
Curcumin Stimulates the Overexpression of Virulence Factors in Salmonella enterica Serovar Typhimurium: In Vitro and Animal Model Studies
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
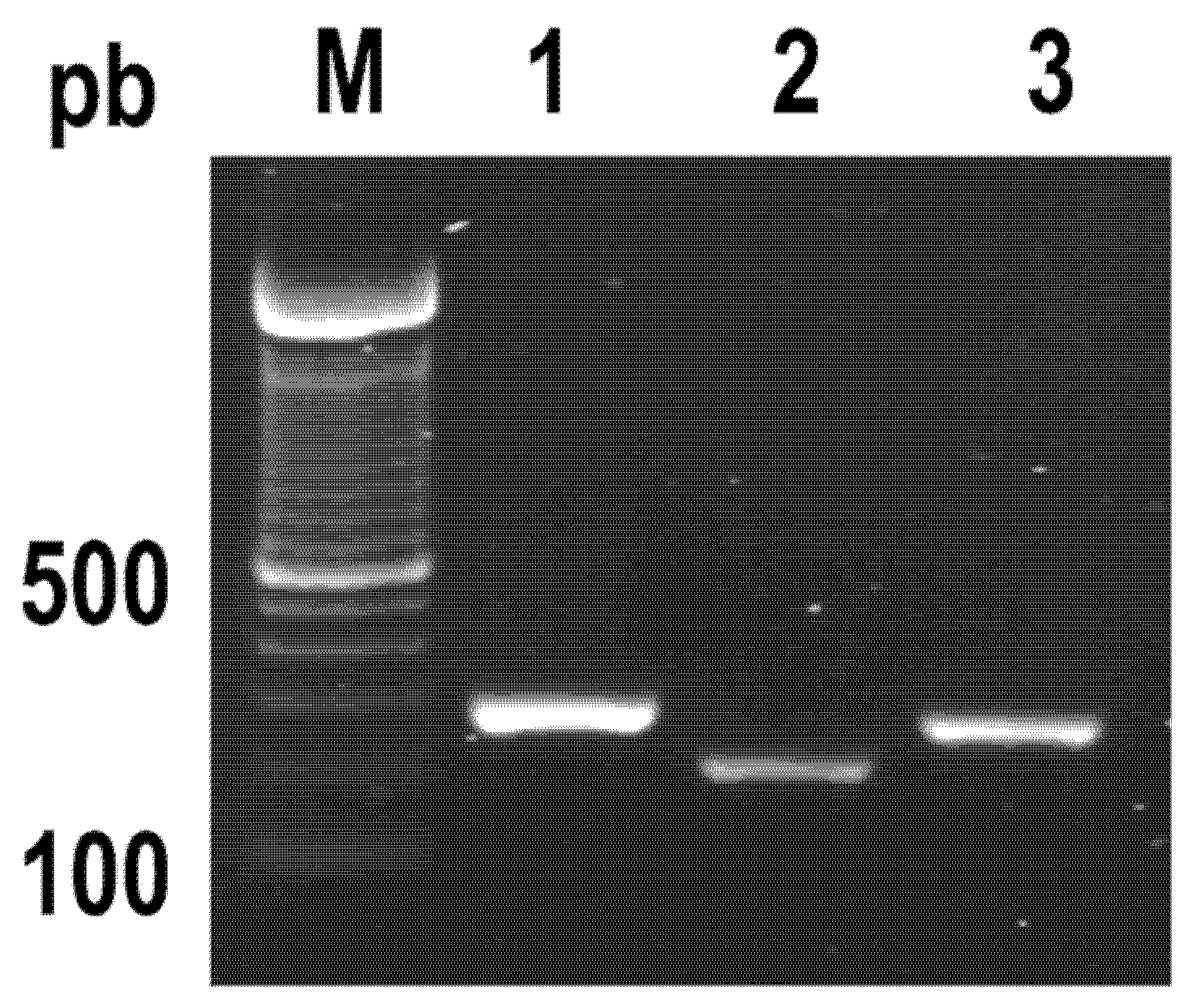
2.1. PCR Identification of Genes Encoding Specific Virulence Factors of S. Typhimurium
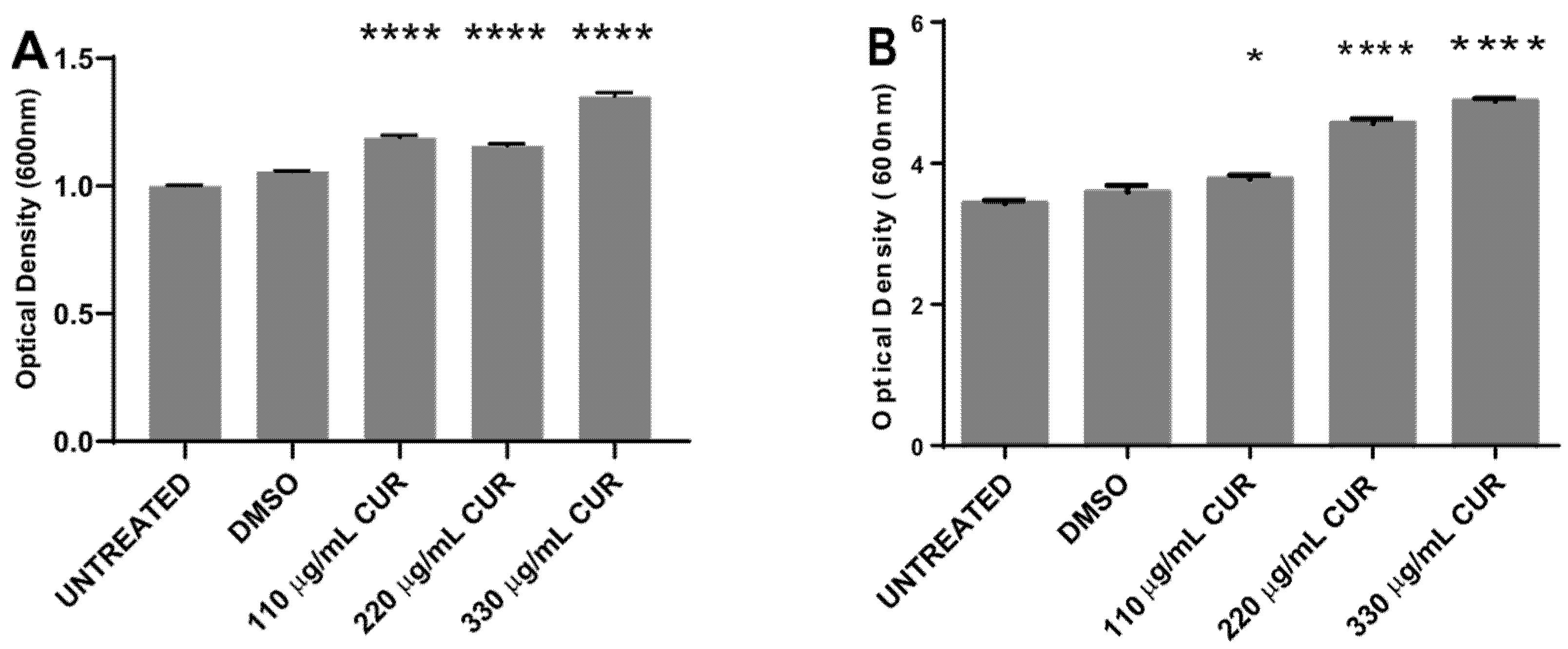
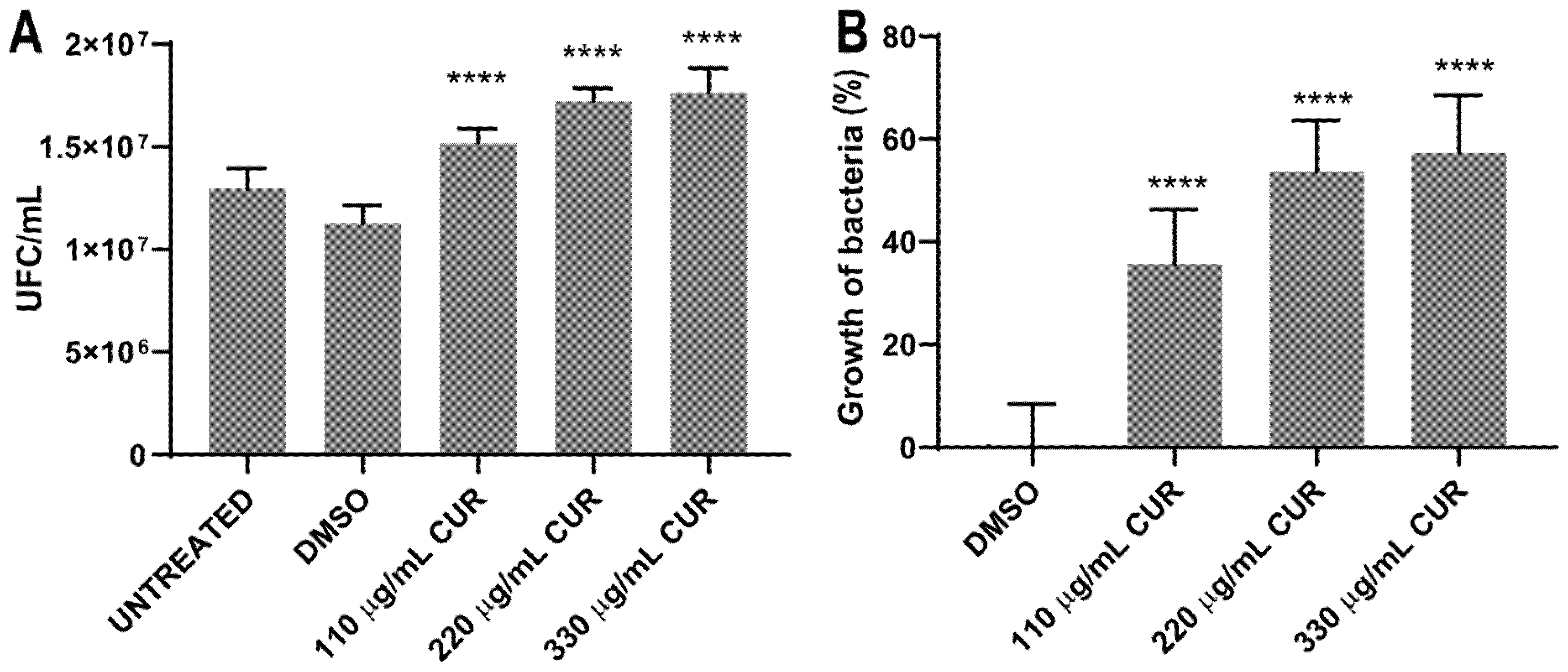
2.2. Curcumin Did Not Show an Antibacterial Effect
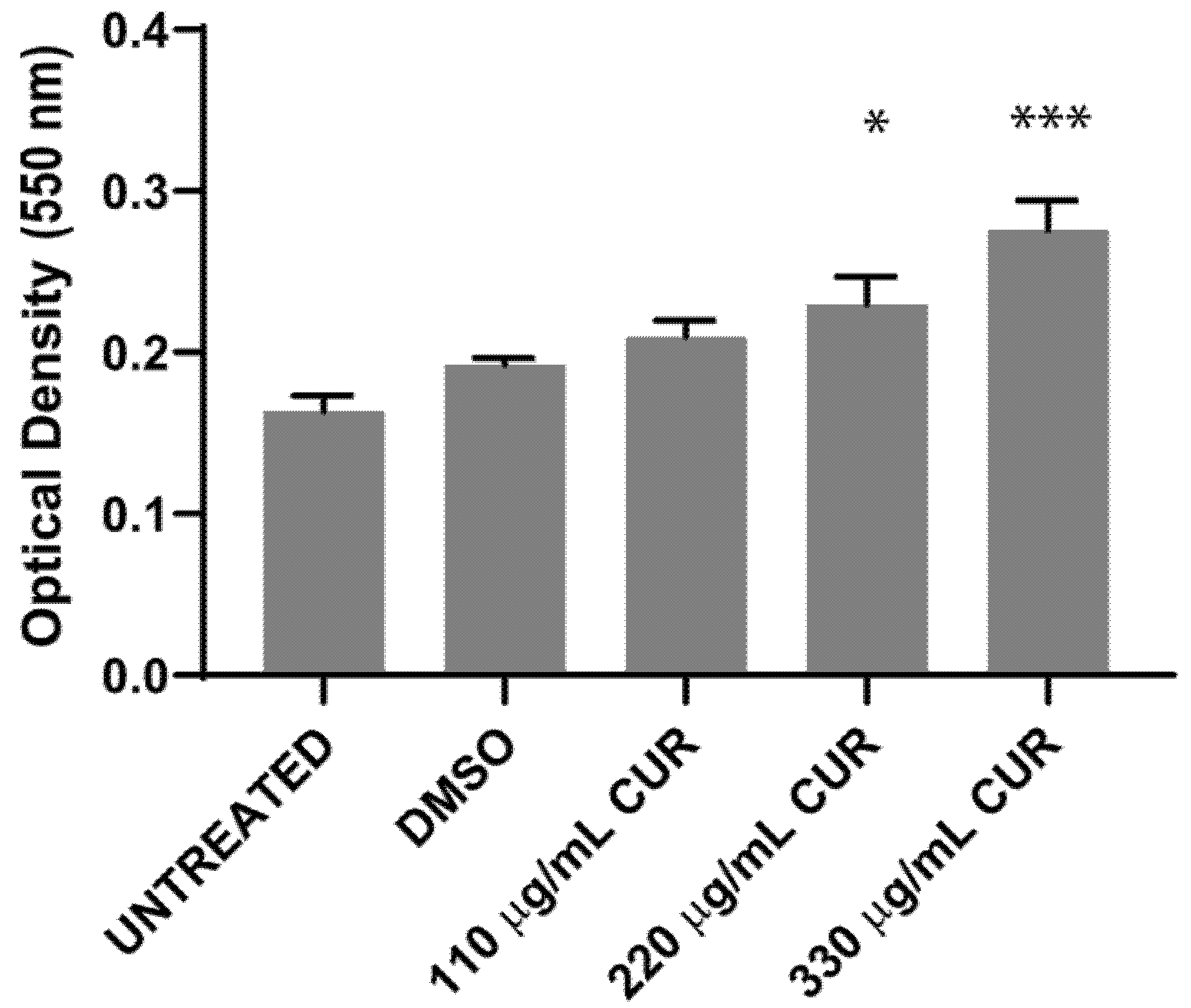
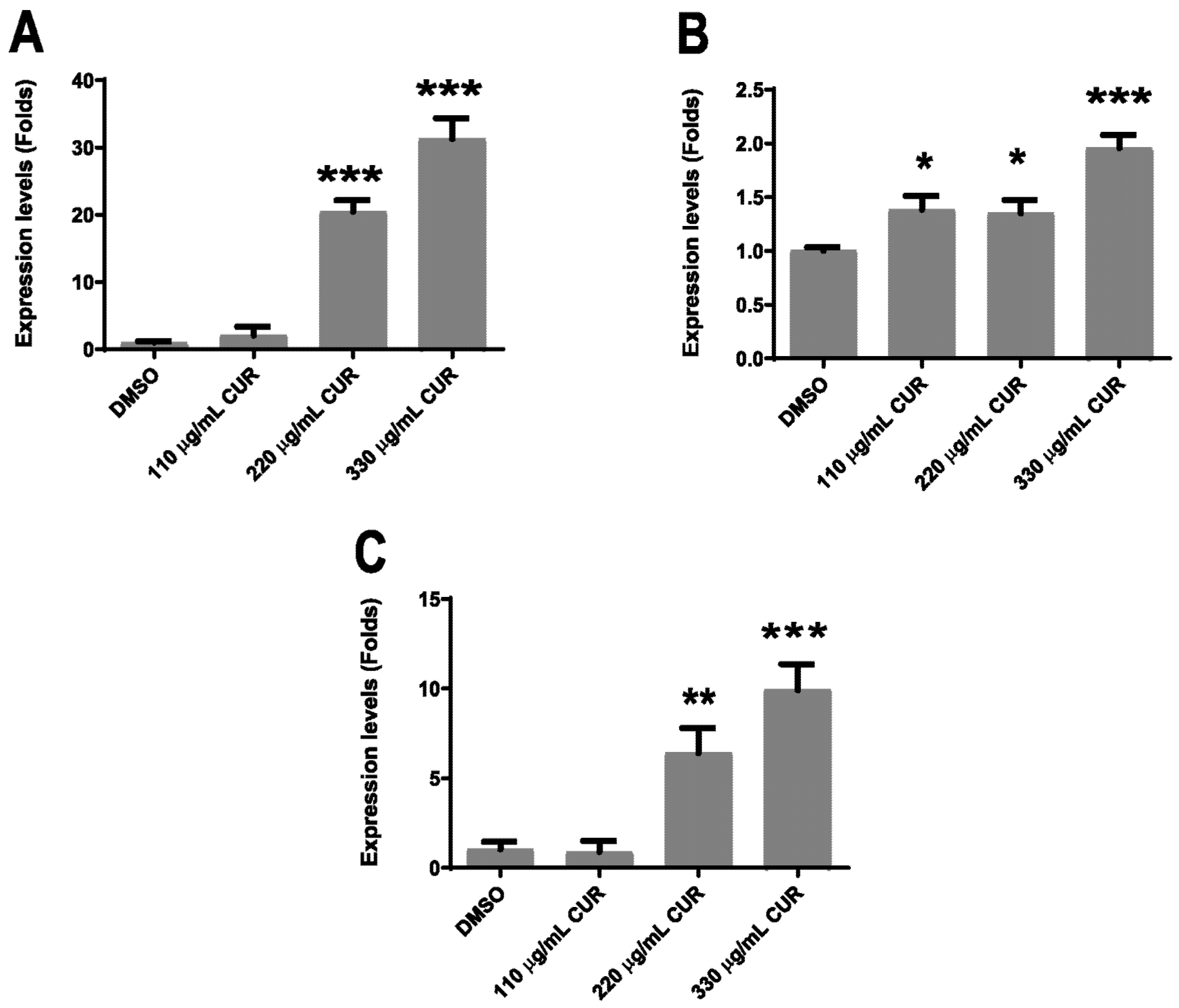
2.3. Virulence Factors Are Upregulated by Curcumin
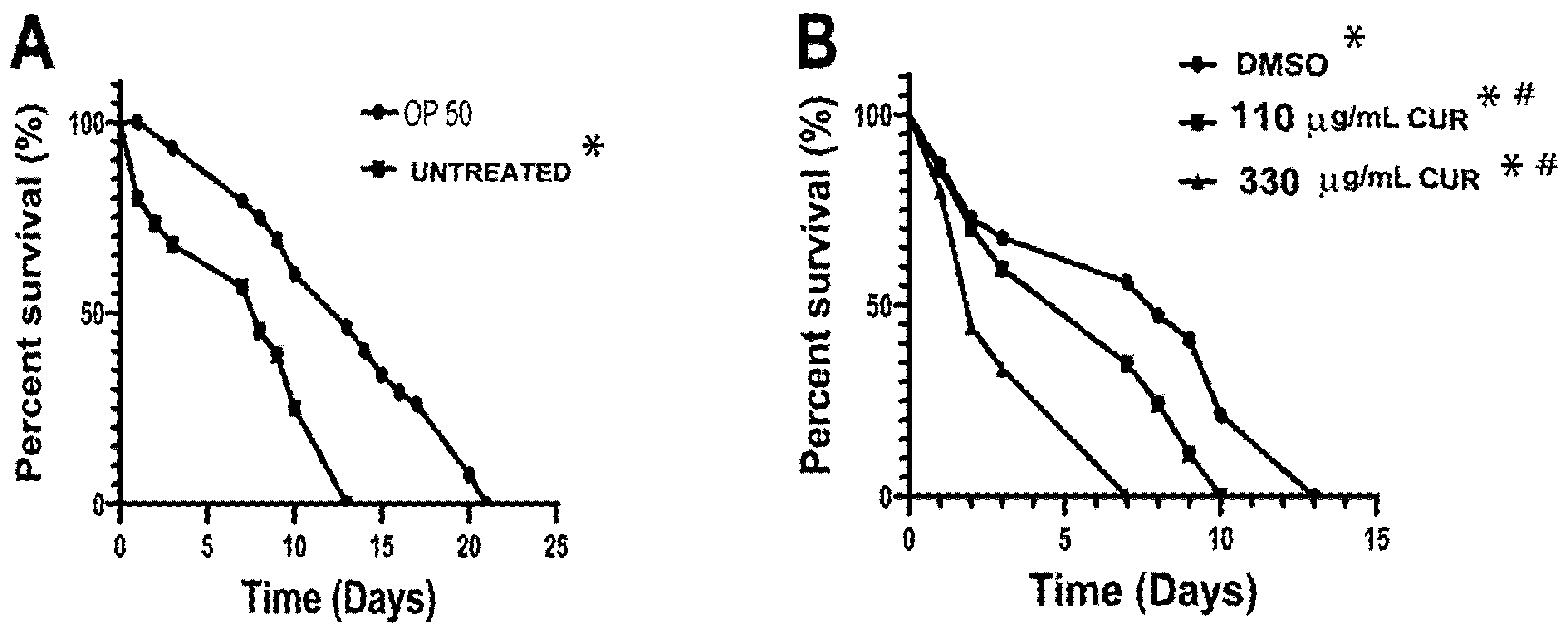
2.4. Curcumin Enhanced the Pathogenicity of S. Typhimurium in C. elegans
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain
4.2. Maintenance and Preservation of Microorganisms
4.3. Extraction of Genomic DNA
4.4. Presence of Virulence Genes
4.5. Preparation of Curcumin Stocks
4.6. Determination of the Antibacterial Activity of Curcumin
4.6.1. Spectrophotometric Method
4.6.2. Pour Plate Method
4.6.3. Assay MTT
4.6.4. Statistical Analysis
4.7. Relative-Quantitative RT-PCR
4.8. Maintenance and Preservation of C. elegans
4.9. Pathogenicity Assays of Salmonella Strains on the C. elegans Model
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Troeger, C.; Blacker, B.F.; Khalil, I.A.; Rao, P.C.; Cao, S.; Zimsen, S.R.M.; Albertson, S.B.; Stanaway, J.D.; Deshpande, A.; Abebe, Z.; et al. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 countries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed]
- Rabsch, W.; Andrews, H.L.; Kingsley, R.A.; Prager, R.; Tschape, H.; Adams, L.G.; Baumler, A.J. Salmonella enterica serotype Typhimurium and its host-adapted variants. Infect. Immun. 2002, 70, 2249–2255. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Msefula, C.L.; Thomson, N.R.; Kariuki, S.; Holt, K.E.; Gordon, M.A.; Harris, D.; Clarke, L.; Whitehead, S.; Sangal, V.; et al. Epidemic multiple drug resistant Salmonella Typhimurium causing invasive disease in sub-Saharan Africa have a distinct genotype. Genome Res. 2009, 19, 2279–2287. [Google Scholar] [CrossRef]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Imke Hansen-Wester, M.H. Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 2001, 3, 549–559. [Google Scholar] [CrossRef]
- Dos Santos, A.M.P.; Ferrari, R.G.; Conte-Junior, C.A. Virulence Factors in Salmonella Typhimurium: The Sagacity of a Bacterium. Curr. Microbiol. 2019, 76, 762–773. [Google Scholar] [CrossRef]
- Elhadad, D.; Desai, P.; Rahav, G.; McClelland, M.; Gal-Mor, O. Flagellin Is Required for Host Cell Invasion and Normal Salmonella Pathogenicity Island 1 Expression by Salmonella enterica Serovar Paratyphi A. Infect. Immun. 2015, 83, 3355–3368. [Google Scholar] [CrossRef]
- Ikeda, J.S.; Schmitt, C.K.; Darnell, S.C.; Watson, P.R.; Bispham, J.; Wallis, T.S.; Weinstein, D.L.; Metcalf, E.S.; Adams, P.; O’Connor, C.D.; et al. Flagellar phase variation of Salmonella enterica serovar Typhimurium contributes to virulence in the murine typhoid infection model but does not influence Salmonella-induced enteropathogenesis. Infect. Immun. 2001, 69, 3021–3030. [Google Scholar] [CrossRef]
- Gerlach, R.G.; Jackel, D.; Stecher, B.; Wagner, C.; Lupas, A.; Hardt, W.D.; Hensel, M. Salmonella Pathogenicity Island 4 encodes a giant non-fimbrial adhesin and the cognate type 1 secretion system. Cell Microbiol. 2007, 9, 1834–1850. [Google Scholar] [CrossRef]
- De, R.; Kundu, P.; Swarnakar, S.; Ramamurthy, T.; Chowdhury, A.; Nair, G.B.; Mukhopadhyay, A.K. Antimicrobial activity of curcumin against Helicobacter pylori isolates from India and during infections in mice. Antimicrob. Agents Chemother. 2009, 53, 1592–1597. [Google Scholar] [CrossRef] [PubMed]
- Niamsa, N.; Sittiwet, C. Antibacterial Activity of Curcuma longa Aqueous extract. J. Pharmacol. Toxicol. 2009, 4, 173–177. [Google Scholar] [CrossRef]
