
Molecular Epidemiology of Antimicrobial Resistance, Virulence and Capsular Serotypes of Carbapenemase-Carrying Klebsiella pneumoniae in China

Abstract
:1. Introduction
2. Results
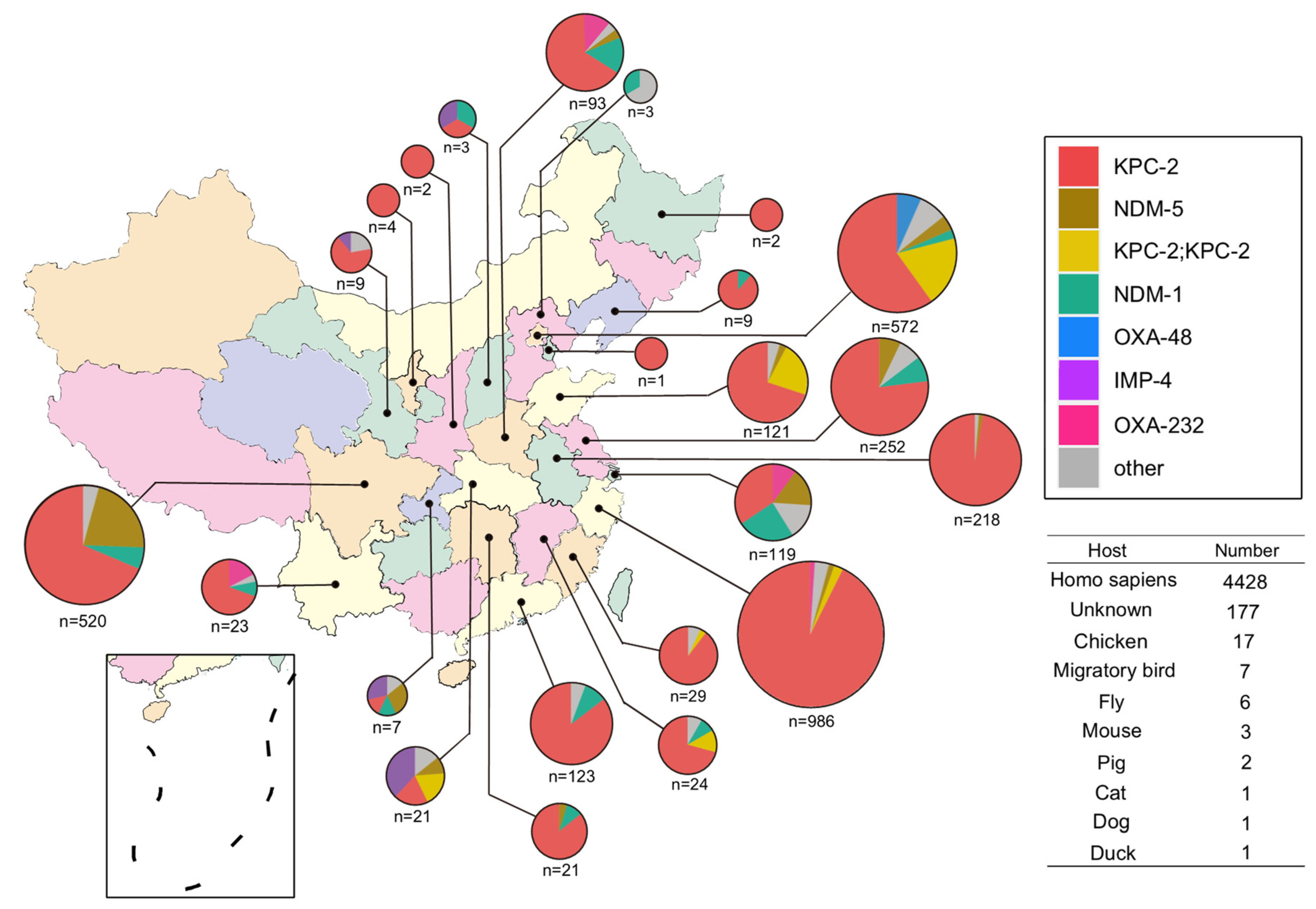
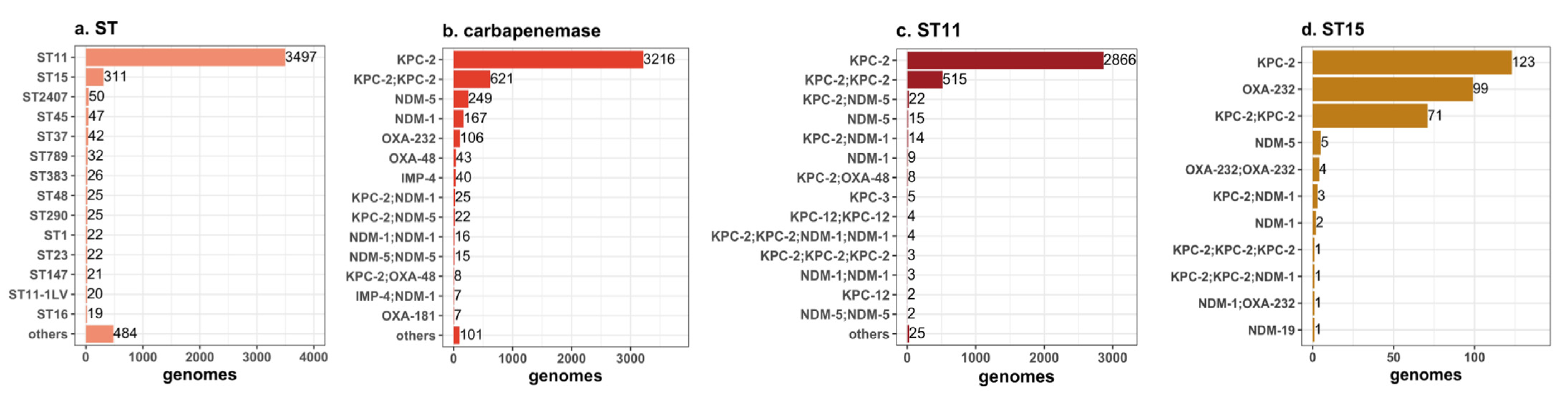
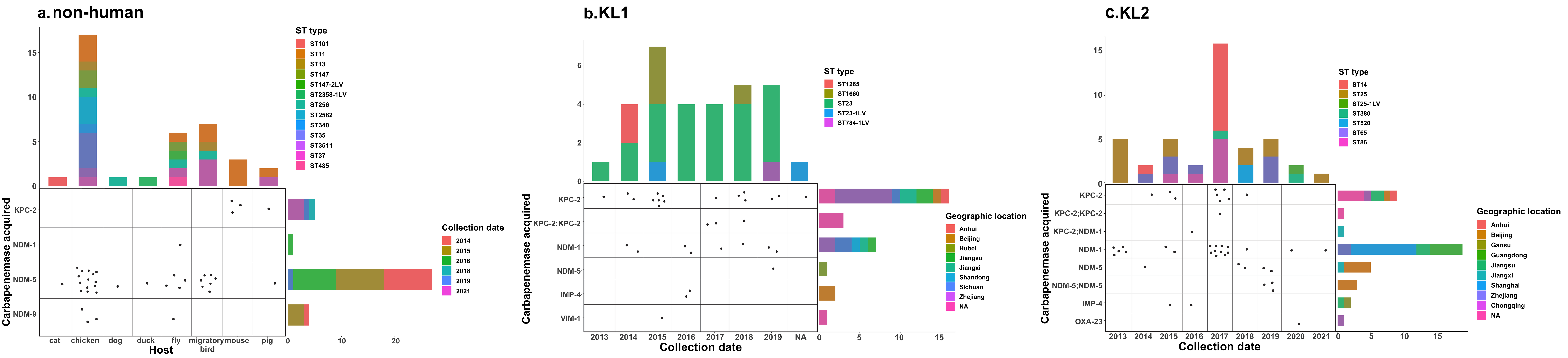
2.1. Antimicrobial Resistance
2.2. Virulence
2.2.1. Aerobactin
2.2.2. Colibactin
2.2.3. Salmochelin
2.2.4. Yersiniabactin
2.2.5. RmpADC
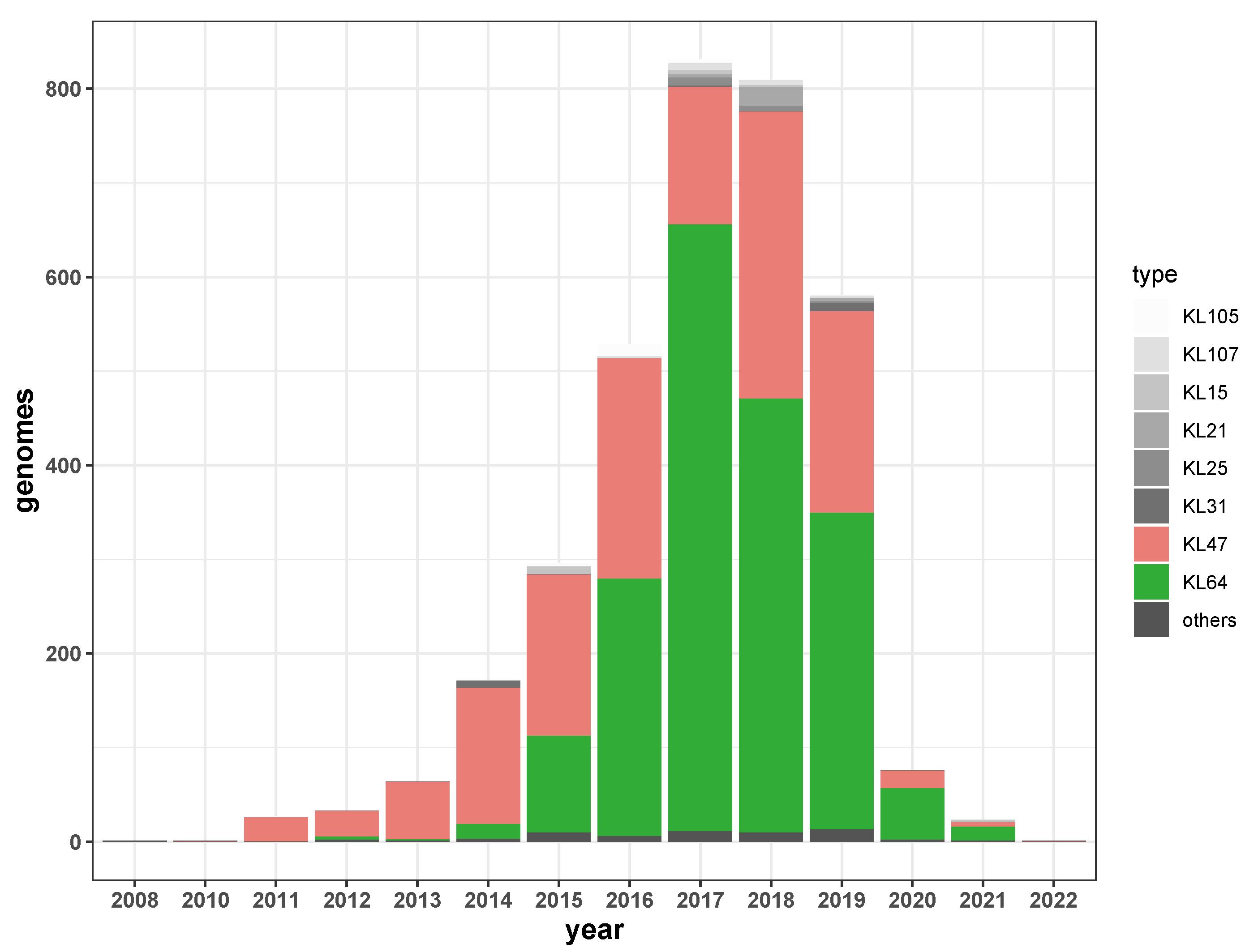
2.3. Serotype
3. Discussion
4. Materials and Methods
4.1. Klebsiella pneumoniae Genome-Wide Data
4.2. Kleborate Analysis
4.3. BioSample Information Collection
4.4. Comparative Analysis of Virulence Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wyres, K.L.; Lam, M.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Gorrie, C.L.; Mirčeta, M.; Wick, R.R.; Edwards, D.J.; Thomson, N.R.; Strugnell, R.A.; Pratt, N.F.; Garlick, J.S.; Watson, K.M.; Pilcher, D.V. Gastrointestinal carriage is a major reservoir of Klebsiella pneumoniae infection in intensive care patients. Clin. Infect. Dis. 2017, 65, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.N. Microbial etiologies of hospital-acquired bacterial pneumonia and ventilator-associated bacterial pneumonia. Clin. Infect. Dis. 2010, 51 (Suppl. 1), S81–S87. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.-J.; Sun, B.; Ding, X.; Li, H.; Feng, X. Changing trends in the bacteriological profiles and antibiotic susceptibility in neonatal sepsis at a tertiary children’s hospital of China. Transl. Pediatrics 2020, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Shon, A.S.; Bajwa RP, S.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae as a key trafficker of drug resistance genes from environmental to clinically important bacteria. Curr. Opin. Microbiol. 2018, 45, 131–139. [Google Scholar] [CrossRef]
