
Characteristics of Antibiotic Resistance and Tolerance of Environmentally Endemic Pseudomonas aeruginosa

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials for Culture Medium
2.2. Isolation of Strains from the Environment
2.3. Isolation of P. aeruginosa from Clinics
2.4. Antibiotic Susceptibility Test for Planktonic Bacteria
2.5. Carbapenem Susceptibility Test for Adherent Bacteria
2.6. Genotype Analysis by PCR-Based ORF Typing (POT)
2.7. Evaluation of Resistance Risk
3. Results
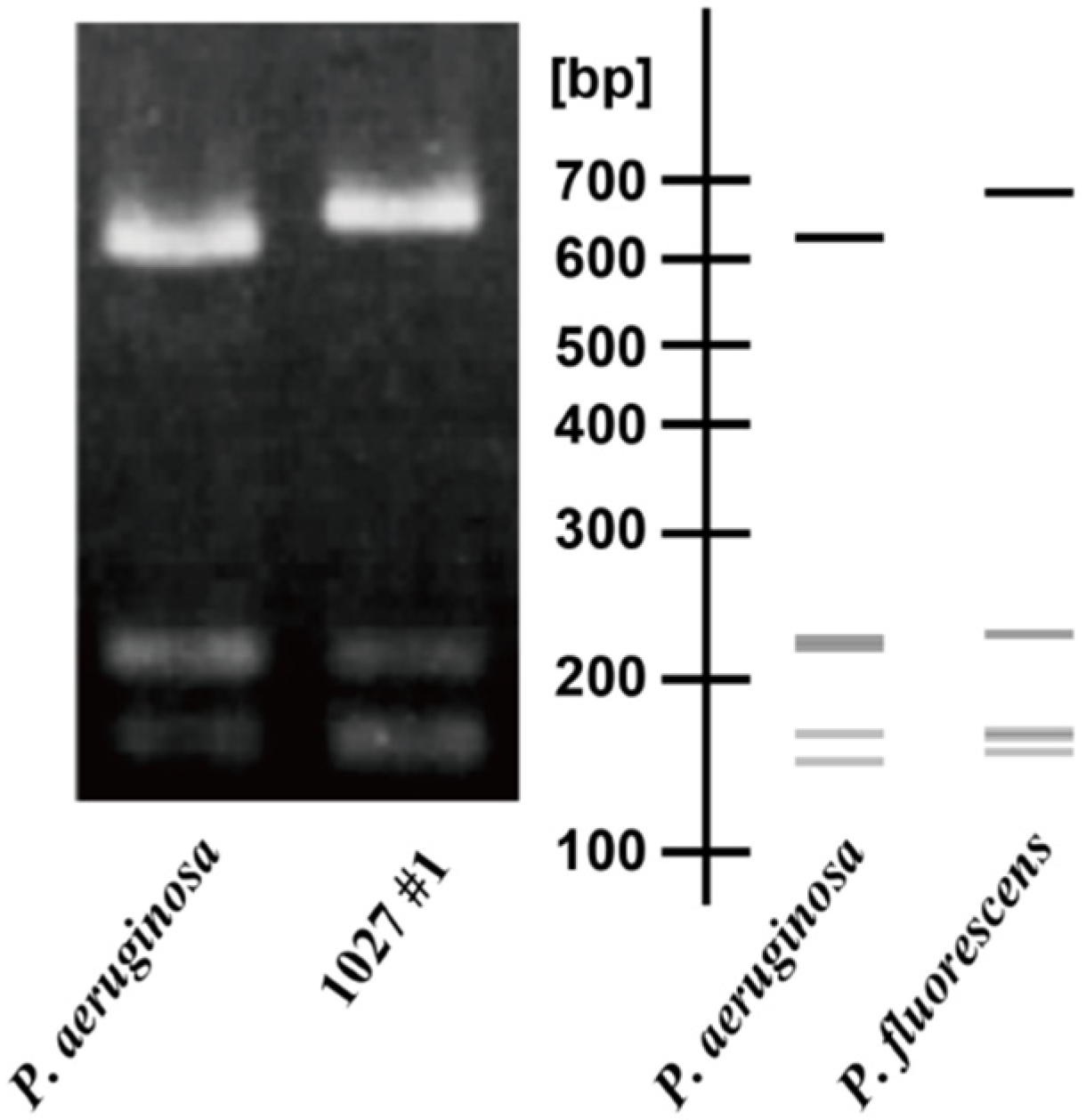
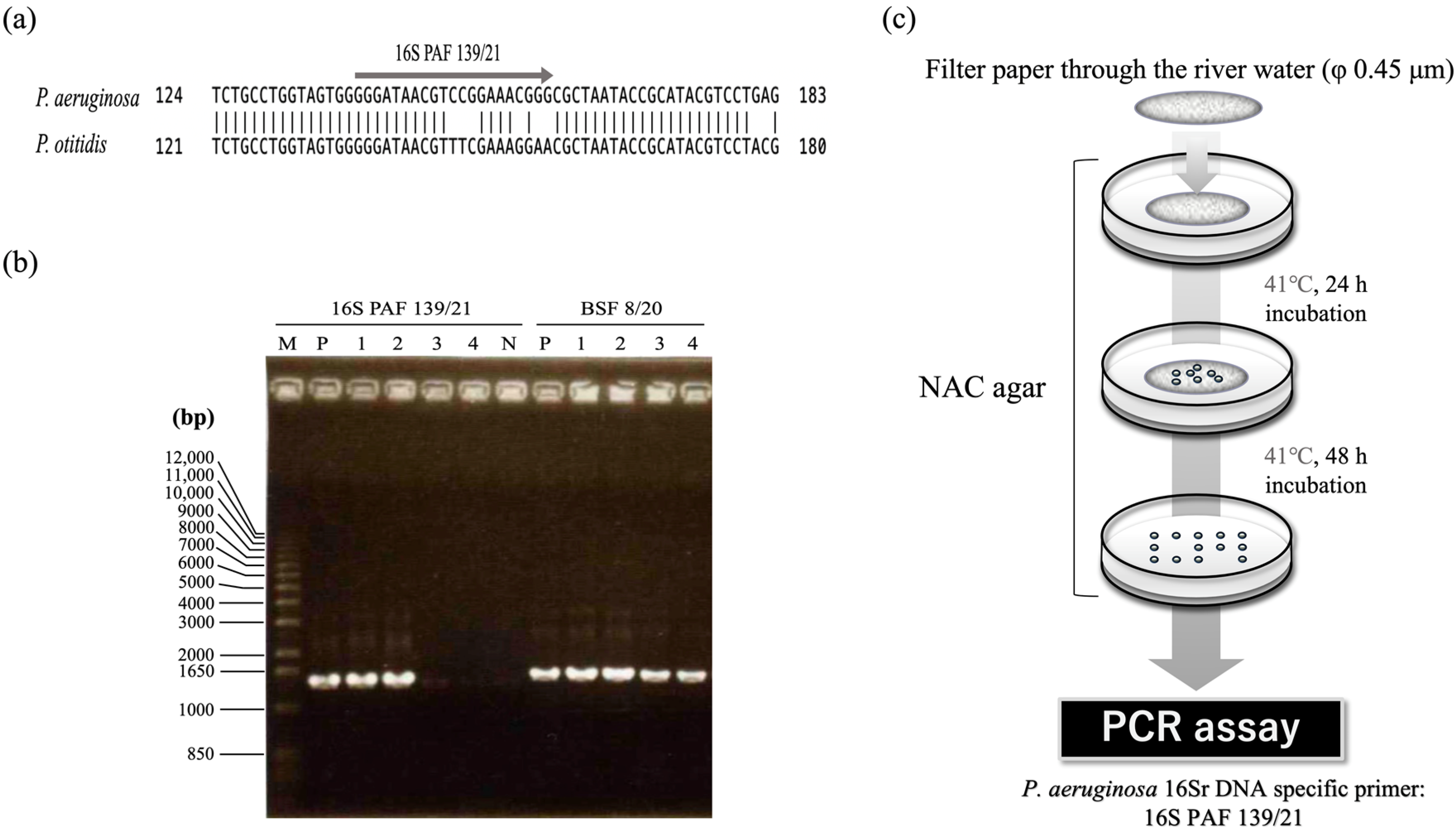
3.1. Construction of the Method for Isolating P. aeruginosa Cells from Environments
3.2. Isolation of P. aeruginosa from River Water
