
Epidemiology of Plasmids in Escherichia coli and Klebsiella pneumoniae with Acquired Extended Spectrum Beta-Lactamase Genes Isolated from Chronic Wounds in Ghana

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
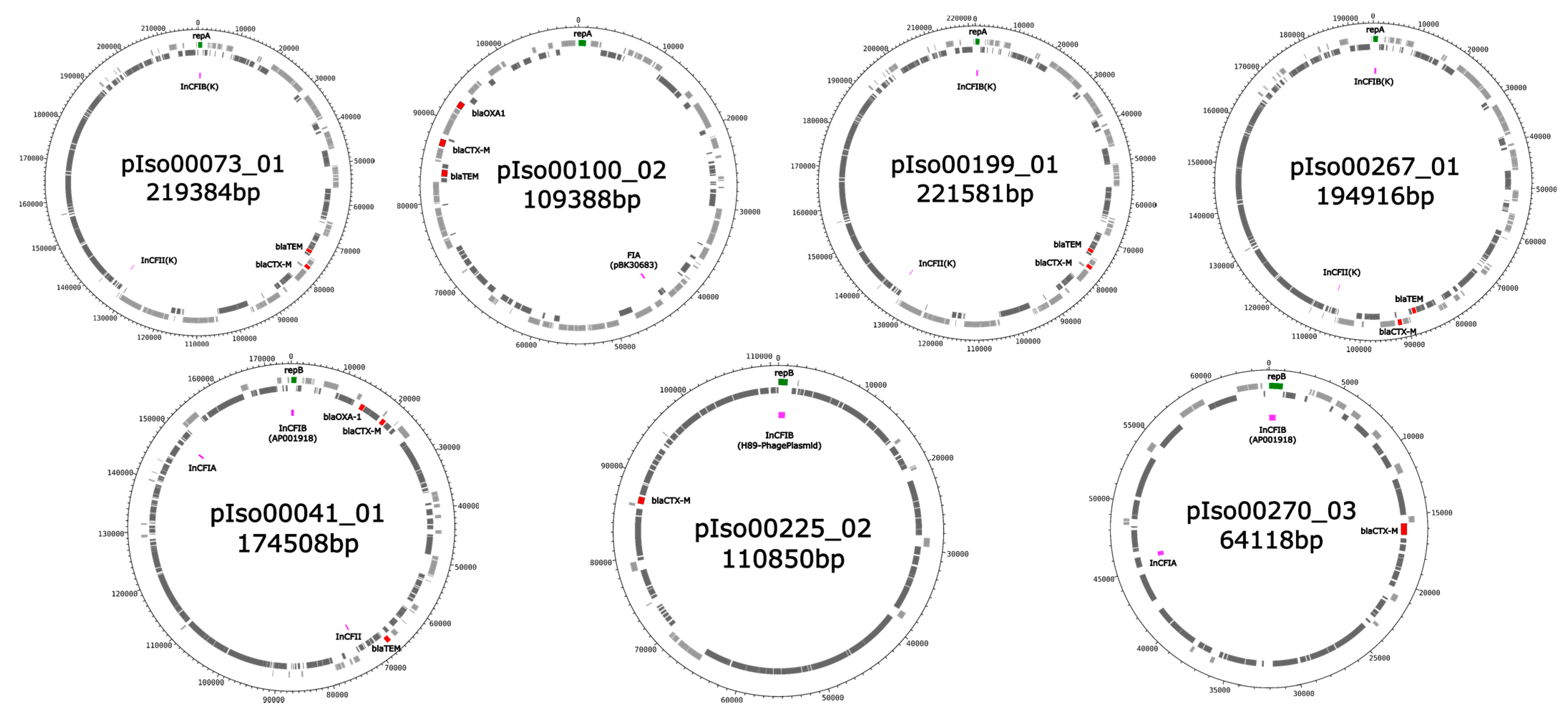
2. Results
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Bacterial Culture, Antibiotic Susceptibility Testing and Whole Genome Sequencing
4.2. Assessment of the Whole Genome Sequencing Data for Plasmids
4.3. Ethical Clearance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Strain/Sequence Type (ST)/GenBank Accession Number of Genomic DNA | Plasmid Number; GenBank Accession Number | Best Hit with Respect to the Query Coverage | Query Coverage (%) | Nucleotide Identity (%) | Geographic Site of NCBI Sequence Submission (Bacterial Species and Source) [Reference] |
|---|---|---|---|---|---|
| K. pneumoniae Iso00073/ST39/CP095150 | pIso00073_01; CP095151 | LR991402.1 | 97% | 99.99% | France (K. pneumoniae, source: unknown) [none] |
| pIso00073_02; CP095152 | CP063009.1 | 94% | 99.99% | Russian Federation (K. pneumoniae, source: human) [75] | |
| pIso00073_03; CP095153 | CP054171.1 | 95% | 99.86% | India (K. pneumoniae, source: human) [none] | |
| K. pneumoniae Iso00100/ST152/CP095145 | pIso00100_01; CP095146 | CP065826.1 | 79% | 99.95% | United States of America, (K. pneumoniae, source: human) [none] |
| pIso00100_02; CP095147 | CP023950.1 | 80% | 99.48% | Canada (K. pneumoniae, source: human) [none] | |
| K. pneumoniae Iso00199/ST17/CP095140 | pIso00199_01; CP095141 | LR991402.1 | 96% | 99.99% | France (K. pneumoniae, source: unknown) [none] |
| K. pneumoniae Iso00267/ST36/CP095132 | pIso00267_01; CP095133 | LR991402.1 | 94% | 99.99% | France (K. pneumoniae, source: unknown) [none] |
| pIso00267_02; CP095134 | CP016810.1 | 72% | 99.97% | United States of America, (K. pneumoniae, source: human) [none] | |
| E. coli Iso00041/ST 2/CP095155 | pIso00041_01; CP095156 | LR890289.1 | 99% | 99.98% | Australia (E. coli, source: unknown) [none] |
| E. coli Iso00225/ST506/CP095137 | pIso00225_01; CP095138 | CP088462.1 | 100% | 100% | South Korea (E. coli, source: human) [none] |
| pIso00225_02; CP095139 | MW590712.1 | 100% | 100% | United Kingdom (E. coli, source: human) [60] | |
| E. coli Iso00270/ST2/CP095125 | pIso00270_03; CP095128 | CP023816.1 | 96% | 99.11% | Germany (E. coli, source: wild bird) [61] |
| Species and Strain Number (73) | Sequence Type | Acquired Antimicrobial Resistance Genes | Recorded Phenotypic Resistance Against Apart From Penicillins and Cephalosporines * | Short-Read Archive (SRA) Accession Number |
