
Antimicrobial Susceptibility and Genetic Prevalence of Extended-Spectrum β-Lactamases in Gram-Negative Rods Isolated from Clinical Specimens in Pakistan

Abstract
:1. Introduction
2. Results
2.1. Socio-Demographic Data and Distribution of ESBL-Producing Bacterial Isolates
2.2. Susceptibility of ESBL-Producing Gram-Negative Rods (GNRs) to Antibiotics
2.3. Susceptibility of Escherichia coli to Various Antibiotics
2.4. Susceptibility of Klebsiella Species to Various Antibiotics
2.5. Susceptibility of Acinetobacter Species and Others to Various Antibiotics
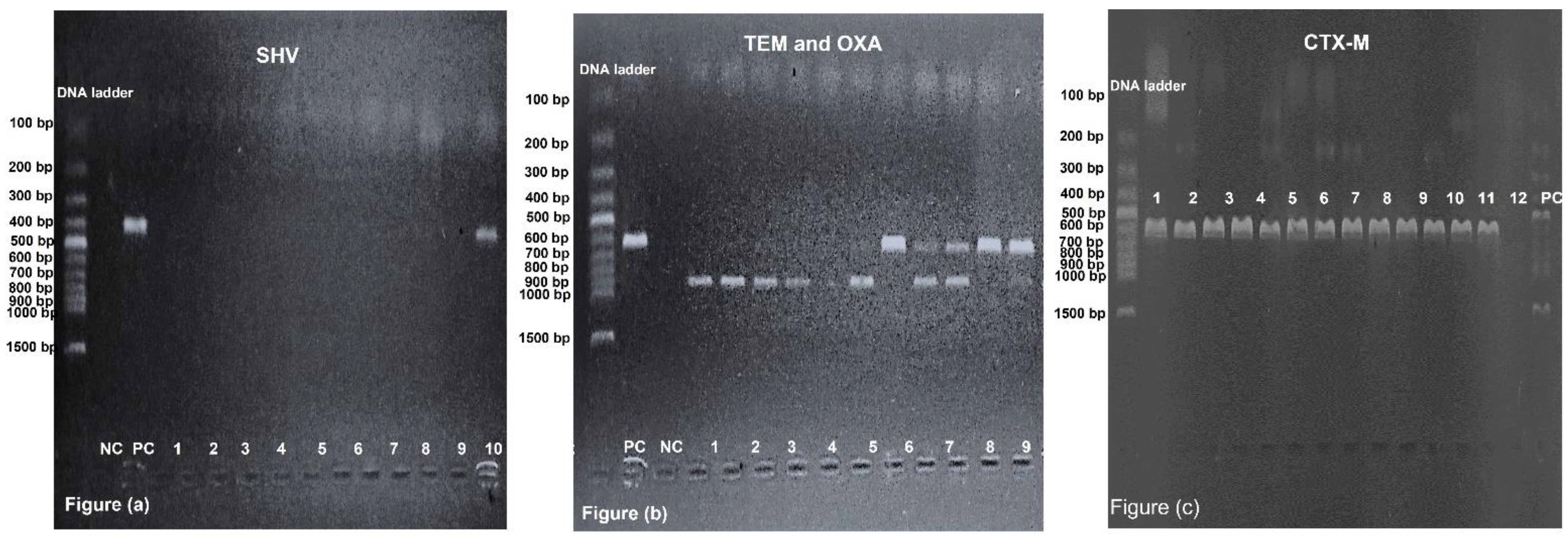
2.6. Prevalence of blaCTX-M, blaOXA, blaTEM and blaSHV among GNRs
2.7. Correlation between Antibiotic Resistance and ESBL Genes
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Transportation and Preservation
4.2. Sample Inoculation and Bacterial Identification
4.3. Bacterial Identification by Gram Staining and Biochemical Test
4.4. Antimicrobial Susceptibility Testing
4.5. Phenotypic Identification of ESBLs
4.6. Molecular Detection of ESBLs
4.7. Quality Control
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bush, K.; Jacoby, G.A.; Medeiros, A.A. A functional classification scheme for beta-lactamases and its correlation with molecular structure. Antimicrob. Agents Chemother. 1995, 39, 1211–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghafourian, S.; Sadeghifard, N.; Soheili, S.; Sekawi, Z. Extended spectrum beta-lactamases: Definition, classification and epidemiology. Curr. Issues Mol. Biol. 2015, 17, 11–22. [Google Scholar] [PubMed]
- Ruppé, É.; Woerther, P.-L.; Barbier, F. Mechanisms of antimicrobial resistance in Gram-negative bacilli. Ann. Intensive Care 2015, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, S.S. What are extended-spectrum beta-lactamases? J. Am. Acad. PAs 2014, 27, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Chong, Y.; Shimoda, S.; Yakushiji, H.; Ito, Y.; Miyamoto, T.; Kamimura, T.; Shimono, N.; Akashi, K. Community spread of extended-spectrum β-lactamase-producing Escherichia coli, Klebsiella pneumoniae and Proteus mirabilis: A long-term study in Japan. J. Med. Microbiol. 2013, 62, 1038–1043. [Google Scholar] [CrossRef]
- Kassakian, S.Z.; Mermel, L.A. Changing epidemiology of infections due to extended spectrum beta-lactamase producing bacteria. Antimicrob. Resist. Infect. Control 2014, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Aidara-Kane, A.; Angulo, F.J.; Conly, J.M.; Minato, Y.; Silbergeld, E.K.; McEwen, S.A.; Collignon, P.J. World Health Organization (WHO) guidelines on use of medically important antimicrobials in food-producing animals. Antimicrob. Resist. Infect. Control 2018, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Datta, N.; Kontomichalou, P. Penicillinase synthesis controlled by infectious R factors in Enterobacteriaceae. Nature 1965, 208, 239–241. [Google Scholar] [CrossRef]
- Knothe, H.; Shah, P.; Krcmery, V.; Antal, M.; Mitsuhashi, S. Transferable resistance to cefotaxime, cefoxitin, cefamandole and cefuroxime in clinical isolates of Klebsiella pneumoniae and Serratia marcescens. Infection 1983, 11, 315–317. [Google Scholar] [CrossRef]