- Basniwal, R.K.; Buttar, H.S.; Jain, V.K.; Jain, N. Curcumin nanoparticles: Preparation, characterization, and antimicrobial study. J. Agric. Food Chem. 2011, 59, 2056–2061. [Google Scholar] [CrossRef]
- Liu, C.H.; Huang, H.Y. Antimicrobial Activity of Curcumin-Loaded Myristic Acid Microemulsions against Staphylococcus epidermidis. Chem. Pharm. Bull. 2012, 60, 1118–1124. [Google Scholar] [CrossRef]
- Liu, C.H.; Huang, H.Y. In Vitro Anti-Propionibacterium Activity by Curcumin Containing Vesicle System. Chem. Pharm. Bull. 2013, 61, 419–425. [Google Scholar] [CrossRef]
- Hu, P.; Huang, P.; Chen, M.W. Curcumin reduces Streptococcus mutans biofilm formation by inhibiting sortase A activity. Arch. Oral Biol. 2013, 58, 1343–1348. [Google Scholar] [CrossRef]
- Infante, K.; Chowdhury, R.; Nimmanapalli, R.; Reddy, G. Antimicrobial Activity of Curcumin Against Food-Borne Pathogens. VRI Bio. Med. Chem. 2014, 2, 12. [Google Scholar] [CrossRef]
- Tyagi, P.; Singh, M.; Kumari, H.; Kumari, A.; Mukhopadhyay, K. Bactericidal activity of curcumin I is associated with damaging of bacterial membrane. PLoS ONE 2015, 10, e0121313. [Google Scholar] [CrossRef]
- Gunes, H.; Gulen, D.; Mutlu, R.; Gumus, A.; Tas, T.; Topkaya, A.E. Antibacterial effects of curcumin: An in vitro minimum inhibitory concentration study. Toxicol. Ind. Health 2016, 32, 246–250. [Google Scholar] [CrossRef]
- Teow, S.Y.; Liew, K.; Ali, S.A.; Khoo, A.S.; Peh, S.C. Antibacterial Action of Curcumin against Staphylococcus aureus: A Brief Review. J. Trop. Med. 2016, 2016, 2853045. [Google Scholar] [CrossRef] [Green Version]
- Yun, D.G.; Lee, D.G. Antibacterial activity of curcumin via apoptosis-like response in Escherichia coli. Appl. Microbiol. Biotechnol. 2016, 100, 5505–5514. [Google Scholar] [CrossRef]
- Adamczak, A.; Ozarowski, M.; Karpinski, T.M. Curcumin, a Natural Antimicrobial Agent with Strain-Specific Activity. Pharmaceuticals 2020, 13, 153. [Google Scholar] [CrossRef]
- Marathe, S.A.; Ray, S.; Chakravortty, D. Curcumin increases the pathogenicity of Salmonella enterica serovar Typhimurium in murine model. PLoS ONE 2010, 5, e11511. [Google Scholar] [CrossRef] [PubMed]