|---|---|---|---|---|
| Klebsiella pneumoniae (73) | ST39 | blaTEM-1B, blaCTX-M-15,sul1, fosA, dfrA27, erm(B), mph(A), tet(D), oqxB, oqxA, aac(6′)-Ib-cr, qnrB2, catA2-like, aadA16, aac(3)-IIa, aph(3″)-Ib, aph(6)-Id | gentamicin, ciprofloxacin, moxifloxacin, trimethoprim/sulfamethoxazole | SRR13617236 |
| Klebsiella pneumoniae (100) | ST152 | blaCTX-M-15, blaOXA-1, blaTEM-1B,sul2, sul1, dfrA1, dfrA27, mph(A), aac(6′)-Ib-cr, oqxB, qnrB6, oqxA, catB3, catA1, ARR-3, aac(3)-IIa, aph(6)-Id, aph(3″)-Ib, aadA1, aadA16, aph(3′)-Ia | gentamicin, ciprofloxacin, moxifloxacin, trimethoprim/sulfamethoxazole | SRR13617311 |
| Klebsiella pneumoniae (199) | ST17 | blaCTX-M-15, blaTEM-1B,sul2, sul1, fosA-like, dfrA16, oqxA, oqxB, aadA2b, aac(3)-IIa | gentamicin, trimethoprim/sulfamethoxazole | SRR13617280 |
| Klebsiella pneumoniae (267) | ST36 | blaCTX-M-15, blaTEM-1B,sul2, sul1, fosA, dfrA27, tet(D), aac(6′)-Ib-cr, oqxA, oqxB, catA2-like, ARR-3, aph(6)-Id, aph(3″)-Ib, aadA16, aac(3)-IIa | gentamicin, trimethoprim/sulfamethoxazole | SRR13617257 |
| Escherichia coli (41) | ST2 | blaOXA-1, blaTEM-1B, blaCTX-M-15,sul1, dfrA17, mph(A), tet(B), aac(6′)-Ib-cr, catB3, catA1, aac(3)-IId, aadA5, mdf(A) | gentamicin, ciprofloxacin, moxifloxacin, trimethoprim/sulfamethoxazole | SRR13617294 |
| Escherichia coli (225) | ST506 | blaTEM-1D, blaCTX-M-15,sul1, sul2, dfrA17, mph(A), tet(A), catA1, aadA5, aph(6)-Id, aph(3″)-Ib, mdf(A)-like | moxifloxacin, trimethoprim/sulfamethoxazole | SRR13617270 |
| Escherichia coli (270) | ST2 | blaCTX-M-15, tet(B), catA1, mdf(A) | ciprofloxacin, moxifloxacin | SRR13617256 |
References
- Mama, M.; Abdissa, A.; Sewunet, T. Antimicrobial susceptibility pattern of bacterial isolates from wound infection and their sensitivity to alternative topical agents at Jimma University Specialized Hospital, South-West Ethiopia. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moremi, N.; Mushi, M.F.; Fidelis, M.; Chalya, P.; Mirambo, M.; Mshana, S.E. Predominance of multi-resistant gram-negative bacteria colonizing chronic lower limb ulcers (CLLUs) at Bugando Medical Center. BMC Res. Notes 2014, 7, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassam, N.A.; Damian, D.J.; Kajeguka, D.; Nyombi, B.; Kibiki, G.S. Spectrum and antibiogram of bacteria isolated from patients presenting with infected wounds in a Tertiary Hospital, northern Tanzania. BMC Res. Notes 2017, 10, 757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, H.; Janssen, I.; Cooper, P.; Kainyah, C.; Pellio, T.; Quintel, M.; Monnheimer, M.; Groß, U.; Schulze, M.H. Antimicrobial-Resistant Bacteria in Infected Wounds, Ghana, 2014. Emerg. Infect. Dis. 2018, 24, 916–919. [Google Scholar] [CrossRef]