- Philippon, A.; Labia, R.; Jacoby, G. Extended-spectrum beta-lactamases. Antimicrob. Agents Chemother. 1989, 33, 1131–1136. [Google Scholar] [CrossRef]
- Livermore, D.M. Defining an extended-spectrum β-lactamase. Clin. Microbiol. Infect. 2008, 14, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Ambler, R.P.; Coulson, A.F.; Frère, J.-M.; Ghuysen, J.-M.; Joris, B.; Forsman, M.; Levesque, R.C.; Tiraby, G.; Waley, S.G. A standard numbering scheme for the class A beta-lactamases. Biochem. J. 1991, 276 Pt 1, 269. [Google Scholar] [CrossRef]
- Thenmozhi, S.; Moorthy, K.; Sureshkumar, B.; Suresh, M. Antibiotic resistance mechanism of ESBL producing Enterobacteriaceae in clinical field: A review. Int. J. Pure Appl. Biosci. 2014, 2, 207–226. [Google Scholar]
- Adamski, C.J.; Cardenas, A.M.; Brown, N.G.; Horton, L.B.; Sankaran, B.; Prasad, B.V.; Gilbert, H.F.; Palzkill, T. Molecular basis for the catalytic specificity of the CTX-M extended-spectrum β-lactamases. Biochemistry 2015, 54, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Shaikh, S.; Fatima, J.; Shakil, S.; Rizvi, S.M.D.; Kamal, M.A. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J. Biol. Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Balkhed, Å.Ö.; Tärnberg, M.; Monstein, H.-J.; Hällgren, A.; Hanberger, H.; Nilsson, L.E. High frequency of co-resistance in CTX-M-producing Escherichia coli to non-beta-lactam antibiotics, with the exceptions of amikacin, nitrofurantoin, colistin, tigecycline, and fosfomycin, in a county of Sweden. Scand. J. Infect. Dis. 2013, 45, 271–278. [Google Scholar] [CrossRef]
- Lahlaoui, H.; Khalifa, A.B.H.; Moussa, M.B. Epidemiology of Enterobacteriaceae producing CTX-M type extended spectrum β-lactamase (ESBL). Med. Mal. Infect. 2014, 44, 400–404. [Google Scholar] [CrossRef]
- Mojtahedi, A. The presence of extended-spectrum β-lactamase as a risk factor for MDR in clinical isolation of Escherichia coli. Trop. Biomed. 2017, 34, 98–109. [Google Scholar]
- Kapil, A. The challenge of antibiotic resistance: Need to contemplate. Indian J. Med. Res. 2005, 121, 83. [Google Scholar]
- Idrees, M.M.; Saeed, A. Genetic and molecular mechanisms of multidrug-resistance in uropathogens and novel therapeutic combat. In Biochemistry of Drug Resistance; Springer: Berlin/Heidelberg, Germany, 2021; pp. 505–538. [Google Scholar]
- Moo, C.-L.; Yang, S.-K.; Yusoff, K.; Ajat, M.; Thomas, W.; Abushelaibi, A.; Lim, S.-H.-E.; Lai, K.-S. Mechanisms of antimicrobial resistance (AMR) and alternative approaches to overcome AMR. Curr. Drug Discov. Technol. 2020, 17, 430–447. [Google Scholar] [CrossRef]
- Argimón, S.; Yeats, C.A.; Goater, R.J.; Abudahab, K.; Taylor, B.; Underwood, A.; Sánchez-Busó, L.; Wong, V.K.; Dyson, Z.A.; Nair, S. A global resource for genomic predictions of antimicrobial resistance and surveillance of Salmonella Typhi at pathogenwatch. Nat. Commun. 2021, 12, 2879. [Google Scholar] [CrossRef] [PubMed]