- Sifri, C.D.; Begun, J.; Ausubel, F.M. The worm has turned—Microbial virulence modeled in Caenorhabditis elegans. Trends Microbiol. 2005, 13, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Couillault, C.; Ewbank, J.J. Diverse bacteria are pathogens of Caenorhabditis elegans. Infect. Immun. 2002, 70, 4705–4707. [Google Scholar] [CrossRef] [PubMed]
- Winfield, M.D.; Groisman, E.A. Role of nonhost environments in the lifestyles of Salmonella and Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 3687–3694. [Google Scholar] [CrossRef]
- Rangel-Castaneda, I.A.; Carranza-Rosales, P.; Guzman-Delgado, N.E.; Hernandez-Hernandez, J.M.; Gonzalez-Pozos, S.; Perez-Rangel, A.; Castillo-Romero, A. Curcumin Attenuates the Pathogenicity of Entamoeba histolytica by Regulating the Expression of Virulence Factors in an Ex-Vivo Model Infection. Pathogens 2019, 8, 127. [Google Scholar] [CrossRef]
- Rudrappa, T.; Bais, H.P. Curcumin, a known phenolic from Curcuma longa, attenuates the virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models. J. Agric. Food Chem. 2008, 56, 1955–1962. [Google Scholar] [CrossRef]
- Powell, J.R.; Ausubel, F.M. Models of Caenorhabditis elegans Infection by Bacterial and Fungal Pathogens. In Innate Immunity; Ewbank, J., Vivier, E., Eds.; Humana Press: Totowa, NJ, USA, 2008; pp. 403–427. [Google Scholar]
- O’Ryan, M.; Prado, V.; Pickering, L.K. A millennium update on pediatric diarrheal illness in the developing world. Semin. Pediatr. Infect. Dis. 2005, 16, 125–136. [Google Scholar] [CrossRef]
- Bada-Alambedji, R.; Fofana, A.; Seydi, M.; Akakpo, A.J. Antimicrobial resistance of Salmonella isolated from poultry carcasses in Dakar (Senegal). Braz. J. Microbiol. 2006, 37, 510–515. [Google Scholar] [CrossRef] [Green Version]
