
Characterization of Carbapenemase-Producing Klebsiella pneumoniae Isolates from Two Romanian Hospitals Co-Presenting Resistance and Heteroresistance to Colistin
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Antimicrobial Susceptibility Testing
2.2. Whole-Genome Sequencing Data Analysis
2.2.1. Quality Metrics for the Assembled Draft Genomes
2.2.2. Genetic Determinants of Antimicrobial Resistance
2.2.3. Molecular Serotyping, Plasmid Replicon Identification, and Sequence Type Determination
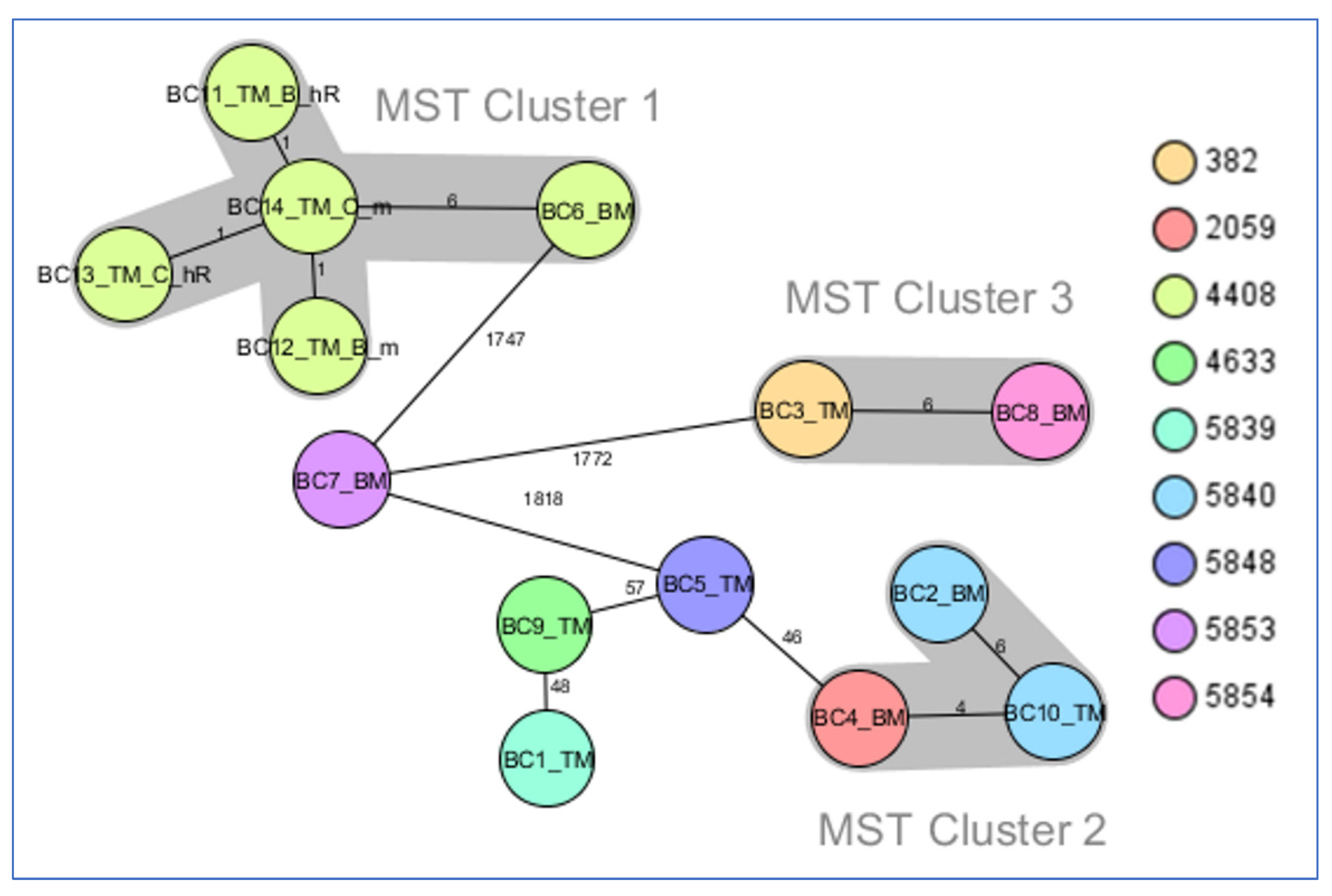
- MST cluster 1, comprising blaKPC-2 producers: two pairs of ChR with their corresponding resistant mutants (from Târgu Mureș) and BC6_BM (from Baia Mare), collected in the period 2019–2020.
- MST cluster 2, consisting of three blaOXA-48 strains, BC2_BM, BC4_BM, and BC10_TM (two from Baia Mare and one from Târgu Mureș), collected in the period 2018–2019.
- MST cluster 3, comprising two blaKPC-2 strains, BC3_TM and BC8_BM (from Târgu Mureș and Baia Mare, respectively), collected in the period 2017–2018.
3. Discussion
Future Directions
4. Materials and Methods
4.1. Bacterial Strains, Setting, and Design of the Study
4.2. Demographic Data of Patients
4.3. Phenotypic Bacterial Identification and Antimicrobial Susceptibility Testing
4.4. Genotypic Characterization Using WGS
4.4.1. Sequencing and Assembly of Draft Genomes
4.4.2. Resistome Analysis
4.4.3. Molecular Typing of Isolates and Plasmid Profiling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Logan, L.K.; Weinstein, R.A. The Epidemiology of carbapenem-resistant Enterobacteriaceae: The impact and evolution of a global menace. J. Infect. Dis. 2017, 15 (Suppl. S1), S28–S36. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Carbapenem-Resistant Enterobacteriaceae; Second Update; Rapid Risk Assessment; ECDC: Stockholm, Sweden, 2019; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/carbapenem-resistant-enterobacteriaceae-risk-assessment-rev-2.pdf (accessed on 10 August 2022).