- Teklu, D.S.; Negeri, A.A.; Legese, M.H.; Bedada, T.L.; Woldemariam, H.K.; Tullu, K.D. Extended-spectrum beta-lactamase production and multi-drug resistance among Enterobacteriaceae isolated in Addis Ababa, Ethiopia. Antimicrob. Resist. Infect. Control 2019, 8, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, B.V.; Sridhar, V. Diversity Studies in Yellow Pericarp Sorghum [Sorghum bicolor (L.) Moench] Genotypes for Yield Attributes. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 361–366. [Google Scholar] [CrossRef]
- Ahmed, Y.M.A.; Mohammed El Imam, M.; Khalid, K.E.; Mohamed, B.A.; Elhag, K.M. Phenotype and genotype of extendedspectrum-beta-lactamases in Sudanese patients with UTI. Basic Res. J. Microbiol. 2014, 3, 1–5. [Google Scholar]
- Hijazi, S.; Fawzi, M.; Ali, F.; Abd El Galil, K. Prevalence and characterization of extended-spectrum beta-lactamases producing Enterobacteriaceae in healthy children and associated risk factors. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 3. [Google Scholar] [CrossRef] [Green Version]
- Salvia, T.; Dolma, K.G.; Dhakal, O.P.; Khandelwal, B.; Singh, L.S. Phenotypic Detection of ESBL, AmpC, MBL, and Their Co-occurrence among MDR Enterobacteriaceae Isolates in a Tertiary Care Hospital in Sikkim, India. J. Lab. Physicians 2022, 14, 329–335. [Google Scholar]
- Merah-Fergani, O.; Sebaihia, M.; Berrazeg, M.; Amraoui, R.; Diene, S.M.; Rolain, J.-M. Occurrence and Diversity of Extended-Spectrum β-lactamases in Clinical Isolates of Enterobacte-riaceae in A Tertiary Care Hospital in Algeria. J. Infect. Dis. Antimicrob. Agents 2022, 39, 1–14. [Google Scholar]
- Casellas, J.; Goldberg, M. Incidence of strains producing extended spectrum β-lactamases in Argentina. Infection 1989, 17, 434–436. [Google Scholar] [CrossRef]
- Aksaray, S.G.S.; DokuzoĞuz, B.; Güvener, E.; Yücesoy, M.; YuluĞ, N.; Kocagöz, S.; Ünal, S.; Çetin, S.; Çalangu, S.; Günaydın, M. Surveillance of antimicrobial resistance among gram-negative isolates from intensive care units in eight hospitals in Turkey. J. Antimicrob. Chemother. 2000, 45, 695–699. [Google Scholar] [CrossRef] [Green Version]
- Sid Ahmed, M.A.; Bansal, D.; Acharya, A.; Elmi, A.A.; Hamid, J.M.; Sid Ahmed, A.M.; Chandra, P.; Ibrahim, E.; Sultan, A.A.; Doiphode, S. Antimicrobial susceptibility and molecular epidemiology of extended-spectrum beta-lactamase-producing Enterobacteriaceae from intensive care units at Hamad Medical Corporation, Qatar. Antimicrob. Resist. Infect. Control 2016, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Ejaz, H.; Younas, S.; Abosalif, K.O.; Junaid, K.; Alzahrani, B.; Alsrhani, A.; Abdalla, A.E.; Ullah, M.I.; Qamar, M.U.; Hamam, S.S. Molecular analysis of bla SHV, bla TEM, and bla CTX-M in extended-spectrum β-lactamase producing Enterobacteriaceae recovered from fecal specimens of animals. PLoS ONE 2021, 16, e0245126. [Google Scholar] [CrossRef]
