

Combination Treatment for Inhibition of the Growth of Staphylococcus aureus with Recombinant SAP8 Endolysin and Nisin



Abstract
:1. Introduction
2. Materials and Methods
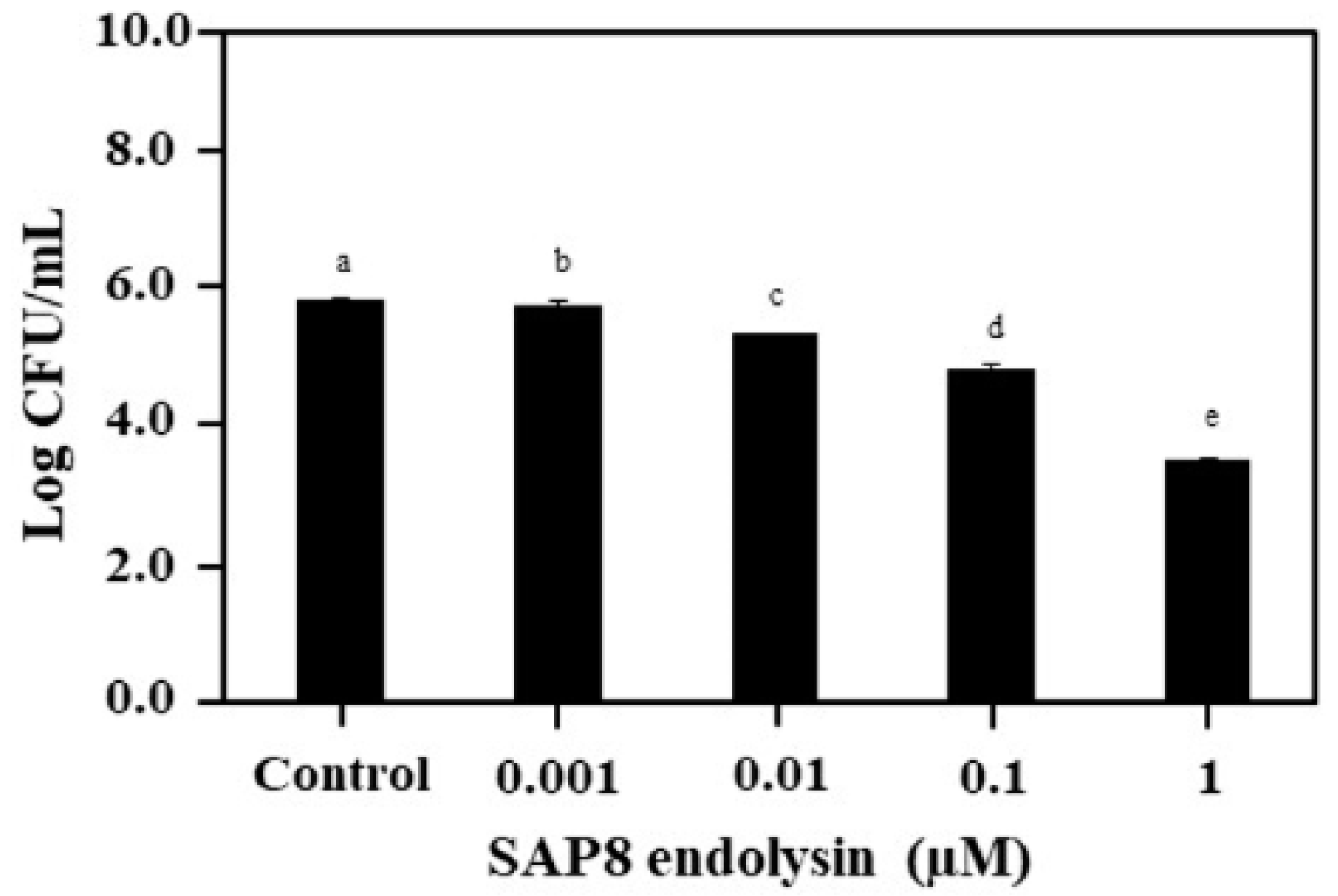
2.1. Antimicrobial Activity of the Recombinant SAP8 Endolysin against Staphylococcus aureus
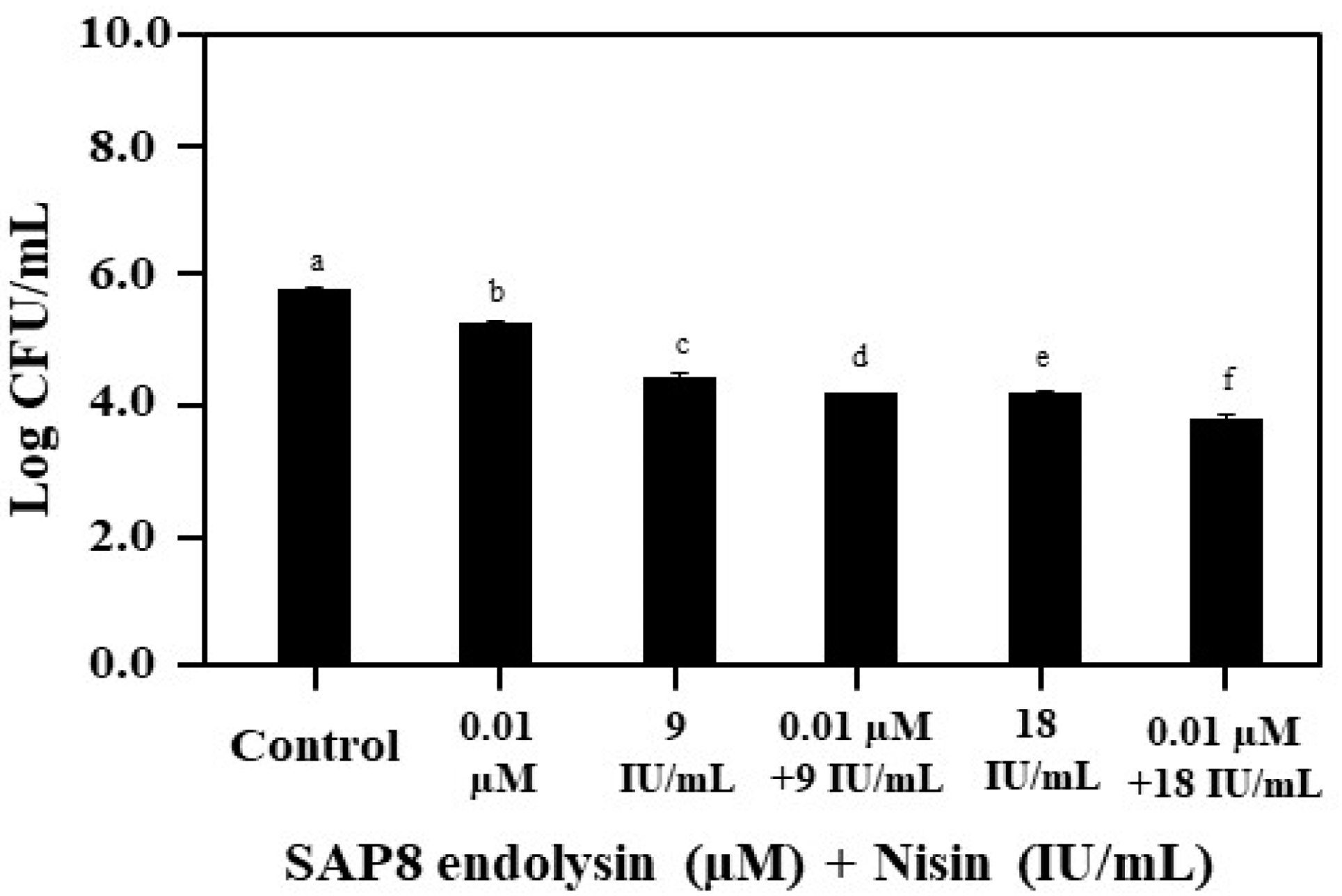
2.2. Co-Treatment with the Recombinant SAP8 Endolysin and Nisin against Staphylococcus aureus
2.3. Scanning Electron Microscopy of S. aureus Co-Treated with Endolysin and Nisin
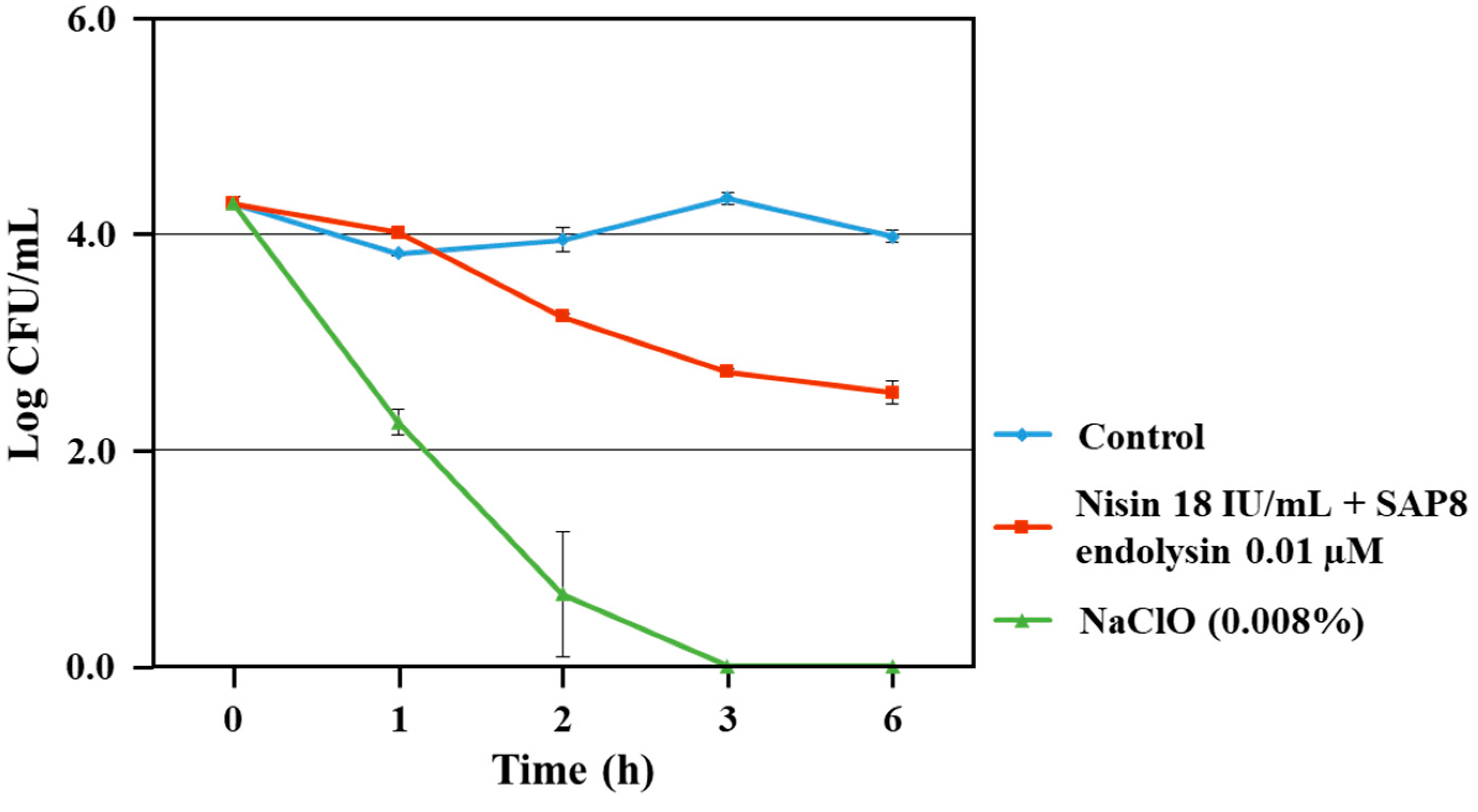
2.4. Fresh Agricultural Food (Lettuce) Application Test
3. Results
3.1. Growth Inhibition of S. aureus KCTC 3881 with Recombinant SAP8 Endolysin
3.2. Growth Inhibition of S. aureus KCTC 3881 Followed by Co-Treatment with Recombinant SAP8 Endolysin and Nisin
3.3. Growth Inhibition of S. aureus KCTC 3881 on Lettuce
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, W.; Yuk, H.G. Antimicrobial Efficacy of Syzygium Antisepticum Plant Extract against Staphylococcus aureus and Methicillin-Resistant S. aureus and Its Application Potential with Cooked Chicken. Food Microbiol. 2018, 72, 176–184. [Google Scholar] [CrossRef]
- Normanno, G.; Firinu, A.; Virgilio, S.; Mula, G.; Dambrosio, A.; Poggiu, A.; Decastelli, L.; Mioni, R.; Scuota, S.; Bolzoni, G.; et al. Coagulase-Positive Staphylococci and Staphylococcus aureus in Food Products Marketed in Italy. Int. J. Food Microbiol. 2005, 98, 73–79. [Google Scholar] [CrossRef]
- Peton, V.; le Loir, Y. Staphylococcusaureus in Veterinary Medicine. Infect Genet Evol 2014, 21, 602–615. [Google Scholar] [CrossRef]
- Lv, G.; Jiang, R.; Zhang, H.; Wang, L.; Li, L.; Gao, W.; Zhang, H.; Pei, Y.; Wei, X.; Dong, H.; et al. Molecular Characteristics of Staphylococcus aureus From Food Samples and Food Poisoning Outbreaks in Shijiazhuang, China. Front. Microbiol. 2021, 12, 1436. [Google Scholar] [CrossRef]