- Rodriguez-Bano, J.; Gutierrez-Gutierrez, B.; Machuca, I.; Pascuala, A. Treatment of infections caused by extended-spectrum-beta-lactamase-, AmpC-, and carbapenemase-producing Enterobacteriaceae. Clin. Microbiol. Rev. 2018, 31, e00079-17. [Google Scholar] [CrossRef]
- Lan, P.; Jiang, Y.; Zhou, J.; Yu, Y. A global perspective on the convergence of hypervirulence and carbapenem resistance in Klebsiella pneumoniae. J. Glob. Antimicrob. Resist. 2021, 25, 26–34. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Antimicrobial Resistance in the EU/EEA (EARS-Net)—Annual Epidemiological Report 2019; ECDC: Stockholm, Sweden, 2020; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/surveillance-antimicrobial-resistance-Europe-2019.pdf (accessed on 10 August 2022).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L. Epidemiology and diagnostics of carbapenem resistance in Gram-negative bacteria. Clin. Infect. Dis. 2019, 69 (Suppl. S7), S521–S528. [Google Scholar] [CrossRef]
- Nordmann, P.; Dortet, L.; Poirel, L. Carbapenem resistance in Enterobacteriaceae: Here is the storm! Trends Mol. Med. 2012, 18, 263–272. [Google Scholar] [CrossRef]
- van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacte-riaceae. Virulence 2017, 8, 460–469. [Google Scholar] [CrossRef]
- David, S.; Reuter, S.; Harris, S.R.; Glasner, C.; Feltwell, T.; Argimon, S.; Abudahab, K.; Goater, R.; Giani, T.; Errico, G.; et al. Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 2019, 4, 1919–1929. [Google Scholar] [CrossRef]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- European Centre for Disease Prevention and Control. Emergence of Hypervirulent Klebsiella pneumoniae ST23 Carrying Carbapenemase Genes in EU/EEA Countries; Rapid Risk Assessment; ECDC: Stockholm, Sweden, 2021; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/Emergence-of-hypervirulent-Klebsiella-pneumoniae-ST23-carrying-carbapenemase-genes.pdf (accessed on 10 August 2022).
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis––10 years. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Expert Consensus Protocol on Colistin Resistance Detection and Characterisation for the Survey of Carbapenem- and/or Colistin-Resistant Enterobacteriaceae; Version 1.0; ECDC: Stockholm, Sweden, 2019; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/expert-consensus-protocol-colistin-resistance.pdf (accessed on 10 August 2022).
- World Health Organization. The Detection and Reporting of Colistin Resistance, 2nd ed.; Global Antimicrobial Resistance and use Surveillance System (GLASS); WHO: Geneva, Switzerland, 2021; Available online: https://apps.who.int/iris/bitstream/handle/10665/343654/9789240019041-eng.pdf?sequence=1 (accessed on 10 August 2022).
- El-Sayed, M.A.E.G.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and its role in the era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef]
- Fordham, S.M.E.; Mantzouratou, A.; Sheridan, E. Prevalence of insertion sequence elements in plasmids relating to mgrB gene disruption causing colistin resistance in Klebsiella pneumoniae. MicrobiologyOpen 2022, 11, e1262. [Google Scholar] [CrossRef]
- Hamel, M.; Chatzipanagiotou, S.; Hadjadj, L.; Petinaki, E.; Papagianni, S.; Charalampaki, N.; Tsiplakou, S.; Papaioannou, V.; Skarmoutsou, N.; Spiliopoulou, I.; et al. Inactivation of mgrB gene regulator and resistance to colistin is becoming endemic in carbapenem-resistant Klebsiella pneumoniae in Greece: A nationwide study from 2014 to 2017. Int. J. Antimicrob. Agents 2020, 55, 105930. [Google Scholar] [CrossRef]
- Yang, T.; Wang, S.; Lin, J.; Griffith, B.T.; Lian, S.; Hong, Z.; Lin, L.; Lu, P.; Tseng, S. Contributions of insertion sequences conferring colistin resistance in Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2020, 55, 105894. [Google Scholar] [CrossRef]
- El-Halfawy, O.M.; Valvano, M.A. Antimicrobial heteroresistance: An emerging field in need of clarity. Clin. Microbiol. Rev. 2015, 28, 191–207. [Google Scholar] [CrossRef]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef]
- Stojowska-Swedrzynska, K.; Lupkowska, A.; Kuczynska-Wisnik, D.; Laskowska, E. Antibiotic heteroresistance in Klebsiella pneumoniae. Int. J. Mol. Sci. 2021, 23, 449. [Google Scholar] [CrossRef]
- Band, V.I.; Satola, S.W.; Smith, R.D.; Hufnagel, D.A.; Bower, C.; Conley, A.B.; Rishishwar, L.; Dale, S.E.; Hardy, D.J.; Vargas, R.L.; et al. Colistin heteroresistance is largely undetected among Carbapenem-Resistant Enterobacterales in the United States. mBio 2021, 12, e02881-20. [Google Scholar] [CrossRef]
- Koser, C.U.; Ellington, M.J.; Peacock, S.J. Whole-genome sequencing to control antimicrobial resistance. Trends Genet. 2014, 30, 401–407. [Google Scholar] [CrossRef]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Centre for Disease Prevention and Control, Surveillance Atlas of Infectious Diseases. 2021. Available online: https://www.ecdc.europa.eu/en/surveillance-atlas-infectious-diseases (accessed on 10 August 2022).