- Kazimoto, T.; Abdulla, S.; Bategereza, L.; Juma, O.; Mhimbira, F.; Weisser, M.; Utzinger, J.; von Müller, L.; Becker, S.L. Causative agents and antimicrobial resistance patterns of human skin and soft tissue infections in Bagamoyo, Tanzania. Acta Trop. 2018, 186, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Lai, P.S.; Bebell, L.M.; Meney, C.; Valeri, L.; White, M.C. Epidemiology of antibiotic-resistant wound infections from six countries in Africa. BMJ Glob. Health 2018, 2 (Suppl. S4), e000475. [Google Scholar] [CrossRef] [Green Version]
- San Millan, A. Evolution of Plasmid-Mediated Antibiotic Resistance in the Clinical Context. Trends Microbiol. 2018, 26, 978–985. [Google Scholar] [CrossRef] [Green Version]
- San Millan, A.; MacLean, R.C. Fitness Costs of Plasmids: A Limit to Plasmid Transmission. Microbiol. Spectr. 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Loftie-Eaton, W.; Bashford, K.; Quinn, H.; Dong, K.; Millstein, J.; Hunter, S.; Thomason, M.K.; Merrikh, H.; Ponciano, J.M.; Top, E.M. Compensatory mutations improve general permissiveness to antibiotic resistance plasmids. Nat. Ecol. Evol. 2017, 1, 1354–1363. [Google Scholar] [CrossRef]
- Dunn, S.J.; Connor, C.; McNally, A. The evolution and transmission of multi-drug resistant Escherichia coli and Klebsiella pneumoniae: The complexity of clones and plasmids. Curr. Opin. Microbiol. 2019, 51, 51–56. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids in Gram negatives: Molecular typing of resistance plasmids. Int. J. Med. Microbiol. 2011, 301, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Choi, M.J.; Ko, K.S. Replicon sequence typing of IncF plasmids and the genetic environments of blaCTX-M-15 indicate multiple acquisitions of blaCTX-M-15 in Escherichia coli and Klebsiella pneumoniae isolates from South Korea. J. Antimicrob. Chemother. 2012, 67, 1853–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machuca, J.; López-Cerero, L.; Fernández-Cuenca, F.; Mora-Navas, L.; Mediavilla-Gradolph, C.; López-Rodríguez, I.; Pascual, Á. OXA-48-Like-Producing Klebsiella pneumoniae in Southern Spain in 2014-2015. Antimicrob. Agents Chemother. 2018, 63, e01396-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Tang, Y.; Fu, P.; Tian, D.; Yu, L.; Huang, Y.; Li, G.; Li, M.; Wang, Y.; Yang, Z.; et al. The type I-E CRISPR-Cas system influences the acquisition of bla(KPC)-IncF plasmid in Klebsiella pneumoniae. Emerg. Microbes Infect. 