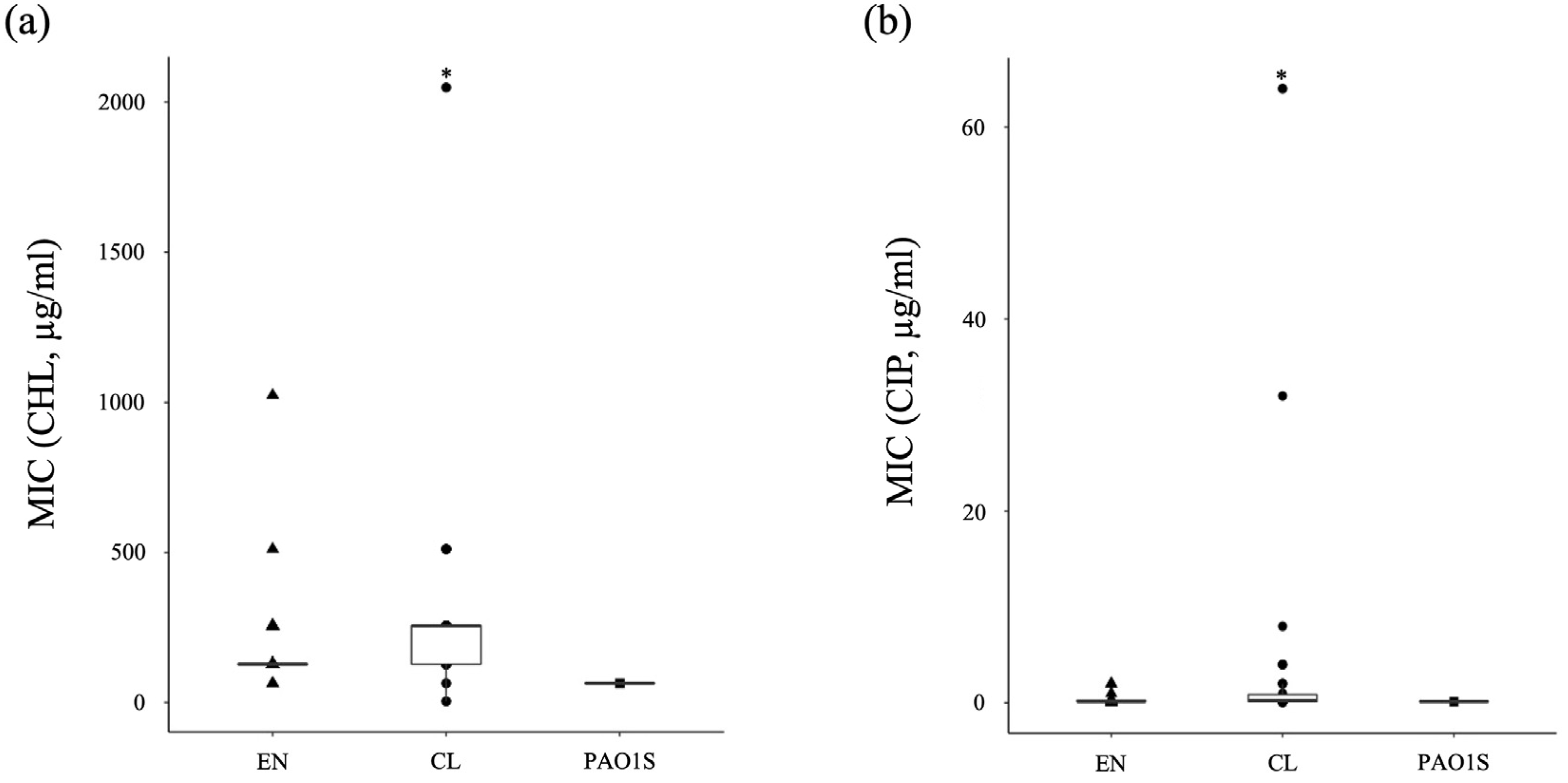
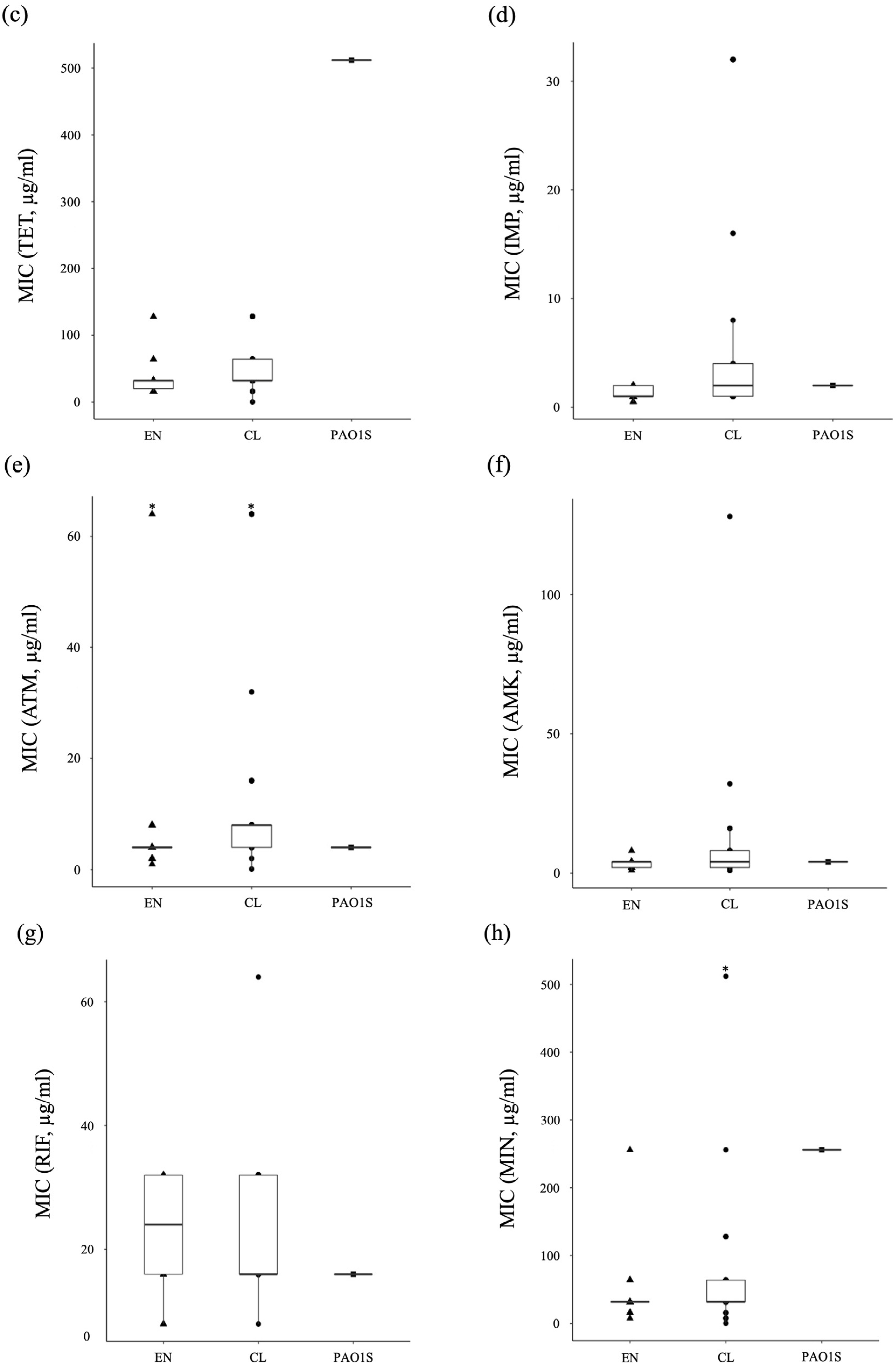
3.3. The Ratio of Antibiotic-Resistant Strains of P. aeruginosa in the Environment and Clinical Setting
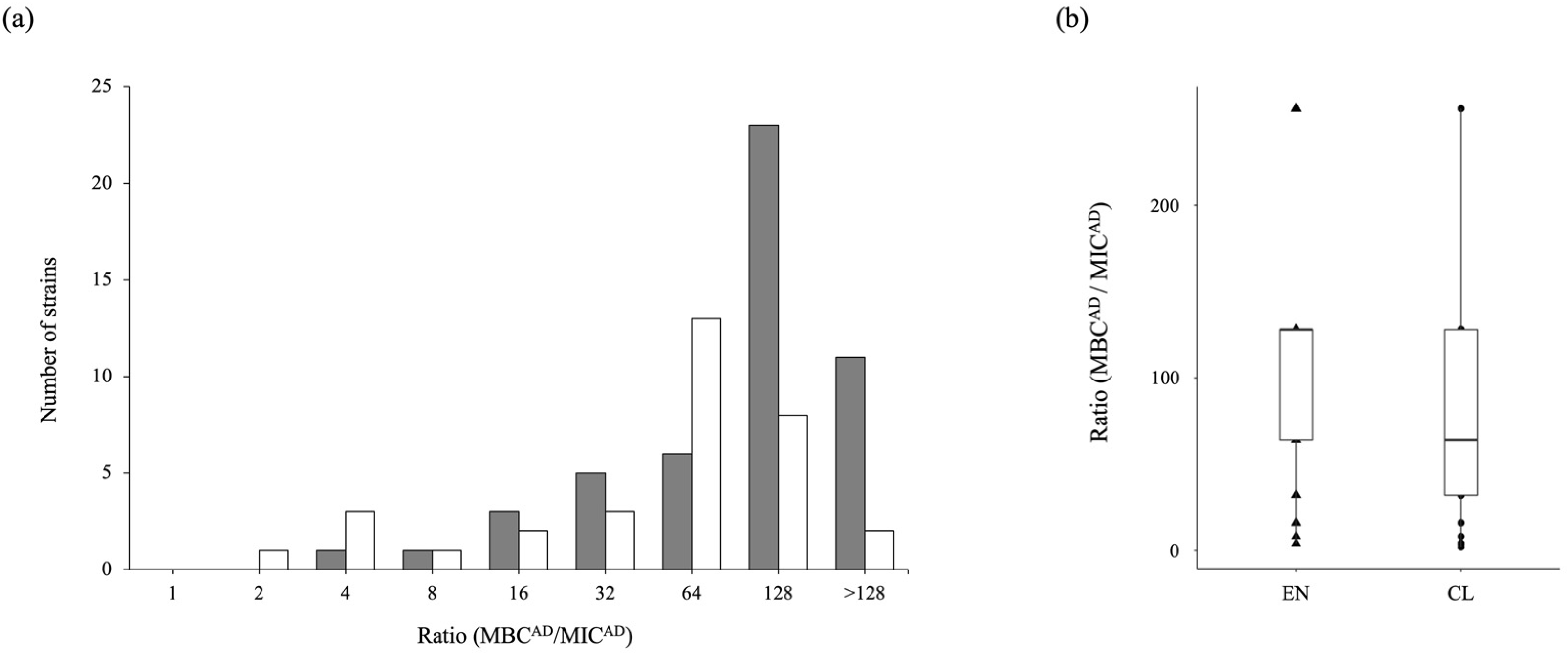
3.4. Antibiotic-Tolerance Level of P. aeruginosa in the Environment and Clinical Setting
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Neill, J. Antimicrobials in Agriculture and the Environment: Reducing Unnecessary Use and Waste. In The Review on Antimicrobial Resistance (AMR). Available online: https://amr-review.org/sites/default/files/Antimicrobials%20in%20agriculture%20and%20the%20environment%20-%20Reducing%20unnecessary%20use%20and%20waste.pdf (accessed on 14 June 2022).
- World Health Organization. No Time to Wait: Securing the Future from Drug-Resistant Infections, Report to the Secretary-General of the United Nations. Available online: https://www.who.int/publications/i/item/no-time-to-wait-securing-the-future-from-drug-resistant-infections (accessed on 16 June 2022).