- Dodds, D.R. Antibiotic resistance: A current epilogue. Biochem. Pharmacol. 2017, 134, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Theuretzbacher, U. Global antimicrobial resistance in Gram-negative pathogens and clinical need. Curr. Opin. Microbiol. 2017, 39, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Berndtson, A.E. Increasing Globalization and the Movement of Antimicrobial Resistance between Countries. Surg. Infect. 2020, 21, 579–585. [Google Scholar] [CrossRef]
- Sasidharan, N.K.; Sreekala, S.R.; Jacob, J.; Nambisan, B. In vitro synergistic effect of curcumin in combination with third generation cephalosporins against bacteria associated with infectious diarrhea. Biomed. Res. Int. 2014, 2014, 561456. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Diaz, A.A.; Hernandez-Patlan, D.; Solis-Cruz, B.; Adhikari, B.; Kwon, Y.M.; Latorre, J.D.; Hernandez-Velasco, X.; Fuente-Martinez, B.; Hargis, B.M.; Lopez-Arellano, R.; et al. Evaluation of curcumin and copper acetate against Salmonella Typhimurium infection, intestinal permeability, and cecal microbiota composition in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Udekwu, K.I.; Parrish, N.; Ankomah, P.; Baquero, F.; Levin, B.R. Functional relationship between bacterial cell density and the efficacy of antibiotics. J. Antimicrob. Chemother. 2009, 63, 745–757. [Google Scholar] [CrossRef]
- Bulitta, J.B.; Yang, J.C.; Yohonn, L.; Ly, N.S.; Brown, S.V.; D’Hondt, R.E.; Jusko, W.J.; Forrest, A.; Tsuji, B.T. Attenuation of colistin bactericidal activity by high inoculum of Pseudomonas aeruginosa characterized by a new mechanism-based population pharmacodynamic model. Antimicrob. Agents Chemother. 2010, 54, 2051–2062. [Google Scholar] [CrossRef]
- Li, Z.; Vaziri, H. Treatment of chronic diarrhoea. Best Pract. Res. Clin. Gastroenterol. 2012, 26, 677–687. [Google Scholar] [CrossRef]
- Michael, R.W.; Brown, P.J.C.; Peter, G. Influence of Growth Rate on Susceptibility to Antimicrobial Agents: Modification of the Cell Envelope and Batch and Continuous Culture Studies. Antimicrob. Agents Chemother. 1990, 34, 1623–1628. [Google Scholar]