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM metallo-β-lactamases and their bacterial producers in health care settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef]
- Pitt, M.E.; Elliott, A.G.; Cao, M.D.; Ganesamoorthy, D.; Karaiskos, I.; Giamarellou, H.; Abboud, C.S.; Blaskovich, M.A.T.; Cooper, M.A.; Coin, L.J.M. Multifactorial chromosomal variants regulate polymyxin resistance in extensively drug-resistant Klebsiella pneumoniae. Microb. Genom. 2018, 4, e000158. [Google Scholar] [CrossRef]
- Esposito, E.P.; Cervoni, M.; Bernardo, M.; Crivaro, V.; Cuccurullo, S.; Imperi, F.; Zarrilli, R. Molecular epidemiology and virulence profiles of colistin-resistant Klebsiella pneumoniae blood isolates from the Hospital Agency “Ospedale dei Colli,” Naples, Italy. Front. Microbiol. 2018, 9, 1463. [Google Scholar] [CrossRef]
- Lomonaco, S.; Crawford, M.A.; Lascols, C.; Timme, R.E.; Anderson, K.; Hodge, D.R.; Fisher, D.J.; Pillai, S.P.; Morse, S.A.; Khan, E.; et al. Resistome of carbapenem- and colistin-resistant Klebsiella pneumoniae clinical isolates. PLoS ONE 2018, 13, e0198526. [Google Scholar] [CrossRef]
- Ruppe, E.; Cherkaoui, A.; Charretier, Y.; Girard, M.; Schicklin, S.; Lazarevic, V.; Schrenzel, J. From genotype to antibiotic susceptibility phenotype in the order Enterobacterales: A clinical perspective. Clin. Microbiol. Infect. 2020, 26, 643.e1–643.e7. [Google Scholar] [CrossRef]
- Ellington, M.J.; Ekelund, O.; Aarestrup, F.M.; Canton, R.; Doumith, M.; Giske, C.; Grundman, H.; Hasman, H.; Holden, M.T.G.; Hopkins, K.L.; et al. The role of whole genome sequencing in antimicrobial susceptibility testing of bacteria: Report from the EUCAST Subcommittee. Clin. Microbiol. Infect. 2017, 23, 2–22. [Google Scholar] [CrossRef]
- Mancini, S.; Marchesi, M.; Imkamp, F.; Wagner, K.; Keller, P.M.; Quiblier, C.; Bodendoerfer, E.; Courvalin, P.; Bottger, E.C. Population-based inference of aminoglycoside resistance mechanisms in Escherichia coli. eBioMedicine 2019, 46, 184–192. [Google Scholar] [CrossRef]
- Vaziri, S.; Afsharian, M.; Mansouri, F.; Azizi, M.; Nouri, F.; Madadi-Goli, N.; Afshar, Z.M.; Zamanian, M.H.; Alvandi, A.; Ahmadi, K. Frequency of qnr and aac(6′)Ib-cr genes among ESBL-producing Klebsiella pneumoniae strains isolated from burn patients in Kermanshah, Iran. Jundishapur J. Microbiol. 2020, 13, e100348. [Google Scholar] [CrossRef]
- Galani, I.; Nafplioti, K.; Adamou, P.; Karaiskos, I.; Giamarellou, H.; Souli, M. Nationwide epidemiology of carbapenem resistant Klebsiella pneumoniae isolates from Greek hospitals, with regards to plazomicin and aminoglycoside resistance. BMC Infect. Dis. 2019, 19, 167. [Google Scholar] [CrossRef]
- Shelenkov, A.; Mikhaylova, Y.; Yanushevich, Y.; Samoilov, A.; Petrova, L.; Fomina, V.; Gusarov, V.; Zamyatin, M.; Shagin, D.; Akimkin, V. Molecular typing, characterization of antimicrobial resistance, virulence profiling and analysis of whole-genome sequence of clinical Klebsiella pneumoniae isolates. Antibiotics 2020, 9, 261. [Google Scholar] [CrossRef]
- Szekely, E.; Damjanova, I.; Janvari, L.; Vas, K.E.; Molnar, S.; Bilca, D.V.; Lorinczi, L.K.; Toth, A. First description of blaNDM−1, blaOXA−48, blaOXA−181 producing Enterobacteriaceae strains in Romania, Int. J. Med. Microbiol. 2013, 303, 697–700. [Google Scholar] [CrossRef]
- Dortet, L.; Flonta, M.; Boudehen, Y.M.; Creton, E.; Bernabeu, S.; Vogel, A.; Naas, T. Dissemination of carbapenemase-producing Enterobacteriaceae and Pseudomonas aeruginosa in Romania. Antimicrob. Agents Chemother. 2015, 59, 7100–7103. [Google Scholar] [CrossRef]
- Poirel, L.; Revathi, G.; Bernabeu, S.; Nordmann, P. Detection of NDM-1-Producing Klebsiella pneumoniae in Kenya. Antimicrob. Agents Chemother. 2011, 55, 934–936. [Google Scholar] [CrossRef]
- Gokmen, T.G.; Nagiyev, T.; Meral, M.; Onlen, C.; Heydari, F.; Koksal, F. NDM-1 and rmtC-producing Klebsiella pneumoniae isolates in Turkey. Jundishapur J. Microbiol. 2016, 9, e33990. [Google Scholar] [CrossRef]
- Al-Agamy, M.H.; El-Mahdy, T.S.; Radwan, H.H.; Poirel, L. Cooccurrence of NDM-1, ESBL, rmtC, aac(6)-Ib, and qnrB in clonally related Klebsiella pneumoniae isolates together with coexistence of CMY-4 and aac(6)-Ib in Enterobacter cloacae isolates from Saudi Arabia. BioMed Res. Int. 2019, 2019, 6736897. [Google Scholar] [CrossRef]
- Galimand, M.; Courvalin, P.; Lambert, T. Plasmid-mediated high-level resistance to aminoglycosides in Enterobacteriaceae due to 16S rRNA methylation. Antimicrob. Agents Chemother. 2003, 47, 2565–2571. [Google Scholar] [CrossRef]
- Wachino, J.I.; Doi, Y.; Arakawa, Y. Aminoglycoside resistance: Updates with a focus on acquired 16S ribosomal RNA methyltransferases. Infect. Dis. Clin. N. Am. 2020, 34, 887–902. [Google Scholar] [CrossRef]
- Fournier, C.; Poirel, L.; Despont, S.; Kessler, J.; Nordmann, P. Increasing trends of association of 16S rRNA methylases and carbapenemases in Enterobacterales clinical isolates from Switzerland, 2017–2020. Microorganisms 2022, 10, 615. [Google Scholar] [CrossRef]
- Sun, D.; Rubio-Aparicio, D.; Nelson, K.; Dudley, M.N.; Lomovskaya, O. Meropenem-vaborbactam resistance selection, resistance prevention, and molecular mechanisms in mutants of KPC-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2017, 61, e01694-17. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, S. Meropenem/vaborbactam: A review in complicated urinary tract infections. Drugs 2018, 78, 1259–1270. [Google Scholar] [CrossRef]