- Fillol-Salom, A.; Alsaadi, A.; Sousa, J.A.M.D.; Zhong, L.; Foster, K.R.; Rocha, E.P.C.; Penadés, J.R.; Ingmer, H.; Haaber, J. Bacteriophages Benefit from Generalized Transduction. PLoS Pathog. 2019, 15, e1007888. [Google Scholar] [CrossRef] [PubMed]
- Headd, B.; Bradford, S.A. The Conjugation window in an Escherichia coli K-12 Strain with an IncFII Plasmid. Appl. Environ. Microbiol. 2020, 86, e00948-20. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, E.; Muth, G.; Espinosa, M. Conjugative plasmid transfer in gram-positive bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 277–301. [Google Scholar] [CrossRef] [PubMed]
- Riva, F.; Riva, V.; Eckert, E.M.; Colinas, N.; Di Cesare, A.; Borin, S.; Mapelli, F.; Crotti, E. An Environmental Escherichia coli strain is naturally competent to acquire exogenous DNA. Front. Microbiol. 2020, 11, 574301. [Google Scholar] [CrossRef] [PubMed]
- Dell’Annunziata, F.; Dell’Aversana, C.; Doti, N.; Donadio, G.; Dal Piaz, F.; Izzo, V.; De Filippis, A.; Galdiero, M.; Altucci, L.; Boccia, G.; et al. Outer membrane vesicles derived from Klebsiella pneumoniae are a driving force for horizontal gene transfer. Int. J. Mol. Sci. 2021, 22, 8732. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Thirumalmuthu, K.; Devarajan, B.; Prajna, L.; Mohankumar, V. Mechanisms of fluoroquinolone and aminoglycoside resistance in keratitis-associated Pseudomonas aeruginosa. Microb. Drug Resist. 2019, 25, 813–823. [Google Scholar] [CrossRef]
- Sindeldecker, D.; Stoodley, P. The Many antibiotic resistance and tolerance strategies of Pseudomonas aeruginosa. Biofilm 2021, 3, 100056. [Google Scholar] [CrossRef]
- Nakae, T.; Yoshihara, E.; Yoneyama, H. Multiantibiotic resistance caused by active drug extrusion in hospital pathogens. J. Infect. Chemother. 1997, 3, 173–183. [Google Scholar] [CrossRef]
- Maseda, H.; Yoneyama, H.; Nakae, T. Assignment of the substrate-selective subunits of the MexEF-OprN multidrug efflux pump of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 658–664. [Google Scholar] [CrossRef]
- Murray, J.L.; Kwon, T.; Marcotte, E.M.; Whiteley, M. Intrinsic antimicrobial resistance determinants in the superbug Pseudomonas aeruginosa. Mbio 2015, 6, e01603-15. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; Braniss, I.; Shoresh, N.; Balaban, N.Q. Antibiotic tolerance facilitates the evolution of resistance. Science 2017, 355, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Keren, I.; Kaldalu, N.; Spoering, A.; Wang, Y.; Lewis, K. Persister cells and tolerance to antimicrobials. FEMS Microbiol. Lett. 2004, 230, 13–18. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells and the riddle of biofilm survival. Biochemistry 2005, 70, 267–274. [Google Scholar] [CrossRef]
- Soares, A.; Alexandre, K.; Etienne, M. Tolerance and persistence of Pseudomonas aeruginosa in biofilms exposed to antibiotics: Molecular mechanisms, antibiotic strategies and therapeutic perspectives. Front. Microbiol. 2020, 11, 2057. [Google Scholar] [CrossRef]
- Bigger, J. Treatment of staphylococcal infections with penicillin by intermittent sterilisation. Lancet 1944, 244, 497–500. [Google Scholar] [CrossRef]
- Dore, M.P.; Osato, M.S.; Realdi, G.; Mura, I.; Graham, D.Y.; Sepulveda, A.R. Amoxycillin tolerance in Helicobacter Pylori. J. Antimicrob. Chemother. 1999, 43, 47–54. [Google Scholar] [CrossRef]
- Perry, J.D.; Jones, A.L.; Gould, F.K. Glycopeptide tolerance in bacteria causing endocarditis. J. Antimicrob. Chemother. 1999, 44, 121–124. [Google Scholar] [CrossRef]
- Zelenitsky, S. Antibiotic tolerance of peritoneal bacterial isolates in dialysis fluids. J. Antimicrob. Chemother. 2002, 49, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Azuma, M.; Murakami, K.; Murata, R.; Kataoka, K.; Fujii, H.; Miyake, Y.; Nishioka, Y. Clinical significance of carbapenem-tolerant Pseudomonas aeruginosa isolated in the respiratory tract. Antibiotics 2020, 9, 626. [Google Scholar] [CrossRef] [PubMed]
