
Effects of Sodium Hexametaphosphate and Fluoride on the pH and Inorganic Components of Streptococcus mutans and Candida albicans Biofilm after Sucrose Exposure

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Evaluation of the Number of Cells Prior to the Treatment of Biofilms
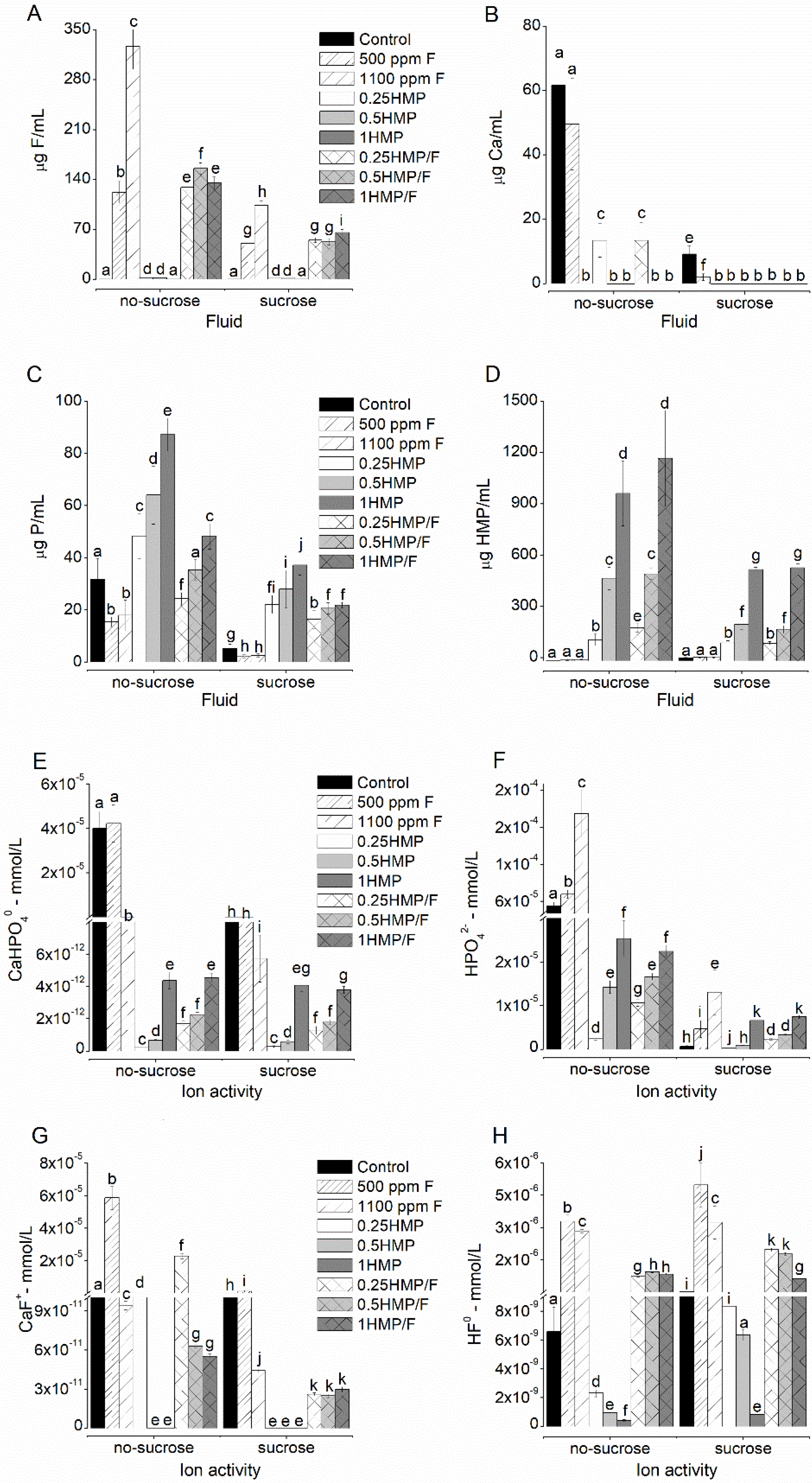
2.2. F, Ca, and P Levels in the Biofilm Fluid
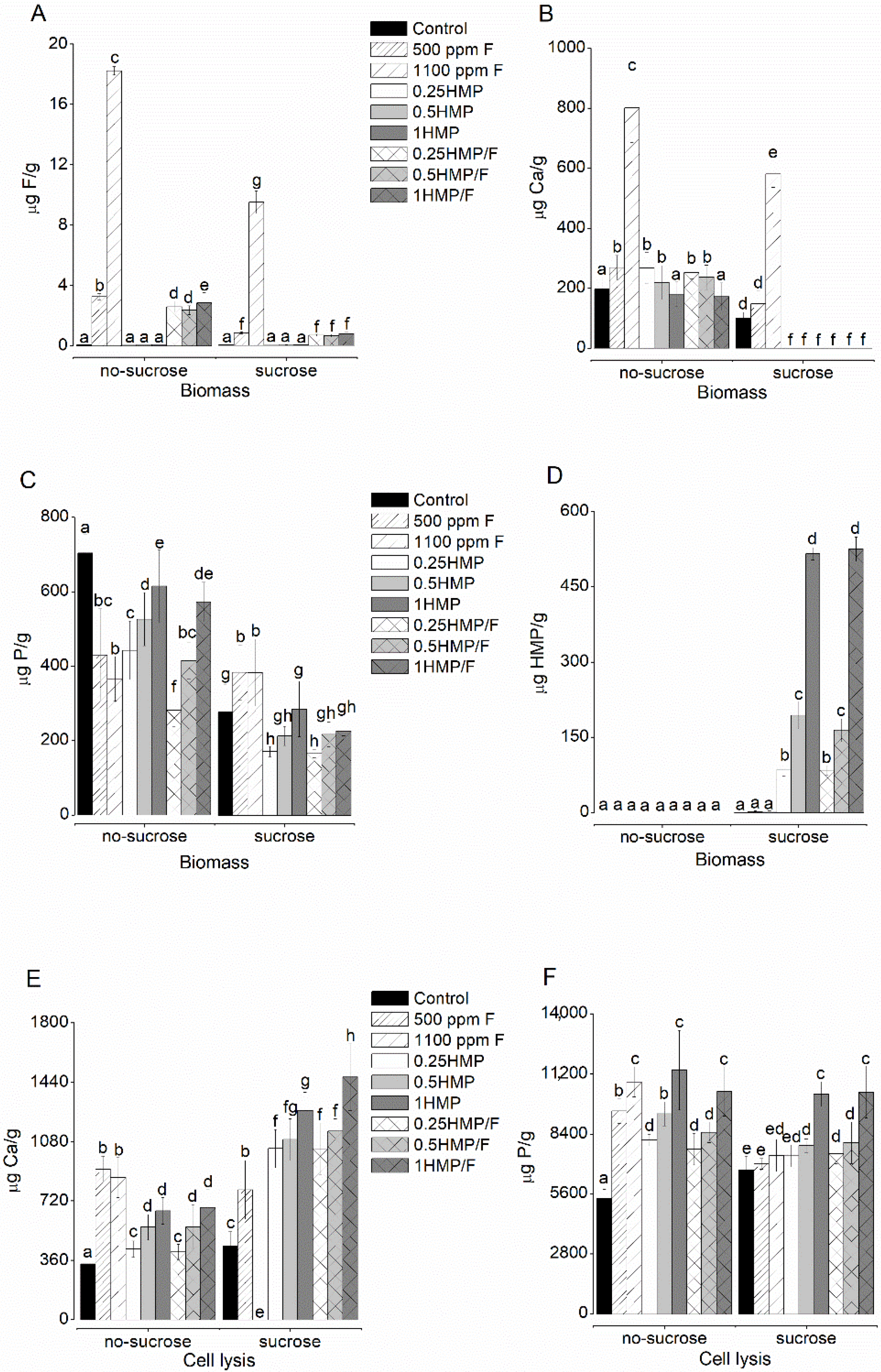
2.3. F, Ca, and P Levels in the Biofilm Biomass
2.4. Biofilm pH
2.5. Determination of Ionic Activities and Degree of Saturation from the Biofilm Fluid
3. Discussion
4. Materials and Methods
4.1. Microorganisms and Growth Conditions
4.2. Evaluation of the Number of Cells Prior to the Treatment of Biofilms
4.3. Treatment of Biofilms and pH Measurement
4.4. Analysis of F, Ca, and P Levels in the Biofilm Fluid
4.5. Analysis of F, Ca, and P Levels in the Biofilm Biomass
4.6. Determination of HMP and Ca Levels after Cell Lysis
4.7. Determination of Ionic Activities and Degree of Saturation from the Biofilm Fluid
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Machiulskiene, V.; Campus, G.; Carvalho, J.C.; Dige, I.; Ekstrand, K.R.; Jablonski-Momeni, A.; Maltz, M.; Manton, D.J.; Martignon, S.; Martinez-Mier, E.A.; et al. Terminology of Dental Caries and Dental Caries Management: Consensus Report of a Workshop Organized by ORCA and Cariology Research Group of IADR. Caries Res. 2020, 54, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Pitts, N.B.; Zero, D.T.; Marsh, P.D.; Ekstrand, K.; Weintraub, J.A.; Ramos-Gomez, F.; Tagami, J.; Twetman, S.; Tsakos, G.; Ismail, A. Dental Caries. Nat. Rev. Dis. Primers 2017, 3, 17030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamont, R.J.; Burne, R.A.; Lantz, M.S.; Leblanc, D.J. Oral Microbiology and Immunology; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- Bowen, W.H.; Burne, R.A.; Wu, H.; Koo, H. Oral biofilms: Pathogens, matrix, and polymicrobial interactions in microenvironments. Trends Microbiol. 2018, 26, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes. virulence of plaque biofilms in vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Grier, A.; Faustoferri, R.C.; Alzoubi, S.; Gill, A.L.; Feng, C.; Liu, Y.; Quivey, R.G.; Kopycka-Kedzierawski, D.T.; Koo, H.; et al. Association between oral Candida and bacteriome in children with severe ECC. J. Dent. Res. 2018, 97, 1468–1476. [Google Scholar] [CrossRef]