- Haider, M.H.; McHugh, T.D.; Roulston, K.; Arruda, L.B.; Sadouki, Z.; Riaz, S. Detection of carbapenemases blaOXA48-blaKPC-blaNDM-blaVIM and extended-spectrum-β-lactamase blaOXA1-blaSHV-blaTEM genes in Gram-negative bacterial isolates from ICU burns patients. Ann. Clin. Microbiol. Antimicrob. 2022, 21, 18. [Google Scholar] [CrossRef]
- Thomson, K.S.; Sanders, C.C. Detection of extended-spectrum beta-lactamases in members of the family Enterobacteriaceae: Comparison of the double-disk and three-dimensional tests. Antimicrob. Agents Chemother. 1992, 36, 1877–1882. [Google Scholar] [CrossRef] [Green Version]
- Bullens, M.; de Cerqueira Melo, A.; Raziq, S.; Lee, J.; Khalid, G.; Khan, S.; Zada, A.; Wailly, Y.; Zeshan, S.; Saad, N. Antibiotic resistance in patients with urinary tract infections in Pakistan. Public Health Action 2022, 12, 48–52. [Google Scholar] [CrossRef]
- Mazzariol, A.; Bazaj, A.; Cornaglia, G. Multi-drug-resistant Gram-negative bacteria causing urinary tract infections: A review. J. Chemother. 2017, 29 (Suppl. 1), 2–9. [Google Scholar] [CrossRef] [Green Version]
- Silver, L.L. Fosfomycin: Mechanism and resistance. Cold Spring Harb. Perspect. Med. 2017, 7, a025262. [Google Scholar] [CrossRef]
- Polse, R.F.; Yousif, S.Y.; Assafi, M.S. Prevalence and molecular characterization of extended spectrum beta-Lactamases-producing uropathogenic Escherichia coli isolated in Zakho, Iraq. J. Microbiol. Infect. Dis. 2016, 6, 163–167. [Google Scholar]
- Alqasim, A.; Abu Jaffal, A.; Alyousef, A.A. Prevalence of multidrug resistance and extended-spectrum β-lactamase carriage of clinical uropathogenic Escherichia coli isolates in Riyadh, Saudi Arabia. Int. J. Microbiol. 2018, 2018, 3026851. [Google Scholar] [CrossRef] [Green Version]
- Mashwal, F.A.; El Safi, S.H.; George, S.K.; Adam, A.A.; Jebakumar, A.Z. Incidence and molecular characterization of the extended spectrum beta lactamase-producing Escherichia coli isolated from urinary tract infections in Eastern Saudi Arabia. Saudi Med. J. 2017, 38, 811. [Google Scholar] [CrossRef]
- Jena, J.; Sahoo, R.K.; Debata, N.K.; Subudhi, E. Prevalence of TEM, SHV, and CTX-M genes of extended-spectrum β-lactamase-producing Escherichia coli strains isolated from urinary tract infections in adults. 3 Biotech 2017, 7, 244. [Google Scholar] [CrossRef]
- Ogefere, H.O.; Aigbiremwen, P.A.; Omoregie, R. Extended-spectrum beta-lactamase (ESBL)–producing Gram-negative isolates from urine and wound specimens in a tertiary health facility in southern Nigeria. Trop. J. Pharm. Res. 2015, 14, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Nwafuluaku, I.G.; Agbakoba, N.R.; Ushie, S.N.; Dilibe, E.A. Prevalence and molecular characterization of extended spectrum beta-lactamase producing uropathogenic Escherichia coli in a tertiary health care facility in Anambra State, Nigeria. World J. Adv. Res. Rev. 2021, 11, 300–311. [Google Scholar] [CrossRef]
- Cantón, R.; Coque, T.M. The CTX-M β-lactamase pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Bali, E.B.; Acik, L.; Sultan, N. Phenotypic and molecular characterization of SHV, TEM, CTX-M and extended-spectrum beta-lactamase produced by Escherichia coli, Acinobacter baumannii and Klebsiella isolates in a Turkish hospital. Afr. J. Microbiol. Res. 2010, 4, 650–654. [Google Scholar]