- Meysman, P.; Sanchez-Rodriguez, A.; Fu, Q.; Marchal, K.; Engelen, K. Expression divergence between Escherichia coli and Salmonella enterica serovar Typhimurium reflects their lifestyles. Mol. Biol. Evol. 2013, 30, 1302–1314. [Google Scholar] [CrossRef]
- Sargo, C.R.; Campani, G.; Silva, G.G.; Giordano, R.C.; Da Silva, A.J.; Zangirolami, T.C.; Correia, D.M.; Ferreira, E.C.; Rocha, I. Salmonella typhimurium and Escherichia coli dissimilarity: Closely related bacteria with distinct metabolic profiles. Biotechnol. Prog. 2015, 31, 1217–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, D.; Devi Negi, V. Stress-induced adaptations in Salmonella: A ground for shaping its pathogenesis. Microbiol. Res. 2019, 229, 126311. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cheng, H.; Wang, F.; Wei, D.; Wang, X. An improved 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reduction assay for evaluating the viability of Escherichia coli cells. J. Microbiol. Methods 2010, 82, 330–333. [Google Scholar] [CrossRef]
- Wilson, E.; Okuom, M.; Kyes, N.; Mayfield, D.; Wilson, C.; Sabatka, D.; Sandoval, J.; Foote, J.R.; Kangas, M.J.; Holmes, A.E.; et al. Using Fluorescence Intensity of Enhanced Green Fluorescent Protein to Quantify Pseudomonas aeruginosa. Chemosensors 2018, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Grela, E.; Kozlowska, J.; Grabowiecka, A. Current methodology of MTT assay in bacteria—A review. Acta Histochem. 2018, 120, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Shishodia, S.; Singh, T.; Chaturvedi, M.M. Modulation of transcription factors by curcumin. Adv. Exp. Med. Biol. 2007, 595, 127–148. [Google Scholar] [PubMed]
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Pinaud, L.; Sansonetti, P.J.; Phalipon, A. Host Cell Targeting by Enteropathogenic Bacteria T3SS Effectors. Trends Microbiol. 2018, 26, 266–283. [Google Scholar] [CrossRef]