- Gaibani, P.; Lombardo, D.; Bussini, L.; Bovo, F.; Munari, B.; Giannella, M.; Bartoletti, M.; Viale, P.; Lazzarotto, T.; Ambretti, S. Epidemiology of meropenem/vaborbactam resistance in KPC-producing Klebsiella pneumoniae causing bloodstream infections in Northern Italy, 2018. Antibiotics 2021, 10, 536. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Sun, D.; Rubio-Aparicio, D.; Nelson, K.; Tsivkovski, R.; Griffith, D.C.; Dudley, M.N. Vaborbactam: Spectrum of beta-lactamase inhibition and impact of resistance mechanisms on activity in Enterobacteriaceae. Antimicrob. Agents Chemother. 2017, 61, e01443-17. [Google Scholar] [CrossRef]
- Gogry, F.A.; Siddiqui, M.T.; Sultan, I.; Haq, Q.M.R. Current update on intrinsic and acquired colistin resistance mechanisms in bacteria. Front. Med. 2021, 8, 677720. [Google Scholar] [CrossRef] [PubMed]
- Jayol, A.; Nordmann, P.; Andre, C.; Poirel, P.; Dubois, V. Evaluation of three broth microdilution systems to determine colistin susceptibility of Gram-negative bacilli. J. Antimicrob. Chemother. 2018, 73, 1272–1278. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Zhu, Y.; Jia, P.; Li, X.; Jia, X.; Yu, W.; Cui, Y.; Yang, R.; Xia, W.; et al. Emergence of colistin-resistant hypervirulent Klebsiella pneumoniae (CoR-HvKp) in China. Emerg. Microbes. Infect. 2022, 11, 648–661. [Google Scholar] [CrossRef] [PubMed]
- Kidd, T.J.; Mills, G.; Sa-Pessoa, J.; Dumigan, A.; Frank, C.G.; Insua, J.L.; Ingram, R.; Hobley, L.; Bengoechea, J.A. A Klebsiella pneumoniae antibiotic resistance mechanism that subdues host defences and promotes virulence. EMBO Mol. Med. 2017, 9, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Zaman, T.U.; Albladi, M.; Siddique, M.I.; Aljohani, S.M.; Balkhy, H.H. Insertion element mediated mgrB disruption and presence of ISKpn28 in colistin-resistant Klebsiella pneumoniae isolates from Saudi Arabia. Infect. Drug Resist. 2018, 11, 1183–1187. [Google Scholar] [CrossRef]
- Azam, M.; Gaind, R.; Yadav, G.; Sharma, A.; Upmanyu, K.; Jain, M.; Singh, R. Colistin resistance among multiple sequence types of Klebsiella pneumoniae is associated with diverse resistance mechanisms: A report from India. Front. Microbiol. 2021, 12, 609840. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Lin, T.L.; Pan, Y.J.; Wang, Y.P.; Lin, Y.; Wang, J.T. Colistin resistance mechanisms in Klebsiella pneumoniae strains from Taiwan. Antimicrob. Agents Chemother. 2015, 59, 2909–2913. [Google Scholar] [CrossRef]
- Castanheira, M.; Doyle, T.B.; Davis, A.P.; Deshpande, L.M.; Mendes, R.E. Disruption of mgrB and Alterations on pmrB Are Most Common Resistance Mechanisms among Colistin-Resistance among Klebsiella pneumoniae from a Global Surveillance Program. ASM Microbe 2018. Available online: https://www.jmilabs.com/data/posters/ASM-Microbe-2018-Klebsiella-pneumoniae-colistin-resistance.pdf (accessed on 10 August 2022).
- Binsker, U.; Kasbohrer, A.; Hammerl, J.A. Global colistin use: A review of the emergence of resistant Enterobacterales and the impact on their genetic basis. FEMS Microbiol. Rev. 2022, 46, fuab049. [Google Scholar] [CrossRef] [PubMed]
- Jayol, A.; Poirel, L.; Brink, A.; Villegas, M.V.; Yilmaz, M.; Nordmann, P. Resistance to colistin associated with a single amino acid change in protein PmrB among Klebsiella pneumoniae isolates of worldwide origin. Antimicrob. Agents Chemother. 2014, 58, 4762–4766. [Google Scholar] [CrossRef] [PubMed]
- Elias, R.; Duarte, A.; Perdigao, J. A Molecular Perspective on colistin and Klebsiella pneumoniae: Mode of action, resistance genetics, and phenotypic susceptibility. Diagnostics 2021, 11, 1165. [Google Scholar] [CrossRef]
- Pitt, M.E.; Cao, M.D.; Butler, M.S.; Ramu, S.; Ganesamoorthy, D.; Blaskovich, M.A.T.; Coin, L.J.M.; Cooper, M.A. Octapeptin C4 and polymyxin resistance occur via distinct pathways in an epidemic XDR Klebsiella pneumoniae ST258 isolate. J. Antimicrob. Chemother. 2019, 74, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Lellouche, J.; Schwartz, D.; Elmalech, N.; Ben Dalak, M.A.; Temkin, E.; Paul, M.; Geffen, Y.; Yahav, D.; Eliakim-Raz, N.; DuranteMangoni, E.; et al. Combining VITEK 2 with colistin agar dilution screening assist timely reporting of colistin susceptibility. Clin. Microbiol. Infect. 2019, 25, 711–716. [Google Scholar] [CrossRef]
- Pfennigwerth, N.; Kaminski, A.; Korte-Berwanger, M.; Pfeifer, Y.; Simon, M.; Werner, G.; Jantsch, J.; Marlinghaus, L.; Gatermann, S.G. Evaluation of six commercial products for colistin susceptibility testing in Enterobacterales. Clin. Microbiol. Infect. 2019, 25, 1385–1389. [Google Scholar] [CrossRef]
- Foldes, A.; Szekely, E.; Voidazan, S.T.; Dobreanu, M. Comparison of six phenotypic assays with reference methods for assessing colistin resistance in clinical isolates of carbapenemase-producing Enterobacterales: Challenges and opportunities. Antibiotics 2022, 11, 377. [Google Scholar] [CrossRef]
- BioMerieux. Urgent Product Correction Notice. 2017. Available online: https://www.bfarm.de/SharedDocs/Kundeninfos/EN/08/2017/04963-17_kundeninfo_en.pdf?__blob=publicationFile&v=1 (accessed on 10 August 2022).