- Amoutzias, G.D.; Nikolaidis, M.; Hesketh, A. The Notable Achievements and the Prospects of Bacterial Pathogen Genomics. Microorganisms 2022, 10, 1040. [Google Scholar] [CrossRef]
- Rohner, P.; Pepey, B.; Auckenthaler, R. Advantage of combining resin with lytic BACTEC blood culture media. J. Clin. Microbiol. 1997, 35, 2634–2638. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Romero, M.I.; Moya, J.M.G.-L.; López, J.J.G.; Mira, N.O. Collection, transport and general processing of clinical specimens in Microbiology laboratory. Enferm. Infecc. Microbiol. Clin. 2019, 37, 127–134. [Google Scholar] [CrossRef]
- Cheesbrough, M. District Laboratory Practice in Tropical Countries, Part 2; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Coico, R. Gram staining. Curr. Protoc. Microbiol. 2006. [Google Scholar] [CrossRef]
- Holmes, B.; Willcox, W.; Lapage, S. Identification of Enterobacteriaceae by the API 20E system. J. Clin. Pathol. 1978, 31, 22–30. [Google Scholar] [CrossRef]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of changes to the clinical and laboratory standards institute performance standards for antimicrobial susceptibility testing, M100. J. Clin. Microbiol. 2021, 59, e0021321. [Google Scholar] [CrossRef]
- Jarlier, V.; Nicolas, M.-H.; Fournier, G.; Philippon, A. Extended broad-spectrum β-lactamases conferring transferable resistance to newer β-lactam agents in Enterobacteriaceae: Hospital prevalence and susceptibility patterns. Clin. Infect. Dis. 1988, 10, 867–878. [Google Scholar] [CrossRef]
- Drieux, L.; Brossier, F.; Sougakoff, W.; Jarlier, V. Phenotypic detection of extended-spectrum β-lactamase production in Enterobacteriaceae: Review and bench guide. Clin. Microbiol. Infect. 2008, 14, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.B.; Dablool, A. Quality improvement of the DNA extracted by boiling method in gram negative bacteria. Int. J. Bioassays 2017, 6, 5347–5349. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, J.; Navon-Venezia, S.; Chmelnitsky, I.; Hammer-Munz, O.; Leavitt, A.; Gold, H.S.; Schwaber, M.J.; Carmeli, Y. Extended-spectrum beta-lactamases among Enterobacter isolates obtained in Tel Aviv, Israel. Antimicrob. Agents Chemother. 2005, 49, 1150–1156. [Google Scholar] [CrossRef] [Green Version]
- Pallecchi, L.; Malossi, M.; Mantella, A.; Gotuzzo, E.; Trigoso, C.; Bartoloni, A.; Paradisi, F.; Kronvall, G.R.; Rossolini, G.M. Detection of CTX-M-type β-lactamase genes in fecal Escherichia coli isolates from healthy children in Bolivia and Peru. Antimicrob. Agents Chemother. 2004, 48, 4556–4561. [Google Scholar] [CrossRef] [Green Version]
- Colom, K.; Pérez, J.; Alonso, R.; Fernández-Aranguiz, A.; Lariño, E.; Cisterna, R. Simple and reliable multiplex PCR assay for detection of bla TEM, bla SHV and bla OXA–1 genes in Enterobacteriaceae. FEMS Microbiol. Lett. 2003, 223, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, M.P.; Lewis, J.S. The clinical and laboratory standards institute subcommittee on antimicrobial susceptibility testing: Background, organization, functions, and processes. J. Clin. Microbiol. 2020, 58, e01864-19. [Google Scholar] [CrossRef]