- Guh, A.Y.; Bulens, S.N.; Mu, Y.; Jacob, J.T.; Reno, J.; Scott, J.; Wilson, L.E.; Vaeth, E.; Lynfield, R.; Shaw, K.M. Epidemiology of carbapenem-resistant Enterobacteriaceae in 7 US communities, 2012–2013. JAMA 2015, 314, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, H.; Glasner, C.; Albiger, B.; Aanensen, D.M.; Tomlinson, C.T.; Andrasević, A.T.; Cantón, R.; Carmeli, Y.; Friedrich, A.W.; Giske, C.G.; et al. Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): A prospective, multinational study. Lancet Infect. Dis. 2017, 17, 153–163. [Google Scholar] [CrossRef]
- Brisse, S.; Fevre, C.; Passet, V.; Issenhuth-Jeanjean, S.; Tournebize, R.; Diancourt, L.; Grimont, P. Virulent clones of Klebsiella pneumoniae: Identification and evolutionary scenario based on genomic and phenotypic characterization. PLoS ONE 2009, 4, e4982. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, Y.; Ye, L.; Yang, J. Molecular epidemiology and virulence factors of pyogenic liver abscess causing Klebsiella pneumoniae in China. Clin. Microbiol. Infect. 2014, 20, O818–O824. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.-H.; Huang, Y.T.; Chang, C.Y.; Hsu, H.S.; Hsueh, P.-R. Capsular serotypes and multilocus sequence types of bacteremic Klebsiella pneumoniae isolates associated with different types of infections. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.-S.; Garin, B.; Le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.-H.; et al. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg. Infect. Dis. 2014, 20, 1812–1820. [Google Scholar] [CrossRef]
- Struve, C.; Roe, C.C.; Stegger, M.; Stahlhut, S.G.; Hansen, D.S.; Engelthaler, D.M.; Andersen, P.S.; Driebe, E.M.; Keim, P.; Krogfelt, K.A. Mapping the evolution of hypervirulent Klebsiella pneumoniae. MBio 2015, 6, e00630-e15. [Google Scholar] [CrossRef] [PubMed]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef]