- Cheng, Y.H.; Lin, T.L.; Lin, Y.T.; Wang, J.T. Amino acid substitutions of CrrB responsible for resistance to colistin through CrrC in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2016, 60, 3709–3716. [Google Scholar] [CrossRef]
- Bardet, L.; Rolain, J.M. Development of new tools to detect colistin-resistance among Enterobacteriaceae strains. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 3095249. [Google Scholar] [CrossRef]
- Jayol, A.; Nordmann, P.; Brink, A.; Poirel, L. Heteroresistance to colistin in Klebsiella pneumoniae associated with alterations in the PhoPQ regulatory system. Antimicrob. Agents Chemother. 2015, 59, 2780–2784. [Google Scholar] [CrossRef] [PubMed]
- Bardet, L.; Baron, S.; Leangapichart, T.; Okdah, L.; Diene, S.M.; Rolain, J.M. Deciphering heteroresistance to colistin in a Klebsiella pneumoniae isolate from Marseille, France. Antimicrob. Agents Chemother. 2017, 61, e00356-17. [Google Scholar] [CrossRef] [PubMed]
- Cheong, H.S.; Kim, S.Y.; Wi, Y.M.; Peck, K.R.; Ko, K.S. Colistin heteroresistance in Klebsiella pneumoniae isolates and diverse mutations of PmrAB and PhoPQ in resistant subpopulations. J. Clin. Med. 2019, 8, 1444. [Google Scholar] [CrossRef]
- Halaby, T.; Kucukkose, E.; Janssen, A.B.; Rogers, M.R.C.; Doorduijn, D.J.; Van Der Zanden, A.G.M.; Al Naiemi, N.; Vandenbroucke-Grauls, C.M.J.E.; Van Schaik, W. Genomic characterization of colistin heteroresistance in Klebsiella pneumoniae during a nosocomial outbreak. Antimicrob. Agents Chemother. 2016, 60, 6837–6843. [Google Scholar] [CrossRef] [PubMed]
- Peirano, G.; Chen, L.; Kreiswirth, B.N.; Pitout, J.D.D. Emerging antimicrobial-resistant high-risk Klebsiella pneumoniae clones ST307 and ST147. Antimicrob. Agents Chemother. 2020, 64, e01148-20. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Outbreak of Carbapenemase-Producing (NDM-1 and OXA-48) and Colistin-Resistant Klebsiella Pneumoniae ST307, North-East Germany, 2019; ECDC: Stockholm, Sweden, 2019; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/Klebsiella-pneumoniae-resistance-Germany-risk-assessment.pdf (accessed on 10 August 2022).
- Surleac, M.; Czobor Barbu, I.; Paraschiv, S.; Popa, L.I.; Gheorghe, I.; Marutescu, L.; Popa, M.; Sarbu, I.; Talapan, D.; Nita, M. Whole genome sequencing snapshot of multi-drug resistant Klebsiella pneumoniae strains from hospitals and receiving wastewater treatment plants in Southern Romania. PLoS ONE 2020, 15, e0228079. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, M.; D’Andrea, M.M.; Pelegrin, A.C.; Mirande, C.; Brkic, S.; Cirkovic, I.; Goossens, H.; Rossolini, G.M.; van Belkum, A. Genomic epidemiology of carbapenem- and colistin-resistant Klebsiella pneumoniae isolates from Serbia: Predominance of ST101 strains carrying a novel OXA-48 plasmid. Front. Microbiol. 2020, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Roe, C.C.; Vazquez, A.J.; Esposito, E.P.; Zarrilli, R.; Sahl, J.W. Diversity, virulence, and antimicrobial resistance in isolates from the newly emerging Klebsiella pneumoniae ST101 lineage. Front. Microbiol. 2019, 10, 542. [Google Scholar] [CrossRef]
- Popa, L.I.; Gheorghe, I.; Czobor Barbu, I.; Surleac, M.; Paraschiv, S.; Marutescu, L.; Popa, M.; Gradisteanu Pîrcalabioru, G.; Talapan, D.; Niţă, M.; et al. Multidrug resistant Klebsiella pneumoniae ST101 clone survival chain from inpatients to hospital effluent after chlorine treatment. Front. Microbiol. 2021, 11, 610296. [Google Scholar] [CrossRef]
- Lam, M.M.C.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.H. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat. Commun. 2021, 12, 4188. [Google Scholar] [CrossRef]
- Becker, L.; Kaase, M.; Pfeifer, Y.; Fuchs, S.; Reuss, A.; von Laer, A.; Sin, M.A.; Korte-Berwanger, M.; Gatermann, S.; Werner, G. Genome-based analysis of carbapenemase-producing Klebsiella pneumoniae isolates from German hospital patients, 2008-2014. Antimicrob. Resist. Infect. Control 2018, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Molnar, S.; Vas, K.E.; Székely, E. Carbapenemase producing Enterobacterales in Romania: Investigating the origins. Revista Romana de Medicina de Laborator 2020, 28, 341–348. [Google Scholar] [CrossRef]
- Phan, H.T.T.; Stoesser, N.; Maciuca, I.E.; Toma, F.; Szekely, E.; Flonta, M.; Hubbard, A.T.M.; Pankhurst, L.; Do, T.; Peto, T.E.A.; et al. Illumina short-read and MinION long-read WGS to characterize the molecular epidemiology of an NDM-1 Serratia marcescens outbreak in Romania. J. Antimicrob. Chemother. 2018, 73, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Villa, L.; Garcia-Fernandez, A.; Fortini, D.; Carattoli, A. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 2010, 65, 2518–2529. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST), Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 12.0. 2022. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_12.0_Breakpoint_Tables.pdf (accessed on 10 August 2022).
- Tygacil. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2013/021821s026s031lbl.pdf (accessed on 10 August 2022).