{kind=link}
| Age | 0–9 | 10–19 | 20–29 | 30–39 | 40–49 | 50–59 | 60–69 | >70 | |
|---|---|---|---|---|---|---|---|---|---|
| Sample Size | (n = 24) | (n = 27) | (n = 22) | (n = 30) | (n = 38) | (n = 55) | (n = 40) | (n = 36) | |
| % | 8.8 | 9.9 | 8.1 | 11.0 | 14.0 | 20.0 | 14.7 | 13.2 | |
| Gender | Male (n = 138) | 11 | 14 | 3 | 14 | 18 | 24 | 26 | 28 |
| 50.7% | (8.0) | (10.1) | (2.2) | (10.1) | (13.0) | (17.4) | (18.8) | (20.3) | |
| Female (n = 134) | 13 | 13 | 19 | 16 | 20 | 31 | 14 | 8 | |
| 49.3% | (9.7) | (9.7) | (14.2) | (11.9) | (14.9) | (23.1) | (10.4) | (6.0) | |
| Ward | OPD (n = 123) | 15 | 10 | 7 | 15 | 21 | 24 | 16 | 15 |
| 45.2% | (12.5) | (8.1) | (5.7) | (12.2) | (17.1) | (19.5) | (13.0) | (12.2) | |
| Nephrology (n = 58) | 0 | 2 | 9 | 9 | 8 | 13 | 8 | 9 | |
| 21.3% | (0.0) | (3.4) | (15.5) | (15.5) | (13.8) | (22.4) | (13.80 | (15.5) | |
| Emergency (n = 26) | 1 | 4 | 1 | 1 | 1 | 9 | 4 | 5 | |
| 9.6% | (3.8) | (15.4) | (3.8) | (3.8) | (3.8) | (34.6) | (15.4) | (19.2) | |
| ICU (n = 13) | 1 | 4 | 3 | 0 | 0 | 1 | 1 | 3 | |
| 4.8% | (7.7) | (30.8) | (23.1) | (0.0) | (0.0) | (7.7) | (7.7) | (23.1) | |
| Urology (n = 52) | 7 | 7 | 2 | 5 | 8 | 8 | 11 | 4 | |
| 19.1% | (13.5) | (13.5) | (3.8) | (9.6) | (15.4) | (15.4) | (21.2) | (7.7) | |
| Bacterial isolate | Escherichia coli (n = 174) | 14 | 14 | 11 | 20 | 28 | 40 | 22 | 25 |
| 64.0 | (8.0) | (8.0) | (6.3) | (11.5) | (16.1) | (23.0) | (12.6) | (14.4) | |
| Klebsiella spp. (n = 74) | 6 | 8 | 6 | 8 | 6 | 14 | 16 | 10 | |
| 27.2 | (8.1) | (10.8) | (8.1) | (10.8) | (8.1) | (18.9) | (21.6) | (13.5) | |
| Acinetobacter spp. (n = 18) | 4 | 3 | 4 | 2 | 4 | 1 | 0 | 0 | |
| 6.6 | (22.2) | (16.7) | (22.2) | (11.1) | (22.2) | (5.6) | (0.0) | (0.0) | |
| Others (n = 6) | 0 | 2 | 1 | 0 | 0 | 0 | 2 | 1 | |
| 2.2 | (0.0) | (33.3) | (16.7) | (0.0) | (0.0) | (0.0) | (33.3) | (16.7) | |
| Sample collection Source | Urine (n = 198) | 16 | 17 | 15 | 21 | 31 | 39 | 33 | 26 |
| 72.8% | (8.1) | (8.6) | (7.6) | (10.6) | (15.7) | (19.7) | (16.7) | (13.1) | |
| Blood (n = 17) | 1 | 4 | 1 | 1 | 3 | 4 | 1 | 2 | |
| 6.3% | (5.9) | (23.5) | (5.9) | (5.9) | (17.6) | (23.5) | (5.9) | (11.8) | |
| Pus (n = 16) | 0 | 1 | 2 | 1 | 1 | 6 | 2 | 3 | |
| 5.9% | (0.0) | (6.3) | (12.5) | (6.3) | (6.3) | (37.5) | (12.5) | (18.8) | |
| Throat aspiration (n = 13) | 4 | 2 | 1 | 1 | 1 | 1 | 1 | 2 | |
| 4.8% | (30.8) | (15.4) | (7.5) | (7.7) | (7.7) | (7.7) | (7.7) | (15.4) | |
| CVC Tip (n = 8) | 1 | 0 | 2 | 1 | 2 | 0 | 1 | 1 | |
| 2.9% | (12.5) | (0.0) | (25.0) | (12.5) | (25.0) | (0.0) | (12.5) | (12.5) | |
| Sputum (n = 5) | 0 | 1 | 0 | 1 | 0 | 2 | 0 | 1 | |
| 1.8% | (0.0) | (20.0) | (0.0) | (20.0) | (0.0) | (40.0) | (0.0) | (20.0) | |
| PCN (n = 4) | 0 | 0 | 0 | 2 | 0 | 1 | 1 | 0 | |
| 1.5% | (0.0) | (0.0) | (0.0) | (50.0) | (0.0) | (25.0) | (25.0) | (0.0) | |