- Jiao, Y.; Wilkinson, J.T.; Christine Pietsch, E.; Buss, J.L.; Wang, W.; Planalp, R.; Torti, F.M.; Torti, S.V. Iron chelation in the biological activity of curcumin. Free Radic. Biol. Med. 2006, 40, 1152–1160. [Google Scholar] [CrossRef]
- Minear, S.; O’Donnell, A.F.; Ballew, A.; Giaever, G.; Nislow, C.; Stearns, T.; Cyert, M.S. Curcumin inhibits growth of Saccharomyces cerevisiae through iron chelation. Eukaryot. Cell 2011, 10, 1574–1581. [Google Scholar] [CrossRef]
- Sem, X.; Rhen, M. Pathogenicity of Salmonella enterica in Caenorhabditis elegans relies on disseminated oxidative stress in the infected host. PLoS ONE 2012, 7, e45417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera-Diaz, E.; Barbosa-Cardenas, C.M.; Perez-Montano, J.A.; Gonzalez-Aguilar, D.; Pacheco-Gallardo, C.; Barba, J. Occurrence, serotype diversity, and antimicrobial resistance of salmonella in ground beef at retail stores in Jalisco state, Mexico. J. Food Prot. 2013, 76, 2004–2010. [Google Scholar] [CrossRef] [PubMed]
- Galan, J.E.; Ginocchio, C.; Costeas, P. Molecular and functional characterization of the Salmonella invasion gene invA: Homology of InvA to members of a new protein family. J. Bacteriol. 1992, 174, 4338–4349. [Google Scholar] [CrossRef] [PubMed]
- Latasa, C.; Roux, A.; Toledo-Arana, A.; Ghigo, J.M.; Gamazo, C.; Penades, J.R.; Lasa, I. BapA, a large secreted protein required for biofilm formation and host colonization of Salmonella enterica serovar Enteritidis. Mol. Microbiol. 2005, 58, 1322–1339. [Google Scholar] [CrossRef] [PubMed]
- Itzia Azucena, R.C.; Jose Roberto, C.L.; Martin, Z.R.; Rafael, C.Z.; Leonardo, H.H.; Gabriela, T.P.; Araceli, C.R. Drug Susceptibility Testing and Synergistic Antibacterial Activity of Curcumin with Antibiotics against Enterotoxigenic Escherichia coli. Antibiotics 2019, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. elegans; Caenorhabditis Genetics Center: Minneapolis, MN, USA, 2006; Volume 11. [Google Scholar]