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; M100-S27; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2017. [Google Scholar]
- van der Zwaluw, K.; de Haan, A.; Pluister, G.N.; Bootsma, H.J.; de Neeling, A.J.; Schouls, L.M. The carbapenem inactivation method (CIM), a simple and low-cost alternative for the Carba NP Test to assess phenotypic carbapenemase activity in Gram-negative rods. PLoS ONE 2015, 10, e0123690. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulilov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemotherapy. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Zankari, E.; Allesae, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [PubMed]
- Sim, N.L.; Kumar, P.; Hu, J.; Henikoff, S.; Schneider, G.; Ng, P.C. SIFT web server: Predicting effects of amino acid substitutions on proteins. Nucleic Acids Res. 2012, 40, W452–W457. [Google Scholar] [CrossRef] [PubMed]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.D.; Brisse, S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [PubMed]
- Junemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; von Haeseler, A.; Stoye, J.; et al. Updating benchtop sequencing performance comparison. Nat. Biotechnol. 2013, 31, 294–296, Erratum in: Nat. Biotechnol. 2013, 31, 1148. [Google Scholar] [CrossRef]
- Wick, R.R.; Heinz, E.; Holt, K.E.; Wyres, K.L. Kaptive Web: User-friendly capsule and lipopolysaccharide serotype prediction for Klebsiella genomes. J. Clin. Microbiol. 2018, 56, e00197-18. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Bacterial Strain | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antimicrobial Agent | BC1_TM | BC2_BM | BC3_TM | BC4_BM | BC5_TM | BC6_BM | BC7_BM | BC8_BM | BC9_TM | BC10_TM | BC11_TM_B_hR | BC12_TM_B_m | BC13_TM_C_hR | BC14_TM_C_m | |
| Ampicillin | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Amoxicillin–clavulanic acid | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Piperacillin–tazobactam | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Ceftazidime–avibactam * | R | S | S | S | S | S | R | S | S | S | S | S | S | S | |
| Meropenem–vaborbactam * | R | R | R | R | R | S | R | R | R | R | S | S | S | S | |
| Imipenem/cilastatin–relebactam * | R | I | S | R | S | S | R | S | R | R | S | S | S | S | |
| Cefuroxime | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Cefoxitin | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Cefotaxime | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Ceftazidime | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Ceftriaxone | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Cefepime | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Aztreonam | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Meropenem | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Imipenem | R | S■ | R | I▲ | R | R | R | R | R | S■ | R | R | R | R | |
| Ertapenem | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Doripenem * | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Gentamicin | R | S | R● | R | S | S | R | R● | R | S | S | S | S | S | |
| Amikacin | R | R | R | R | R | R | R | R | S | R | R | R | R | R | |
| Tobramycin | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Ciprofloxacin | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Levofloxacin | R | R | R | R | R | R | R | R | R | R | R | R | R | R | |
| Trimethoprim–sulfamethoxazole | R | S | R | S | R | R | R | R | S | S | R | R | R | R | |
| Chloramphenicol | R● | R● | R | R● | R | R | R | R | S | R● | R | R | R | R | |
| Tigecycline ** | S | S | S | I | S | S | S | S | S | I | S | S | S | S | |
| Colistin ** (MIC mg/L) | AST-N222 | NT | ≤0.5 S | 2S | NT | NT | NT | NT | NT | NT | NT | ≤0.5 S | ≥16 R | ≤0.5 S | ≥16 R |
| AST-XN05 | ≥16R | 1S | 4R | ≥16 R | 8R | ≥16R | ≥16 R | ≥16 R | ≥16 R | ≥16 R | ≤0.5 S | ≥16 R | ≤0.5 S | ≥16 R | |
| Bacterial Isolate | Micronaut MIC-Strip (MIC mg/L) | Etest, MHE (MIC mg/L) | COLR Medium | Rapid Polymyxin NP Test | CBDE (MIC mg/L) | BMD (MIC mg/L) |
|---|---|---|---|---|---|---|
| BC1_TM | 32 | 4 | Positive | Positive | ≥4 | 32 |
| BC2_BM | 16 | 2 | Positive | Positive | ≥4 | 8 |
| BC3_TM | 8 | 2 | Positive | Positive | ≥4 | 16 |
| BC4_BM | 16 | 4 | Positive | Positive | ≥4 | 16 |
| BC5_TM | 8 | 4 | Positive | Positive | ≥4 | 8 |
| BC6_BM | 32 | 16 | Positive | Positive | ≥4 | 32 |
| BC7_BM | 16 | 8 | Positive | Positive | ≥4 | 16 |
| BC8_BM | 8 | 2 | Positive | Positive | ≥4 | 8 |
| BC9_TM | 32 | 32 | Positive | Positive | ≥4 | 32 |
| BC10_TM | 16 | 4 | Positive | Positive | ≥4 | 16 |
| BC11_TM_B_hR | 0.5 | 0.25 | Negative | Positive | ≤1 | 0.25 |
| BC12_TM_B_m | >64 | 192 | Positive | Positive | ≥4 | >64 |
| BC13_TM_C_hR | 0.25 | 0.25 | Negative | Positive | ≤1 | 0.5 |
| BC14_TM_C_m | >64 | 128 | Positive | Positive | ≥4 | >64 |