- Cha, Y.; Son, B.; Ryu, S. Effective Removal of Staphylococcal Biofilms on Various Food Contact Surfaces by Staphylococcus aureus Phage Endolysin LysCSA13. Food Microbiol. 2019, 84, 103245. [Google Scholar] [CrossRef]
- Chevallereau, A.; Pons, B.J.; van Houte, S.; Westra, E.R. Interactions between Bacterial and Phage Communities in Natural Environments. Nat. Rev. Microbiol. 2022, 20, 49–62. [Google Scholar] [CrossRef]
- Mills, S.; Ross, R.P.; Hill, C. Bacteriocins and bacteriophage; a narrow-minded approach to food and gut microbiology. FEMS Microbiol. Rev. 2017, 41, 129–153. [Google Scholar] [CrossRef]
- Özel, B.; Şimşek, Ö.; Akçelik, M.; Saris, P.E. Innovative approaches to nisin production. Appl. Microbiol. Biotechnol. 2018, 102, 6299–6307. [Google Scholar] [CrossRef]
- Wang, C.; Yang, J.; Zhu, X.; Lu, Y.; Xue, Y.; Lu, Z. Effects of Salmonella bacteriophage, nisin and potassium sorbate and their combination on safety and shelf life of fresh chilled pork. Food Control 2017, 73, 869–877. [Google Scholar] [CrossRef]
- Duc, H.M.; Son, H.M.; Ngan, P.H.; Sato, J.; Masuda, Y.; Honjoh, K.I.; Miyamoto, T. Isolation and application of bacteriophages alone or in combination with nisin against planktonic and biofilm cells of Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2020, 104, 5145–5158. [Google Scholar] [CrossRef]
- Wittebole, X.; de Roock, S.; Opal, S.M. A Historical Overview of Bacteriophage Therapy as an Alternative to Antibiotics for the Treatment of Bacterial Pathogens. Virulence 2014, 5, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Hussain, F.A.; Dubert, J.; Elsherbini, J.; Murphy, M.; Vaninsberghe, D.; Arevalo, P.; Kauffman, K.; Rodino-Janeiro, B.K.; Gavin, H.; Gomez, A.; et al. Rapid evolutionary turnover of mobile genetic elements drives bacterial resistance to phages. Science 2021, 374, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Martínez, B.; Obeso, J.M.; Rodríguez, A.; García, P. Nisin-bacteriophage cross resistance in Staphylococcus aureus. Int. J. Food Microbiol. 2008, 122, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Jasim, H.N.; Hafidh, R.R.; Abdulamir, A.S. Formation of Therapeutic Phage Cocktail and Endolysin to Highly Studymulti-Drug Resistant Acinetobacter Baumannii: In Vitro and in Vivo. Iran. J. Basic Med. Sci. 2018, 21, 1100–1108. [Google Scholar]