- Lam, M.M.C.; Wyres, K.L.; Judd, L.M.; Wick, R.R.; Jenney, A.; Brisse, S.; Holt, K.E. Tracking key virulence loci encoding aerobactin and salmochelin siderophore synthesis in Klebsiella pneumoniae. Genome Med. 2018, 10, 77. [Google Scholar] [CrossRef]
- Lam, M.M.C.; Wyres, K.L.; Judd, L.M.; Wick, R.R.; Jenney, A.; Brisse, S.; Holt, K.E. Genetic diversity, mobilisation and spread of the yersiniabactin-encoding mobile element ICEKp in Klebsiella pneumoniae populations. Microb. Genom. 2018, 4, e000196. [Google Scholar] [CrossRef]
- Walker, K.A.; Miner, T.A.; Palacios, M.; Trzilova, D.; Frederick, D.R.; Broberg, C.; Sepúlveda, V.E.; Quinn, J.; Miller, V.L. A Klebsiella pneumoniae regulatory mutant has reduced capsule expression but retains hypermucoviscosity. MBio 2019, 10, e00089-e19. [Google Scholar] [CrossRef]
- Walker, K.A.; Treat, L.P.; Sepúlveda, V.E.; Miller, V.L. The small protein RmpD drives hypermucoviscosity in Klebsiella pneumoniae. MBio 2020, 11, e01750-e20. [Google Scholar] [CrossRef]
- Zhang, R.; Lin, D.; Chan, E.W.-C.; Gu, D.; Chen, G.-X.; Chen, S. Emergence of carbapenem-resistant serotype K1 hypervirulent Klebsiella pneumoniae strains in China. Antimicrob. Agents Chemother. 2016, 60, 709–711. [Google Scholar] [CrossRef]
- Gu, D.; Dong, N.; Zheng, Z.; Lin, D.; Huang, M.; Wang, L.; Chan, E.W.-C.; Shu, L.; Yu, J.; Zhang, R.; et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: A molecular epidemiological study. Lancet Infect. Dis. 2018, 18, 37–46. [Google Scholar] [CrossRef]
- Gonzalez-Ferrer, S.; Peñaloza, H.F.; Budnick, J.A.; Bain, W.G.; Nordstrom, H.R.; Lee, J.S.; Van Tyne, D. Finding order in the chaos: Outstanding questions in Klebsiella pneumoniae pathogenesis. Infect. Immun. 2021, 89, e00693-e20. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Wick, R.R.; Judd, L.M.; Froumine, R.; Tokolyi, A.; Gorrie, C.L.; Lam, M.M.C.; Duchêne, S.; Jenney, A.; Holt, K.E. Distinct evolutionary dynamics of horizontal gene transfer in drug resistant and virulent clones of Klebsiella pneumoniae. PLoS Genet. 2019, 15, e1008114. [Google Scholar] [CrossRef]