| Bacterial Strain ID | Number of Contigs ≥ 200 bp | N50 | Draft Genome Length (bp) |
|---|---|---|---|
| BC1_TM | 122 | 173,972 | 5,474,448 |
| BC2_BM | 151 | 174,945 | 5,948,608 |
| BC3_TM | 177 | 203,739 | 5,596,147 |
| BC4_BM | 154 | 180,250 | 5,878,102 |
| BC5_TM | 157 | 171,932 | 5,749,801 |
| BC6_BM | 135 | 204,587 | 5,682,036 |
| BC7_BM | 124 | 197,811 | 5,731,764 |
| BC8_BM | 131 | 172,882 | 5,655,780 |
| BC9_TM | 141 | 173,978 | 5,589,246 |
| BC10_TM | 159 | 204,669 | 5,905,163 |
| BC11_TM_B_hR | 183 | 170,134 | 5,637,259 |
| BC12_TM_B_m | 215 | 172,730 | 5,669,127 |
| BC13_TM_C_hR | 188 | 151,322 | 5,628,400 |
| BC14_TM_C_m | 166 | 206,499 | 5,692,480 |
| Bacterial Strain | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Class | Gene | BC1_TM | BC2_BM | BC3_TM | BC4_BM | BC5_TM | BC6_BM | BC7_BM | BC8_BM | BC9_TM | BC10_TM | BC11_TM_B_hR | BC12_TM_B_m | BC13_TM_C_hR | BC14_TM_C_m |
| β-L | blaCMY-16 | x | |||||||||||||
| blaCTX-M-15 | x | x | x | x | x | x | x | x | x | x | x | x | |||
| blaKPC-2 | x | x | x | x | x | x | x | x | |||||||
| blaNDM-1 | x | x | |||||||||||||
| blaOXA-1 | x | x | |||||||||||||
| blaOXA-9 | x | x | x | x | x | x | x | ||||||||
| blaOXA-10 | x | ||||||||||||||
| blaOXA-48 | x | x | x | x | |||||||||||
| blaSHV-1 | x | x | x | x | x | ||||||||||
| blaSHV-11 | x | x | x | x | x | x | |||||||||
| blaSHV-12 | x | x | |||||||||||||
| blaSHV-28 | x | ||||||||||||||
| blaTEM-1 | x | x | x | x | x | x | x | x | x | x | x | x | |||
| Ag | aac(3)-IId | xG | |||||||||||||
| aac(3)-IIe | xG | xG | xG | ||||||||||||
| aac(6′)-Ib | xA,T,K | xA,T,K | xA,T,K | xA,T,K | xA,T,K | xA,T,K | xA,T,K | xA,T,K | xA,T,K | xA,T,K | xA,T,K | xA,T,K | |||
| aac(6′)-Ib-cr5 | xA,T,K | xA,T,K | |||||||||||||
| aadA1 | xS | xS | xS | xS | xS | xS | |||||||||
| aadA2 | xS | xS | xS | xS | xS | xS | xS | xS | |||||||
| aph(3′)-Ia | xK | xK | xK | xK | xK | xK | xK | xK | |||||||
| aph(3′)-VI | xA,K | xA,K | |||||||||||||
| aph(3′’)-Ib | xS | xS | |||||||||||||
| aph(6)-Id | xS | ||||||||||||||
| rmtC | xA,T,H,K,KS,S,SP | ||||||||||||||
| Gy | ble | x | x | ||||||||||||
| Fos | fosA | x | x | x | x | x | x | x | x | x | x | x | x | x | x |
| Ma | mph(A) | x | x | x | x | x | x | x | x | ||||||
| Ch | catA1 | x | x | x | x | x | x | x | |||||||
| cmlA5 | x | ||||||||||||||
| Ph | oqxA | x | x | x | x | x | x | x | x | x | x | x | x | x | |
| oqxB/B19/B20 | x | x | x | x | x | x | x | x | x | x | x | x | x | x | |
| Flq | aac(6′)-Ib-cr5 | x | x | ||||||||||||
| gyrAD87N | x | x | x | x | x | x | |||||||||
| gyrAS83I | x | x | x | x | x | x | x | x | |||||||
| gyrAS83Y | x | x | x | x | x | x | |||||||||
| oqxA | x | x | x | x | x | x | x | x | x | x | x | x | x | ||
| oqxB/B19/B20 | x | x | x | x | x | x | x | x | x | x | x | x | x | x | |
| parCS80I | x | x | x | x | x | x | x | x | x | x | x | x | x | x | |
| qnrS1 | x | ||||||||||||||
| Rf | arr-2 | x | |||||||||||||
| Sph | sul1 | x | x | x | x | x | x | x | x | x | |||||
| sul2 | x | x | |||||||||||||
| Tr | dfrA12 | x | x | x | x | x | x | x | x | ||||||
| dfrA14 | x | x | x | x | x | ||||||||||
| Te | tet(A) | x | |||||||||||||
| tet(D) | x | ||||||||||||||
| Bacterial Strain | mcrGene | mgrBGene | phoP Gene | phoQ Gene | pmrA Gene | pmrB Gene | crrA Gene | crrB Gene |
| BC1_TM | Negative * | Gene disruption by ISKpn25 insertion (nucleotides 130–144 deleted) | None | None | A217Vn | T246An | Negative | Negative |
| BC2_BM | Negative | M27Kd | L4Fd | Q426Ld | A217Vn | T246An | Negative | Negative |
| BC3_TM | Negative | None ** | None | L26Qd | None | T246An, R256Gd | None | C68Sn, Q296Ln |
| BC4_BM | Negative | M27Kd | None | Q426Ld | A217Vn | T246An | Negative | Negative |
| BC5_TM | Negative | M27Kd | None | None | A217Vn | T246An | Negative | Negative |
| BC6_BM | Negative | Gene disruption by ISKpn25 insertion (nucleotides 1–5 deleted) | None | None | None | T246An, R256Gd | None | C68Sn |
| BC7_BM | Negative | None | None | None | A41Tn | P95Ld, L213Mn, T246An | None | C68Sn |
| BC8_BM | Negative | None | None | None | None | T157Pd, T246An, R256Gd, V352Ed | None | C68Sn, Q296Ln |
| BC9_TM | Negative | Gene disruption by ISKpn26 insertion (nucleotides 74–144 deleted) | None | None | A217Vn | T246An | Negative | Negative |
| BC10_TM | Negative | M27Kd | None | Q426Ld | A217Vn | T246An | Negative | Negative |
| BC11_TM_B_hR | Negative | Gene disruption by ISKpn25 insertion (nucleotides 1–5 deleted) | None | L224Qd | None | T246An, R256Gd | None | C68Sn |
| BC12_TM_B_m | Negative | Gene disruption by ISKpn25 insertion (nucleotides 1–5 deleted) | None | L224Qd, Q317Kd | None | T246An, R256Gd | None | C68Sn |
| BC13_TM_C_hR | Negative | Gene disruption by ISKpn25 insertion (nucleotides 1–5 deleted) | None | L224Qd | None | T246An, R256Gd | None | C68Sn |