- Maseda, H.; Saito, K.; Nakajima, A.; Nakae, T. Variation of the mexT gene, a regulator of the MexEF-OprN efflux pump expression in wild-type strains of Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2000, 192, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Ichise, Y.-K.; Kosuge, T.; Uwate, M.; Nakae, T.; Maseda, H. Complete genome sequence of Pseudomonas aeruginosa strain 8380, isolated from the human gut. Genome Announc. 2015, 3, e00520-15. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Yamada, K.; Aoki, M.; Hosoba, E.; Matsumoto, M.; Baba, H.; Iinuma, Y. Applying a PCR-based open-reading frame typing method for easy genotyping and molecular epidemiological analysis of Pseudomonas aeruginosa. J. Appl. Microbiol. 2016, 120, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Lilly, H.A.; Lowbury, E.J. Cetrimide-nalidixic acid agar as a selective medium for Pseudomonas aeruginosa. J. Med. Microbiol. 1972, 5, 151–153. [Google Scholar] [CrossRef]
- Widmer, F.; Seidler, R.J.; Gillevet, P.M.; Watrud, L.S.; Di Giovanni, G.D. A Highly selective PCR protocol for detecting 16S rRNA genes of the genus Pseudomonas (Sensu Stricto) in environmental samples. Appl. Environ. Microbiol. 1998, 64, 2545–2553. [Google Scholar] [CrossRef]
- Brenner, E.J.; Krieg, N.R.; Staley, J.T. Bergey’s Manual of Systematic Bacteriology: Volume Two, 2nd ed.; The Proteobacteria, Part C; Springer: New York, NY, USA, 2005; pp. 582–583. [Google Scholar]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Second Informational Supplement. In CLSI Document M100-S22; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Miyake, Y.; Fujiwara, S.; Usui, T.; Suginaka, H. Simple method for measuring the antibiotic concentration required to kill adherent bacteria. Chemotherapy 1992, 38, 286–290. [Google Scholar] [CrossRef]
- Aaron, S.D.; Ferris, W.; Ramotar, K.; Vandemheen, K.; Chan, F.; Saginur, R. Single and combination antibiotic susceptibilities of planktonic, adherent, and biofilm-grown Pseudomonas aeruginosa isolates cultured from sputa of adults with cystic fibrosis. J. Clin. Microbiol. 2002, 40, 4172–4179. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Ono, T.; Viducic, D.; Somiya, Y.; Kariyama, R.; Hori, K.; Amoh, T.; Hirota, K.; Kumon, H.; Parsek, M.R.; et al. Role of psl Genes in Antibiotic Tolerance of Adherent Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2017, 61, e02587-16. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Cantón, R.; Campo, P.; Baquero, F.; Blázquez, J. High frequency of hypermutable Pseudomonas aeruginosa in cystic fibrosis lung infection. Science 2000, 288, 1251–1254. [Google Scholar] [CrossRef]
- WHO. Implementation Manual to Prevent and Control the Spread of Carbapenem-Resistant Organisms at the National and Health Care Facility Level: Interim Practical Manual Supporting Implementation of the Guidelines for the Prevention and Control of Carbapenem-Resistant Enterobacteriaceae, Acinetobacter Baumannii and Pseudomonas Aeruginosa in Health Care Facilities; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Centers for Disease Control and Prevention (U.S.); National Center for Emerging Zoonotic and Infectious Diseases (U.S.); Division of Healthcare Quality Promotion. Antibiotic Resistance Coordination and Strategy Unit. In Antibiotic Resistance Threats in the United States. Atlanta, GA, USA; 2019. Available online: https://stacks.cdc.gov/view/cdc/82532 (accessed on 26 April 2020).
- Larsson, D.G.J.; Flach, C.-F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and resistance of Pseudomonas aeruginosa biofilms to antimicrobial agents—How P. aeruginosa can escape antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Masai, S.; Murakami, K.; Azuma, M.; Kataoka, K.; Sebe, M.; Shimizu, K.; Itayama, T.; Whangchai, N.; Whangchai, K.; et al. Characteristics of Antibiotic Resistance and Tolerance of Environmentally Endemic Pseudomonas aeruginosa. Antibiotics 2022, 11, 1120. https://doi.org/10.3390/antibiotics11081120
Kim S, Masai S, Murakami K, Azuma M, Kataoka K, Sebe M, Shimizu K, Itayama T, Whangchai N, Whangchai K, et al. Characteristics of Antibiotic Resistance and Tolerance of Environmentally Endemic Pseudomonas aeruginosa. Antibiotics. 2022; 11(8):1120. https://doi.org/10.3390/antibiotics11081120
Chicago/Turabian StyleKim, Seryoung, Satomi Masai, Keiji Murakami, Momoyo Azuma, Keiko Kataoka, Mayu Sebe, Kazuya Shimizu, Tomoaki Itayama, Niwooti Whangchai, Kanda Whangchai, and et al. 2022. "Characteristics of Antibiotic Resistance and Tolerance of Environmentally Endemic Pseudomonas aeruginosa" Antibiotics 11, no. 8: 1120. https://doi.org/10.3390/antibiotics11081120