| Bronchial aspiration (n = 4) | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | |
| 1.5% | (25.0) | (0.0) | (0.0) | (25.0) | (0.0) | (0.0) | (25.0) | (25.0) | |
| Others (n = 7) | 1 | 2 | 1 | 1 | 0 | 2 | 0 | 0 | |
| 2.6% | (14.3) | (28.6) | (14.3) | (14.3) | (0.0) | (28.6) | (0.0) | (0.0) | |
| Antibiotics | Escherichia coli n = 174 | Klebsiella Species n = 74 | Acinetobacter Species n = 18 | Others n = 6 | Total n = 272 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | |
| TZP | 45 | 1 | 128 | 43 | 1 | 30 | 17 | 0 | 1 | 2 | 0 | 4 | 107 | 2 | 163 |
| (25.9) | (0.6) | (73.6) | (58.1) | (1.4) | (40.5) | (94.4) | (0.0) | (5.6) | (33.3) | (0.0) | (66.7) | (39.3) | (0.7) | 3 (59.9) | |
| MEM | 33 | 1 | 140 | 46 | 0 | 28 | 17 | 0 | 1 | 2 | 0 | 4 | 98 | 1 | 173 |
| (19.0) | (0.6) | (80.5) | (62.2) | (0.0) | (37.8) | (94.4) | (0.0) | (5.6) | (33.3) | (0.0) | (66.7) | (36.0) | (0.4) | (63.6) | |
| IPM | 35 | 2 | 137 | 44 | 0 | 30 | 17 | 0 | 1 | 2 | 0 | 4 | 98 | 2 | 172 |
| (20.1) | (1.1) | (78.7) | (59.5) | (0.0) | (40.5) | (94.4) | (0.0) | (5.6) | (33.3) | (0.0) | (66.7) | (36.0) | (0.7) | 63.2) | |
| G | 63 | 1 | 110 | 45 | 0 | 29 | 15 | 0 | 3 | 3 | 0 | 3 | 126 | 1 | 145 |
| (36.2) | (0.6) | (63.2) | (60.8) | (0.0) | (39.2) | (83.3) | (0.0) | (16.7) | (50.0) | (0.0) | (50.0) | (46.3) | (0.4) | (53.3) | |
| AK | 19 | 5 | 150 | 31 | 3 | 40 | 15 | 0 | 3 | 2 | 0 | 4 | 67 | 8 | 197 |
| (10.9) | (2.9) | (86.2) | (41.9) | (4.1) | (54.1) | (83.3) | (0.0) | (16.7) | (33.3) | (0.0) | (66.7) | (24.6) | (2.9) | (72.4) | |
| NA | 156 | 3 | 15 | 64 | 0 | 10 | 17 | 0 | 1 | 5 | 0 | 1 | 242 | 3 | 27 |
| (89.7) | (1.7) | (8.6) | (86.5) | (0.0) | (13.5) | (94.4) | (0.0) | (5.6) | (83.3) | (0.0) | (16.7) | (89.0) | (1.1) | (9.9) | |
| NOR | 150 | 0 | 24 | 58 | 1 | 15 | 17 | 0 | 1 | 6 | 0 | 0 | 231 | 1 | 40 |
| (86.2) | (0.0) | (13.8) | (78.4) | (1.4) | (20.3) | (94.4) | (0.0) | (5.6) | (100.0) | (0.0) | (0.0) | (84.9) | (0.4) | (14.7) | |
| CIP | 141 | 0 | 33 | 61 | 0 | 13 | 17 | 0 | 1 | 5 | 0 | 1 | 234 | 0 | 48 |
| (81.0) | (0.0) | (19.0) | (82.4) | (0.0) | (17.6) | (94.4) | (0.0) | (5.6) | (83.3) | (0.0) | (16.7) | (82.4) | (0.0) | (17.6) | |
| SXT | 146 | 0 | 28 | 64 | 0 | 10 | 17 | 0 | 1 | 4 | 0 | 2 | 231 | 0 | 41 |
| (83.9) | (0.0) | (16.1) | (86.5) | (0.0) | (13.5) | (94.4) | (0.0) | (5.6) | (66.7) | (0.0) | (33.3) | (84.9) | (0.0) | (15.1) | |
| F | 30 | 3 | 141 | 45 | 2 | 27 | 15 | 0 | 3 | 3 | 0 | 3 | 93 | 5 | 174 |
| (17.2) | (1.7) | (81.0) | (60.8) | (2.7) | (36.5) | (83.3) | (0.0) | (16.7) | (50.0) | (0.0) | (50.0) | (34.2) | (1.8) | (64.0) | |
| SCF | 50 | 2 | 122 | 45 | 1 | 28 | 16 | 1 | 1 | 2 | 0 | 4 | 113 | 4 | 155 |
| (28.7) | (1.1) | (70.1) | (60.8) | (1.4) | (37.8) | (88.9) | (5.6) | (5.6) | (33.3) | (0.0) | (66.7) | (41.5) | (1.5) | (57.0) | |
| FOS | 8 | 0 | 166 | 28 | 4 | 42 | 15 | 1 | 2 | 3 | 0 | 3 | 54 | 5 | 213 |
| (4.6) | (0.0) | (95.4) | (37.8) | (5.40 | (56.8) | (83.3) | (5.6) | (11.1) | (50.0) | (0.0) | (50.0) | (19.9) | (1.8) | (78.3) | |