| BC14_TM_C_m | Negative | Gene disruption by ISKpn25 insertion (nucleotides 1–5 deleted) | None | L224Qd | None | T246An, R256Gd | None | C68Sn, P151Sd |
| Protein | Mutation | PROVEAN Score | Prediction |
|---|---|---|---|
| PhoP | L4F | −3.5 | Deleterious |
| PhoQ | Q426L | −3.6 | Deleterious |
| L26Q | −4.2 | Deleterious | |
| L224Q | −4.7 | Deleterious | |
| Q317K | −2.7 | Deleterious | |
| PmrB | R256G | −5.4 | Deleterious |
| P95L | −9.6 | Deleterious | |
| T157P | −5.7 | Deleterious | |
| V352E | −3.9 | Deleterious | |
| CrrB | P151S | −8 | Deleterious |
| Strain ID | Cp. Gene | Serotype | MLST Sequence Type | cgMLST Complex Type | Plasmid Replicons (% Identity) |
| BC1_TM | NDM-1 | KL17; O1v1 | ST101 | CT5839 | Col440II (99.29%), IncFIB(pQil) (100%), IncFII(K) (97.97%) |
| BC2_BM | OXA-48 | KL17; O1v1 | ST101 | CT5840 | ColRNAI (100%), IncFIA(HI1) (98.45%), IncFIB(K) (100%), IncFII(K) (97.97%), IncL (100%), IncR (100%) |
| BC3_TM | KPC-2 | KL106; O2v2 | ST258 | CT382 | ColRNAI (100%), IncFIB(K) (100%), IncFII(K) (97.97%), IncX3(100%) |
| BC4_BM | OXA-48 | KL17; O1v1 | ST101 | CT2059 | Col440II (99.29%), ColRNAI (100%), IncFIA(HI1) (98.45%), IncFIB(K) (100%), IncFII(K) (97.97%), IncL (100%), IncR (100%) |
| BC5_TM | KPC-2 | KL17; O1v1 | ST101 | CT5848 | Col440II (99.29%), ColRNAI (100%), ColpVC (98.45%), IncFIA(HI1) (98.45%), IncFIB(K) (100%), IncFII(K) (97.97%), IncR (100%) |
| BC6_BM | KPC-2 | KL112; O2v2 | ST147 | CT4408 | ColRNAI (100%), ColpVC (98.45%), IncFIA(HI1) (97.16%), IncFIB(K) (100%), IncFIB(pQil) (100%), IncFII(K) (97.97%) |
| BC7_BM | NDM-1 | KL102; O2v2 | ST307 | CT5853 | IncFIB(K) (98.93%), IncFII(Yp) (99.13%), IncC (100%) |
| BC8_BM | KPC-2 | KL106; O2v2 | ST258 | CT5854 | ColRNAI (100%), IncFIB(K) (100%), IncFII(K) (97.97%), IncX3 (100%) |
| BC9_TM | OXA-48 | KL17; O1v1 | ST101 | CT4633 | Col440II (99.29%), IncFIA(HI1) (98.45%), IncL (100%), IncR (100%) |
| BC10_TM | OXA-48 | KL17; O1v1 | ST101 | CT5840 | Col440II (99.29%), ColRNAI (100%), IncFIA(HI1) (98.45%), IncFIB(K) (100%), IncFII(K) (97.97%), IncL (100%), IncR (100%) |
| BC11_TM_B_hR | KPC-2 | KL112; O2v2 | ST147 | CT4408 | ColRNAI (100%), IncFIB(K) (100%), IncFIB (pQIL) (100%), IncFII(K) (97.97%) |
| BC12_TM_B_m | KPC-2 | KL112; O2v2 | ST147 | CT4408 | ColRNAI (100%), IncFIB(K) (100%), IncFIB (pQIL) (100%), IncFII(K) (100%) |
| BC13_TM_C_hR | KPC-2 | KL112; O2v2 | ST147 | CT4408 | ColRNAI (100%), ColpVC (98.45%), IncFIB(K) (100%), IncFIB (pQIL) (100%), IncFII(K) (97.97%) |
| BC14_TM_C_m | KPC-2 | KL112; O2v2 | ST147 | CT4408 | ColRNAI (100%), ColpVC (98.45%), IncFIB(K) (100%), IncFIB (pQIL) (100%), IncFII(K) (97.97%) |
| Bacterial Isolate | Cp. Type | Source | Date of Collection | Town of Isolation | Hospital Unit | Patient Gender | Patient Age (Years) |
|---|---|---|---|---|---|---|---|
| BC1_TM | MBL | Blood | 14.01.2019 | Târgu Mureș | Medical | M | 63 |
| BC2_BM | OXA-48-like | Wound | 30.07.2019 | Baia Mare | Medical | M | 79 |
| BC3_TM | KPC | Tracheal aspirate | 01.02.2018 | Târgu Mureș | ICU | M | 66 |
| BC4_BM | OXA-48-like | Urine | 11.06.2018 | Baia Mare | Surgical | M | 70 |
| BC5_TM | KPC | Wound | 09.08.2018 | Târgu Mureș | ICU | M | 26 |
| BC6_BM | KPC | Urine | 23.06.2020 | Baia Mare | Medical | M | 75 |
| BC7_BM | MBL | Urine | 23.04.2021 | Baia Mare | Surgical | F | 68 |
| BC8_BM | KPC | Blood | 21.11.2017 | Baia Mare | Surgical | M | 67 |
| BC9_TM | OXA-48-like | Blood | 28.01.2019 | Târgu Mureș | ICU | F | 91 |
| BC10_TM | OXA-48-like | Tracheal aspirate | 08.01.2019 | Târgu Mureș | ICU | M | 68 |
| BC11_TM_B_hR | KPC | Tracheal aspirate | 28.03.2019 | Târgu Mureș | ICU | M | 45 |
| BC12_TM_B_m | KPC | - | - | - | - | - | - |
| BC13_TM_C_hR | KPC | Tracheal aspirate | 04.03.2019 | Târgu Mureș | ICU | F | 64 |
| BC14_TM_C_m | KPC | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Főldes, A.; Oprea, M.; Székely, E.; Usein, C.-R.; Dobreanu, M. Characterization of Carbapenemase-Producing Klebsiella pneumoniae Isolates from Two Romanian Hospitals Co-Presenting Resistance and Heteroresistance to Colistin. Antibiotics 2022, 11, 1171. https://doi.org/10.3390/antibiotics11091171
Főldes A, Oprea M, Székely E, Usein C-R, Dobreanu M. Characterization of Carbapenemase-Producing Klebsiella pneumoniae Isolates from Two Romanian Hospitals Co-Presenting Resistance and Heteroresistance to Colistin. Antibiotics. 2022; 11(9):1171. https://doi.org/10.3390/antibiotics11091171
Chicago/Turabian StyleFőldes, Annamária, Mihaela Oprea, Edit Székely, Codruța-Romanița Usein, and Minodora Dobreanu. 2022. "Characterization of Carbapenemase-Producing Klebsiella pneumoniae Isolates from Two Romanian Hospitals Co-Presenting Resistance and Heteroresistance to Colistin" Antibiotics 11, no. 9: 1171. https://doi.org/10.3390/antibiotics11091171