- Vázquez, R.; Domenech, M.; Iglesias-Bexiga, M.; Menéndez, M.; García, P. Csl2, a Novel Chimeric Bacteriophage Lysin to Fight Infections Caused by Streptococcus Suis, an Emerging Zoonotic Pathogen. Sci. Rep. 2017, 7, 16506. [Google Scholar] [CrossRef]
- Yu, J.H.; Lim, J.A.; Chang, H.J.; Park, J.H. Characteristics and Lytic Activity of Phage-Derived Peptidoglycan Hydrolase, LysSAP8, as a Potent Alternative Biocontrol Agent for Staphylococcus aureus. J. Microbiol. Biotechnol. 2019, 29, 1916–1924. [Google Scholar] [CrossRef]
- Gough, R.; Gómez-Sala, B.; O’Connor, P.M.; Rea, M.C.; Miao, S.; Hill, C.; Brodkorb, A. A Simple Method for the Purification of Nisin. Probiotics Antimicrob. Proteins 2017, 9, 363–369. [Google Scholar] [CrossRef]
- Pinto, M.S.; de Carvalho, A.F.; dos Santos Pires, A.C.; Souza, A.A.C.; da Silva, P.H.F.; Sobral, D.; de Paula, J.C.J.; de Lima Santos, A. The effects of nisin on Staphylococcus aureus count and the physicochemical properties of Traditional Minas Serro cheese. Int. Dairy J. 2011, 21, 90–96. [Google Scholar] [CrossRef]
- Yang, J.M.; Moon, G.S. Isolation of a Lactococcus lactis strain producing anti-staphylococcal bacteriocin. Korean J. Food Sci. Anim. Resour. 2018, 38, 1315–1321. [Google Scholar] [CrossRef]
- Teh, C.H.; Nazni, W.A.; Nurulhusna, A.H.; Norazah, A.; Lee, H.L. Determination of antibacterial activity and minimum inhibitory concentration of larval extract of fly via resazurin-based turbidometric assay. BMC Microbiol. 2017, 17, 36. [Google Scholar] [CrossRef]
- Kaláb, M.; Yang, A.F.; Chabot, D. Conventional scanning electron microscopy of bacteria. Infocus Mag. 2008, 10, 42–61. [Google Scholar] [CrossRef]