- Montelongo-Jauregui, D.; Lopez-Ribot, J.L. Candida interactions with the oral bacterial microbiota. J. Fungi 2018, 4, 122. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, A.A.; Souza, S.E.; Ricomini-Filho, A.P.; Del Bel Cury, A.A.; Cavalcanti, Y.W.; Cury, J.A. Candida albicans increases dentine demineralization provoked by Streptococcus mutans biofilm. Caries Res. 2019, 53, 322–331. [Google Scholar] [CrossRef]
- Vaara, M. Agents that increase the permeability of the outer membrane. Microbiol. Rev. 1992, 56, 395–411. [Google Scholar] [CrossRef]
- Van der Mei, H.C.; White, D.; Cox, E.; Geertsema-Doornbusch, G.; Busscher, H. Bacterial detachment from salivary conditioning films by dentifrice supernates. J. Clin. Dent. 2002, 13, 44–49. [Google Scholar]
- Garcia, L.S.G.; Delbem, A.C.B.; Pessan, J.P.; Dos Passos Silva, M.; Neto, F.N.S.; Gorup, L.F.; de Camargo, E.R.; Danelon, M. Anticaries effect of toothpaste with nano-sized sodium hexametaphosphate. Clin. Oral Investig. 2019, 23, 3535–3542. [Google Scholar] [CrossRef]
- Hosida, T.Y.; Pessan, J.P.; Cavazana, T.P.; Sampaio, C.; Monteiro, D.R.; Delbem, A.C.B. Effect of sodium hexametaphosphate and fluoride on dual-species biofilms of Candida albicans and Streptococcus mutans. Biofouling 2021, 37, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.M.; Hartman, P.A.; Stahr, H.M.; Olson, D.G.; Williams, F.D. Antibacterial mechanism of long-chain polyphosphates in Staphylococcus aureus. J. Food Prot. 1994, 57, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Changgen, L.; Yongxin, L. Selective flotation of scheelite from calcium minerals with sodium oleate as a collector and phosphates as modifiers. II. The mechanism of the interaction between phosphate modifiers and minerals. Int. J. Miner. Process. 1983, 10, 219–235. [Google Scholar] [CrossRef]
- ten Cate, J.M.; Dundon, K.A.; Vernon, P.G.; Damato, F.A.; Huntington, E.; Exterkate, R.A.; Wefel, J.S.; Jordan, T.; Stephen, K.W.; Roberts, A.J. Preparation and measurement of artificial enamel lesion, a four-laboratory ring test. Caries Res. 1996, 30, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Shaw, L.; Murray, J.J.; Burchell, K.; Best, J.S. Calcium and phosphorus content of plaque and saliva in relation to dental caries. Caries Res. 1983, 17, 543–548. [Google Scholar] [CrossRef]
- Tanaka, M.; Margolis, H.C. Release mineral ions in dental plaque following acid production. Arch. Oral Biol. 1999, 44, 253–258. [Google Scholar] [CrossRef]