- Zhou, K.; Xiao, T.; David, S.; Wang, Q.; Zhou, Y.; Guo, L.; Aanensen, D.; Holt, K.E.; Thomson, N.R.; Grundmann, H.; et al. Novel subclone of carbapenem-resistant Klebsiella pneumoniae sequence type 11 with enhanced virulence and transmissibility, China. Emerg. Infect. Dis. 2020, 26, 289. [Google Scholar] [CrossRef]
- El-Ashry, A.H.; Hendawy, S.R.; Mahmoud, N.M. Prevalence of pks genotoxin among hospital-acquired Klebsiella pneumoniae. AIMS Microbiol. 2022, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-C.; Lin, A.-C.; Chiang, M.-K.; Dai, Y.-H.; Hsu, C.-C.; Lu, M.-C.; Liau, C.-Y.; Chen, Y.-T. Genotoxic Klebsiella pneumoniae in Taiwan. PLoS ONE 2014, 9, e96292. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Olson, R.; MacDonald, U.; Metzger, D.; Maltese, L.M.; Drake, E.J.; Gulick, A. Aerobactin mediates virulence and accounts for increased siderophore production under iron-limiting conditions by hypervirulent (hypermucoviscous) Klebsiella pneumoniae. Infect. Immun. 2014, 82, 2356–2367. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Olson, R.; MacDonald, U.; Beanan, J.; Davidson, B.A. Aerobactin, but not yersiniabactin, salmochelin, or enterobactin, enables the growth/survival of hypervirulent (hypermucoviscous) Klebsiella pneumoniae Ex Vivo and In Vivo. Infect. Immun. 2015, 83, 3325–3333. [Google Scholar] [CrossRef]
- Sugawara, E.; Kojima, S.; Nikaido, H. Klebsiella pneumoniae major porins OmpK35 and OmpK36 allow more efficient diffusion of β-lactams than their Escherichia coli homologs OmpF and OmpC. J. Bacteriol. 2016, 198, 3200–3208. [Google Scholar] [CrossRef]