- Yun, B.; Lee, H.-S.; An, H.M.; Kim, W.-I.; Kim, H.-Y.; Han, S.; Kim, H.-J.; Ryu, J.-G.; Kim, S.-R. Effect of Chlorine Dioxide and Sodium Hypochlorite Treatment on the Reduction of Foodborne Pathogen in Korean Chive. J. Food Hyg. Saf. 2017, 32, 154–162. [Google Scholar] [CrossRef]
- Carfora, V.; Giacinti, G.; Sagrafoli, D.; Marri, N.; Giangolini, G.; Alba, P.; Feltrin, F.; Sorbara, L.; Amoruso, R.; Caprioli, A.; et al. Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus in Dairy Sheep and in-Contact Humans: An Intra-Farm Study. J. Dairy Sci. 2016, 99, 4251–4258. [Google Scholar] [CrossRef]
- Giacinti, G.; Carfora, V.; Caprioli, A.; Sagrafoli, D.; Marri, N.; Giangolini, G.; Amoruso, R.; Iurescia, M.; Stravino, F.; Dottarelli, S.; et al. Prevalence and Characterization of Methicillin-Resistant Staphylococcus aureus Carrying MecA or MecC and Methicillin-Susceptible Staphylococcus aureus in Dairy Sheep Farms in Central Italy. J. Dairy Sci. 2017, 100, 7857–7863. [Google Scholar] [CrossRef]
- Spanu, V.; Scarano, C.; Cossu, F.; Pala, C.; Spanu, C.; de Santis, E.P.L. Antibiotic Resistance Traits and Molecular Subtyping of Staphylococcus aureus Isolated from Raw Sheep Milk Cheese. J. Food Sci. 2014, 79, M2066–M2071. [Google Scholar] [CrossRef]
- Lee, A.S.; De Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Prim. 2018, 4, 18033. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; García, P. Bacteriophages as weapons against bacterial biofilms in the food industry. Front. Microbiol. 2016, 7, 825. [Google Scholar] [CrossRef]
- Jevons, M.P. “Celbenin”-resistant staphylococci. Br. Med. J. 1961, 1, 124. [Google Scholar] [CrossRef]
- International Working Group on the Classification of Staphylococcal Cassette Chromosome Elements (IWG-SCC). Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 53, 4961. [Google Scholar] [CrossRef]
- Hartman, B.J.; Tomasz, A. Low-affinity penicillin-binding protein associated with beta-lactam resistance in Staphylococcus aureus. J. Bacteriol. 1984, 158, 513–516. [Google Scholar] [CrossRef]