| Bacterial Isolate N = 272 | CTX-M n = 248 (91.2%) | OXA n = 107 (39.3%) | TEM n = 168 (61.8%) | SHV n = 48 (17.6%) | p-Value | |
|---|---|---|---|---|---|---|
| Escherichia coli | Positive | 160 (92.0) | 71 (40.8) | 105 (60.3) | 25 (14.4) | 0.00 * |
| Negative | 14 (8.0) | 103 (59.2) | 69 (39.7) | 149 (85.6) | ||
| Klebsiella species | Positive | 68 (91.9) | 30 (40.5) | 46 (62.2) | 18 (24.3) | 0.00 * |
| Negative | 6 (8.1) | 44 (59.5) | 28 (37.8) | 56 (75.7) | ||
| Acinetobacter species | Positive | 16 (88.9) | 5 (27.8) | 13 (72.2) | 4 (22.2) | 0.00 * |
| Negative | 2 (11.1) | 13 (72.2) | 5 (27.8) | 14 (77.8) | ||
| Others | Positive | 4 (66.7) | 1 (16.7) | 4 (66.7) | 1 (16.7) | 0.00 * |
| Negative | 2 (33.3) | 5 (83.7) | 2 (33.3) | 5 (83.7) | ||
| Genes | Total N (%) | Non-β-Lactam Antibiotics |
|---|---|---|
| blaCTX-M + blaTEM | 83 (30.5) | - |
| blaCTX-M | 46 (16.9) | - |
| blaCTX-M + blaOXA + blaTEM | 38 (14.0) | NA, NOR, CIP |
| blaCTX-M + blaOXA | 37 (13.6) | - |
| blaCTX-M + blaTEM + blaSHV | 19 (7.0) | SXT |
| blaCTX-M + blaTEM + blaOXA + blaSHV | 13 (4.8) | NA, NOR, CIP, SXT, |
| blaTEM | 7 (2.6) | NA, NOR, CIP, |
| blaOXA | 6 (2.2) | - |
| blaCTX-M + blaSHV | 6 (2.2) | NA, NOR, CIP, SXT, F, SCF, FOS |
| blaCTX-M + blaOXA + blaSHV | 6 (2.2) | NA, NOR, CIP, SXT, F |
| blaOXA + blaTEM | 5 (1.8) | NA, NOR, CIP, SXT |
| blaOXA + blaTEM + blaSHV | 2 (0.7) | MEM, IPM, G, AK, NA, NOR, CIP, SXT, SCF, F, FOS |
| Nil | 2 (0.7) | NA, NOR, CIP |
| blaSHV | 1 (0.4) | MEM, IPM, NA, NOR, CIP, SXT, SCF |
| blaTEM + blaSHV | 1 (0.4) | MEM, IPM, G, AK, NA, NOR, CIP, SXT, SCF, F, FOS |
| blaOXA + blaSHV | 0 | - |
| Target Gene | Primer Sequence | Product Size | Annealing Temperature (°C) | References | |
|---|---|---|---|---|---|
| blaTEM | F | 5′-TCAACATTTCCGTGTCG-3′ | 860 | 56 | [56] |
| R | 5′-CTGACAGTTACCAATGCTTA-3′ | ||||
| blaCTX-M | F | 5′-ATGTGCAGYACCAGTAARGT-3′ | 593 | 50 | [57] |
| R | 5′-TGGGTRAARTARGTSACCAGA-3′ | ||||
| blaOXA | F | 5′-ATATCTCTACTGTTGCATCTCC-3′ | 619 | 56 | [58] |
| R | 5′-AAACCCTTCAAACCATCC-3′ | ||||
| blaSHV | F | 5′-AGGATTGACTGCCTTTTTG-3′ | 392 | 56 | [58] |
| R | 5′-ATTTGCTGATTTCGCTCG-3′ | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Idrees, M.M.; Rimsha, R.; Idrees, M.D.; Saeed, A. Antimicrobial Susceptibility and Genetic Prevalence of Extended-Spectrum β-Lactamases in Gram-Negative Rods Isolated from Clinical Specimens in Pakistan. Antibiotics 2023, 12, 29. https://doi.org/10.3390/antibiotics12010029
Idrees MM, Rimsha R, Idrees MD, Saeed A. Antimicrobial Susceptibility and Genetic Prevalence of Extended-Spectrum β-Lactamases in Gram-Negative Rods Isolated from Clinical Specimens in Pakistan. Antibiotics. 2023; 12(1):29. https://doi.org/10.3390/antibiotics12010029
Chicago/Turabian StyleIdrees, Muhammad Mubashar, Rimsha Rimsha, Muhammad Daoud Idrees, and Ali Saeed. 2023. "Antimicrobial Susceptibility and Genetic Prevalence of Extended-Spectrum β-Lactamases in Gram-Negative Rods Isolated from Clinical Specimens in Pakistan" Antibiotics 12, no. 1: 29. https://doi.org/10.3390/antibiotics12010029