- Wong, J.L.C.; Romano, M.; Kerry, L.E.; Kwong, H.-S.; Low, W.-W.; Brett, S.J.; Clements, A.; Beis, K.; Frankel, G. OmpK36-mediated carbapenem resistance attenuates ST258 Klebsiella pneumoniae in vivo. Nat. Commun. 2019, 10, 3957. [Google Scholar] [CrossRef]
- Tsai, Y.-K.; Fung, C.-P.; Lin, J.-C.; Chen, J.-H.; Chang, F.-Y.; Chen, T.-L.; Siu, L.K. Klebsiella pneumoniae outer membrane porins OmpK35 and OmpK36 play roles in both antimicrobial resistance and virulence. Antimicrob. Agents Chemother. 2011, 55, 1485–1493. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.M.C.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.E. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat. Commun. 2021, 12, 4188. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Aerobactin (iuc Lineage) | Colibactin (clb Lineage) | Salmochelin (iro Lineage) | Yersiniabactin (ybt Lineage) | RmpADC (rmp Lineage) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Type | No. | Prevalence | Type | No. | Prevalence | Type | No. | Prevalence | Type | No. | Prevalence | Type | No. | Prevalence | |
| KL1 (270) | iuc-1 | 242 | 89.6% | clb 2 | 226 | 83.7% | iro 1 | 234 | 86.6% | ybt1; ICEKp10 | 239 | 88.5% | rmp1; KpVP-1 | 225 | 83.3% |
| ESBL− a;Carb− b | - | 20 | 7.4% | - | 28 | 10.3% | - | 17 | 6.2% | ybt2; ICEKp1 | 17 | 6.2% | - | 24 | 8.8% |
| rmp3; ICEKp1 | 14 | 5.1% | |||||||||||||
| KL1 (31) | iuc-1 | 28 | 90.3% | clb 2 | 30 | 96.7% | iro 1 | 27 | 87.1% | ybt1; ICEKp10 | 30 | 96.7% | rmp1; KpVP-1 | 25 | 80.6% |
| Carb+ c | - | 3 | 9.7% | - | 1 | 3.2% | - | 4 | 12.9% | - | 4 | 12.9% | |||
| KL2 (129) | iuc-1 | 98 | 75.9% | - | 84 | 65.1% | iro 1 | 101 | 78.2% | - | 57 | 44.1% | rmp1; KpVP-1 | 95 | 73.6% |
| ESBL−; Carb− | - | 17 | 13.1% | clb 3 | 35 | 27.1% | - | 14 | 10.8% | ybt17; ICEKp10 | 36 | 27.9% | - | 16 | 12.4% |
| iuc-2 | 9 | 6.9% | clb 1 | 8 | 6.2% | iro 2 | 9 | 6.9% | ybt9; ICEKp3 | 9 | 6.9% | rmp2; KpVP-2 | 9 | 6.9% | |