- Pandit, S.; Jung, J.E.; Choi, H.M.; Jeon, J.G. Effect of brief periodic fluoride treatments on the virulence and composition of a cariogenic biofilm. Biofouling 2018, 34, 53–61. [Google Scholar] [CrossRef]
- Pandit, S.; Ca, J.N.; Jung, J.E.; Jeon, J.G. Effect of 1-minute fluoride treatment on potential virulence and viability of a cariogenic biofilm. Caries Res. 2015, 49, 449–457. [Google Scholar] [CrossRef]
- Lanigan, R.S. Final report on the safety assessment of sodium metaphosphate, sodium trimetaphosphate, and sodium hexametaphosphate. Int. J. Toxicol. 2001, 20, 75–89. [Google Scholar] [CrossRef]
- Buzalaf, M.A.R.; Pessan, J.P.; Honório, H.M.; ten Cate, J.M. Mechanisms of action of fluoride for caries control. Monogr. Oral Sci. 2011, 22, 97–114. [Google Scholar] [CrossRef]
- Cury, J.A.; Rebello, M.A.; Del Bel Cury, A.A. In situ relationship between sucrose exposure and the composition of dental plaque. Caries Res. 1997, 31, 356–360. [Google Scholar] [CrossRef]
- Qeli, E.; Toti, Ç.; Odorici, A.; Blasi, E.; Tragaj, E.; Tepedino, M.; Masedu, F.; Kaçani, G.; Hysi, D.; Meto, A.; et al. Effectiveness of Two Different Fluoride-Based Agents in the Treatment of Dentin Hypersensitivity: A Prospective Clinical Trial. Materials 2022, 15, 1266. [Google Scholar] [CrossRef] [PubMed]
- Klompmaker, S.H.; Kohl, K.; Fasel, N.; Mayer, A. Magnesium uptake by connecting fluid-phase endocytosis to an intracellular inorganic cation filter. Nat. Commun. 2017, 8, 1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochrane, N.J.; Saranathan, S.; Cai, F.; Cross, K.J.; Reynolds, E.C. Enamel subsurface lesion remineralization with casein phosphopeptide stabilized solutions of calcium, phosphate and fluoride. Caries Res. 2008, 42, 88–97. [Google Scholar] [CrossRef]
- Monteiro, D.R.; Silva, S.; Negri, M.; Gorup, L.F.; de Camargo, E.R.; Oliveira, R.; Barbosa, D.B.; Henriques, M. Silver colloidal nanoparticles: Effect on matrix composition and structure of Candida albicans and Candida glabrata biofilms. J. Appl. Microbiol. 