- Brenner, S. The Genetics of Caenorhabditis Elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Hope, I.A. C. elegans: A Practical Approach, 1st ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 1999; Volume 213, p. 304. [Google Scholar]
- Amrit, F.R.; Ratnappan, R.; Keith, S.A.; Ghazi, A. The C. elegans lifespan assay toolkit. Methods 2014, 68, 465–475. [Google Scholar] [CrossRef]
- Park, H.H.; Jung, Y.; Lee, S.V. Survival assays using Caenorhabditis elegans. Mol. Cells 2017, 40, 90–99. [Google Scholar] [CrossRef]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | A Average Number of Colonies Per Milliliter with Dilution 104 |
|---|---|
| S. Typhimurium | |
| Untreated | 1.30 × 107 CFU/mL |
| DMSO | 1.13 × 107 CFU/mL |
| CUR 110 μg/mL | 1.52 × 107 CFU/mL |
| CUR 220 μg/mL | 1.72 × 107 CFU/mL |
| CUR 330 μg/mL | 1.77 × 107 CFU/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zermeño-Ruiz, M.; Rangel-Castañeda, I.A.; Suárez-Rico, D.O.; Hernández-Hernández, L.; Cortés-Zárate, R.; Hernández-Hernández, J.M.; Camargo-Hernández, G.; Castillo-Romero, A. Curcumin Stimulates the Overexpression of Virulence Factors in Salmonella enterica Serovar Typhimurium: In Vitro and Animal Model Studies. Antibiotics 2022, 11, 1230. https://doi.org/10.3390/antibiotics11091230
Zermeño-Ruiz M, Rangel-Castañeda IA, Suárez-Rico DO, Hernández-Hernández L, Cortés-Zárate R, Hernández-Hernández JM, Camargo-Hernández G, Castillo-Romero A. Curcumin Stimulates the Overexpression of Virulence Factors in Salmonella enterica Serovar Typhimurium: In Vitro and Animal Model Studies. Antibiotics. 2022; 11(9):1230. https://doi.org/10.3390/antibiotics11091230
Chicago/Turabian StyleZermeño-Ruiz, Martin, Itzia A. Rangel-Castañeda, Daniel Osmar Suárez-Rico, Leonardo Hernández-Hernández, Rafael Cortés-Zárate, José M. Hernández-Hernández, Gabriela Camargo-Hernández, and Araceli Castillo-Romero. 2022. "Curcumin Stimulates the Overexpression of Virulence Factors in Salmonella enterica Serovar Typhimurium: In Vitro and Animal Model Studies" Antibiotics 11, no. 9: 1230. https://doi.org/10.3390/antibiotics11091230