| KL2 (42) | - | 21 | 50.0% | - | 36 | 85.7% | - | 26 | 61.9% | - | 16 | 38.0% | - | 26 | 61.9% |
| Carb+ | iuc-1 | 11 | 26.1% | clb 3 | 5 | 11.9% | iro 1 | 13 | 30.9% | ybt16; ICEKp12 | 10 | 23.8% | rmp1; KpVP-1 | 13 | 30.9% |
| iuc-3 | 7 | 16.6% | ybt17; ICEKp10 | 5 | 11.9% | ||||||||||
| CKP (366) | - | 302 | 82.5% | - | 350 | 95.6% | - | 318 | 86.8% | - | 237 | 64.7% | - | 326 | 89.0% |
| ESBL−; Carb− | iuc-1 | 31 | 8.4% | iro 1 | 32 | 8.7% | ybt9; ICEKp3 | 33 | 9.0% | rmp1; KpVP-1 | 25 | 6.8% | |||
| iuc-3 | 13 | 3.5% | ybt10; ICEKp4 | 20 | 5.4% | ||||||||||
| ST11-CRKP (3497) | - | 1764 | 50.4% | - | 3496 | 99.9% | - | 3302 | 94.4% | ybt9; ICEKp3 | 3400 | 97.2% | - | 2460 | 70.3% |
| Carb+ | iuc-1 | 1694 | 48.4% | - | 38 | 1.0% | rmp1; KpVP-1 | 936 | 26.7% | ||||||
| ST15-CRKP (311) | - | 174 | 55.9% | - | 311 | 100.0% | - | 310 | 99.6% | ybt14; ICEKp5 | 144 | 46.3% | - | 311 | 100.0% |
| Carb+ | iuc-1 | 129 | 41.4% | ybt16; ICEKp12 | 102 | 32.7% | |||||||||
| ybt14; ICEKp12 | 30 | 9.6% | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Xia, X.; Yuan, T.; Zhu, J.; Shen, Z.; Li, M. Molecular Epidemiology of Antimicrobial Resistance, Virulence and Capsular Serotypes of Carbapenemase-Carrying Klebsiella pneumoniae in China. Antibiotics 2022, 11, 1100. https://doi.org/10.3390/antibiotics11081100
Zhao L, Xia X, Yuan T, Zhu J, Shen Z, Li M. Molecular Epidemiology of Antimicrobial Resistance, Virulence and Capsular Serotypes of Carbapenemase-Carrying Klebsiella pneumoniae in China. Antibiotics. 2022; 11(8):1100. https://doi.org/10.3390/antibiotics11081100
Chicago/Turabian StyleZhao, Lina, Xinxin Xia, Ting Yuan, Junying Zhu, Zhen Shen, and Min Li. 2022. "Molecular Epidemiology of Antimicrobial Resistance, Virulence and Capsular Serotypes of Carbapenemase-Carrying Klebsiella pneumoniae in China" Antibiotics 11, no. 8: 1100. https://doi.org/10.3390/antibiotics11081100