- García-Álvarez, L.; Holden, M.T.; Lindsay, H.; Webb, C.R.; Brown, D.F.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; van Alen, S.; Idelevich, E.A.; Schleimer, N.; Seggewiß, J.; Mellmann, A.; Kaspar, U.; Peters, G. Plasmid-encoded transferable mecB-mediated methicillin resistance in Staphylococcus aureus. Emerg. Infect. Dis. 2018, 24, 242. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage Cocktails and the Future of Phage Therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Leung, C.Y.; Henry, M.; Morello, E.; Singh, D.; di Santo, J.P.; Weitz, J.S.; Debarbieux, L. Synergy between the Host Immune System and Bacteriophage Is Essential for Successful Phage Therapy against an Acute Respiratory Pathogen. Cell Host Microbe 2017, 22, 38–47.e4. [Google Scholar] [CrossRef]
- Akmal, M.; Rahimi-Midani, A.; Hafeez-Ur-rehman, M.; Hussain, A.; Choi, T.J. Isolation, Characterization, and Application of a Bacteriophage Infecting the Fish Pathogen Aeromonas Hydrophila. Pathogens 2020, 9, 215. [Google Scholar] [CrossRef]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage Therapy: An Alternative to Antibiotics in the Age of Multi-Drug Resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162. [Google Scholar] [CrossRef]
- McCallin, S.; Sacher, J.C.; Zheng, J.; Chan, B.K. Current State of Compassionate Phage Therapy. Viruses 2019, 11, 343. [Google Scholar] [CrossRef]
- Hyman, P. Phages for phage therapy: Isolation, characterization, and host range breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef]
- Cui, Z.; Guo, X.; Dong, K.; Zhang, Y.; Li, Q.; Zhu, Y.; Zeng, L.; Tang, R.; Li, L. Safety assessment of Staphylococcus phages of the family Myoviridae based on complete genome sequences. Sci. Rep. 2017, 7, 41259. [Google Scholar] [CrossRef]
- Brum, L.F.W.; dos Santos, C.; Zimnoch Santos, J.H.; Brandelli, A. Structured Silica Materials as Innovative Delivery Systems for the Bacteriocin Nisin. Food Chem. 2021, 366, 130599. [Google Scholar] [CrossRef]
