Vreugde, S.; Cooksley, C.; Carda-Diéguez, M.; Mira, A.; Jesudason, D.; et al. The Bacteriology of Diabetic Foot Ulcers and Infections and Incidence of Staphylococcus aureus Small Colony Variants. J. Med. Microbiol. 2023, 72, 001716. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-García, E.; García-Gonzalez, R.; Reyes-Torres, A.; Resendiz-Albor, A.A.; Salazar-Schettino, P.M. Staphylococcus aureus Small Colony Variants in Diabetic Foot Infections. Diabet. Foot Ankle 2015, 6, 4–8. [Google Scholar] [CrossRef]
- Boles, B.R.; Horswill, A.R. Agr-Mediated Dispersal of Staphylococcus aureus Biofilms. PLoS Pathog. 2008, 4, e1000052. [Google Scholar] [CrossRef]
- Yarwood, J.M.; Bartels, D.J.; Volper, E.M.; Greenberg, E.P. Quorum Sensing in Staphylococcus aureus Biofilms. J. Bacteriol. 2004, 186, 1838–1850. [Google Scholar] [CrossRef]
- Le, K.Y.; Dastgheyb, S.; Ho, T.V.; Otto, M. Molecular Determinants of Staphylococcal Biofilm Dispersal and Structuring. Front. Cell. Infect. Microbiol. 2014, 4, 167. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | MRSA from Hospital Setting | MRSA from Community Setting | MSSA |
|---|---|---|---|
| North America | ST5, ST8, ST36, ST45 | ST8, ST30 | ST1, ST8, ST30, ST45, ST398 (CC398) |
| South America | ST5, ST239 | ST5, ST8, ST30 | ST1, ST8, ST30, ST45 |
| Europe | ST5, ST8, ST22 | ST8, ST30, ST80 | ST1, ST8, ST30, ST45, ST398 (CC398) |
| Asia | ST5, ST22, ST239 | ST8, ST30, ST59, ST88, ST93, ST772 | ST398 (CC398) |
| Australia and New Zealand | ST1, ST22 | ST5, ST93 | -- |
| Africa | ST5, ST239 | ST80, ST88 | ST1, ST8, ST30, ST45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gehrke, A.-K.E.; Giai, C.; Gómez, M.I. Staphylococcus aureus Adaptation to the Skin in Health and Persistent/Recurrent Infections. Antibiotics 2023, 12, 1520. https://doi.org/10.3390/antibiotics12101520
Gehrke A-KE, Giai C, Gómez MI. Staphylococcus aureus Adaptation to the Skin in Health and Persistent/Recurrent Infections. Antibiotics. 2023; 12(10):1520. https://doi.org/10.3390/antibiotics12101520
Chicago/Turabian StyleGehrke, Ana-Katharina E., Constanza Giai, and Marisa I. Gómez. 2023. "Staphylococcus aureus Adaptation to the Skin in Health and Persistent/Recurrent Infections" Antibiotics 12, no. 10: 1520. https://doi.org/10.3390/antibiotics12101520