2013, 114, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.A.; Monteiro, D.R.; Arias, L.S.; Fernandes, G.L.; Delbem, A.C.B.; Barbosa, D.B. Biofilm formation by Candida albicans and Streptococcus mutans in the presence of farnesol: A quantitative evaluation. Biofouling 2016, 32, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamfon, H.; Porter, S.R.; McCullough, M.; Pratten, J. Formation of Candida albicans biofilms on non-shedding oral surfaces. Eur. J. Oral Sci. 2003, 111, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Cavazana, T.P.; Pessan, J.P.; Hosida, T.Y.; Monteiro, D.R.; Delbem, A.C.B. pH changes of mixed biofilms of Streptococcus mutans and Candida albicans after exposure to sucrose solutions in vitro. Arch. Oral Biol. 2018, 90, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.; Schobel, B.; Scott-Anne, K.; Watson, G.; Bowen, W.H.; Cury, J.A.; Rosalen, P.L.; Park, Y.K. Apigenin and Tt-Farnesol with fluoride effects on S. mutans biofilms and dental caries. J. Dent. Res. 2005, 84, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Cavazana, T.P.; Pessan, J.P.; Hosida, T.Y.; Sampaio, C.; Amarante, V.O.Z.; Monteiro, D.R.; Delbem, A.C.B. Effects of Sodium Trimetaphosphate, Associated or Not with Fluoride, on the Composition and pH of Mixed Biofilms, before and after Exposure to Sucrose. Caries Res. 2020, 54, 358–368. [Google Scholar] [CrossRef]
- Vogel, G.L.; Chow, L.C.; Brown, W.E. A microanalytical procedure for the determination of calcium, phosphate and fluoride in enamel biopsy samples. Caries Res. 1983, 17, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Fiske, C.H.; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar] [CrossRef]
- Anderson, W.; Dingwall, D.; Stephen, K.W. Dissolution of two commercial preparations of calcium glycerophosphate in human saliva. Arch. Oral Biol. 1977, 22, 159–162. [Google Scholar] [CrossRef]
- Cury, J.A.; Rebelo, M.A.; Del Bel Cury, A.A.; Derbyshire, M.T.; Tabchoury, C.P. Biochemical composition and cariogenicity of dental plaque formed in the presence of sucrose or glucose and fructose. Caries Res. 2000, 34, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Nobre Dos Santos, M.; Melo Dos Santos, L.; Francisco, S.B.; Cury, J.A. Relationship among dental plaque composition, daily sugar exposure and caries in the primary dentition. Caries Res. 2002, 36, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Gaitán, A.C.; Fernández-Pereira, J.F.; Valentin, E.; Tamo-Mas, T.; Eraso, E.; Permán, J.; de Groot, P.W.J. Molecular identification of Candida auris by PCR amplification of species-specific GPI proyein-encoding genes. Int. J. Med. Microbiol. 2018, 308, 812–818. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Groups | pH | * Degree of Saturation (log) | ||||