- Niaz, T.; Shabbir, S.; Noor, T.; Abbasi, R.; Imran, M. Alginate-Caseinate Based PH-Responsive Nano-Coacervates to Combat Resistant Bacterial Biofilms in Oral Cavity. Int. J. Biol. Macromol. 2020, 156, 1366–1380. [Google Scholar] [CrossRef]
- Wiedemann, I.; Breukink, E.; van Kraaij, C.; Kuipers, O.P.; Bierbaum, G.; de Kruijff, B.; Sahl, H.G. Specific binding of nisin to the peptidoglycan precursor lipid II combines pore formation and inhibition of cell wall biosynthesis for potent antibiotic activity. J. Biol. Chem. 2001, 276, 1772–1779. [Google Scholar] [CrossRef]
- García, P.; Martínez, B.; Rodríguez, L.; Rodríguez, A. Synergy between the Phage Endolysin LysH5 and Nisin to Kill Staphylococcus aureus in Pasteurized Milk. Int. J. Food Microbiol. 2010, 141, 151–155. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | S. aureus (Log CFU/mL) |
|---|---|
| Control (DW) | 5.90 |
| Sap8 endolysin (µM) | |
| 0.001 µM | 5.83 |
| 0.01 µM | 5.39 |
| 0.1 µM | 4.88 |
| 1 µM | 3.55 |
| Nisin (IU/mL) | |
| 9 IU/mL | 4.53 |
| 18 IU/mL | 4.26 |
| SAP8 endolysin (µM) + Nisin (IU/mL) | |
| 0.01 µM + 9 IU/mL | 4.26 |
| 0.01 µM + 18 IU/mL | 3.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-G.; Khan, S.A.; Lee, Y.-D.; Park, J.-H.; Moon, G.-S. Combination Treatment for Inhibition of the Growth of Staphylococcus aureus with Recombinant SAP8 Endolysin and Nisin. Antibiotics 2022, 11, 1185. https://doi.org/10.3390/antibiotics11091185
Kim S-G, Khan SA, Lee Y-D, Park J-H, Moon G-S. Combination Treatment for Inhibition of the Growth of Staphylococcus aureus with Recombinant SAP8 Endolysin and Nisin. Antibiotics. 2022; 11(9):1185. https://doi.org/10.3390/antibiotics11091185
Chicago/Turabian StyleKim, Seon-Gyu, Shehzad Abid Khan, Young-Duck Lee, Jong-Hyun Park, and Gi-Seong Moon. 2022. "Combination Treatment for Inhibition of the Growth of Staphylococcus aureus with Recombinant SAP8 Endolysin and Nisin" Antibiotics 11, no. 9: 1185. https://doi.org/10.3390/antibiotics11091185