|---|---|---|---|---|---|---|
| HA | CaF2 | |||||
| No Sucrose | Sucrose | No Sucrose | Sucrose | No Sucrose | Sucrose | |
| Control | 6.10 a,A (0.07) | 4.77 a,B (0.02) | −3.44 a,A (0.10) | −13.18 a,B (1.04) | 1.30 a,A (0.24) | −5.12 a,B (0.19) |
| 500 ppm F | 6.62 b,A (0.22) | 6.20 b,B (0.32) | 2.98 b,A (0.93) | −9.65 b,B (1.62) | 2.85 b,A (0.13) | 0.77 b,B (0.18) |
| 1100 ppm F | 7.08 c,A (0.16) | 6.49 c,B (0.31) | −29.40 c,A (1.52) | −34.62 c,B (1.02) | −3.23 c,A (0.38) | −4.24 c,B (0.06) |
| 0.25HMP | 6.46 bd,A (0.26) | 5.52 d,B (0.25) | 2.14 b,A (0.74) | −39.46 d,B (0.86) | −4.61 d,A (0.21) | −11.36 d,B (0.03) |
| 0.5HMP | 6.60 bf,A (0.05) | 5.73 e,B (0.14) | −29.80 c,A (0.52) | −36.26 e,B (0.43) | −11.22 e,A (0.06) | −11.02 e,B (0.26) |
| 1HMP | 6.87 cg,A (0.23) | 6.51 c,B (0.10) | −28.77 c,A (1.12) | −31.25 f,B (0.70) | −11.31 e,A (0.20) | −11.37 d,A (0.05) |
| 0.25HMP/F | 7.22 e,A (0.08) | 6.52 c,B (0.12) | 5.23 d,A (0.92) | −31.84 f,B (0.87) | 2.32 b,A (0.25) | −4.79 f,B (0.05) |
| 0.5HMP/F | 7.27 e,A (0.06) | 6.58 c,B (0.18) | −27.46 e,A (0.54) | −31.31 f,B (0.66) | −4.04 f,A (0.04) | −4.83 f,B (0.08) |
| 1HMP/F | 7.28 e,A (0.10) | 6.99 f,B (0.13) | −27.90 e,A (0.44) | −29.29 g,B (0.43) | −4.20 f,A (0.11) | −4.68 f,B (0.06) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosida, T.Y.; Pessan, J.P.; Cavazana, T.P.; Sampaio, C.; de Morais, L.A.; Monteiro, D.R.; Delbem, A.C.B. Effects of Sodium Hexametaphosphate and Fluoride on the pH and Inorganic Components of Streptococcus mutans and Candida albicans Biofilm after Sucrose Exposure. Antibiotics 2022, 11, 1044. https://doi.org/10.3390/antibiotics11081044
Hosida TY, Pessan JP, Cavazana TP, Sampaio C, de Morais LA, Monteiro DR, Delbem ACB. Effects of Sodium Hexametaphosphate and Fluoride on the pH and Inorganic Components of Streptococcus mutans and Candida albicans Biofilm after Sucrose Exposure. Antibiotics. 2022; 11(8):1044. https://doi.org/10.3390/antibiotics11081044
Chicago/Turabian StyleHosida, Thayse Yumi, Juliano Pelim Pessan, Thamires Priscila Cavazana, Caio Sampaio, Leonardo Antônio de Morais, Douglas Roberto Monteiro, and Alberto Carlos Botazzo Delbem. 2022. "Effects of Sodium Hexametaphosphate and Fluoride on the pH and Inorganic Components of Streptococcus mutans and Candida albicans Biofilm after Sucrose Exposure" Antibiotics 11, no. 8: 1044. https://doi.org/10.3390/antibiotics11081044