2020, 9, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Fu, P.; Zhou, Y.; Xie, Y.; Jin, J.; Wang, B.; Yu, L.; Huang, Y.; Li, G.; Li, M.; et al. Absence of the type I-E CRISPR-Cas system in Klebsiella pneumoniae clonal complex 258 is associated with dissemination of IncF epidemic resistance plasmids in this clonal complex. J. Antimicrob. Chemother. 2020, 75, 890–895. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, W.; Zhao, Z.; Ye, C.; Zhou, S.; Wu, S.; Han, L.; Han, Z.; Ye, H. Molecular characterization of carbapenem-resistant Klebsiella pneumoniae isolates with focus on antimicrobial resistance. BMC Genom. 2019, 20, 822. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Liu, L.; McNally, A.; Zong, Z. Coexistence of three bla(KPC-2) genes on an IncF/IncR plasmid in ST11 Klebsiella pneumoniae. J. Glob. Antimicrob. Resist. 2019, 17, 90–93. [Google Scholar] [CrossRef]
- Peirano, G.; Bradford, P.A.; Kazmierczak, K.M.; Chen, L.; Kreiswirth, B.N.; Pitout, J.D. Importance of Clonal Complex 258 and IncF(K2-like) Plasmids among a Global Collection of Klebsiella pneumoniae with bla(KPC). Antimicrob. Agents Chemother. 2017, 61, e02610-16. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Vázquez, M.; Sola Campoy, P.J.; Ortega, A.; Bautista, V.; Monzón, S.; Ruiz-Carrascoso, G.; Mingorance, J.; González-Barberá, E.M.; Gimeno, C.; Aracil, B.; et al. Emergence of NDM-producing Klebsiella pneumoniae and Escherichia coli in Spain: Phylogeny, resistome, virulence and plasmids encoding blaNDM-like genes as determined by WGS. J. Antimicrob. Chemother. 2019, 74, 3489–3496. [Google Scholar] [CrossRef]
- Villa, L.; García-Fernández, A.; Fortini, D.; Carattoli, A. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 2010, 65, 2518–2529. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaibani, P.; Scaltriti, E.; Benni, C.; Pongolini, S.; Ambretti, S.; Landini, M.P.; Viale, P.; Giannella, M.; Re, M.C. Characterization of an IncL/M plasmid carrying blaOXA-48 in a Klebsiella pneumoniae strain from Italy. New Microbiol. 2017, 40, 284–285. [Google Scholar] [PubMed]
- Carattoli, A.; Seiffert, S.N.; Schwendener, S.; Perreten, V.; Endimiani, A. Differentiation of IncL and IncM Plasmids Associated with the Spread of Clinically Relevant Antimicrobial Resistance. PLoS ONE 2015, 10, e0123063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamczuk, M.; Zaleski, P.; Dziewit, L.; Wolinowska, R.; Nieckarz, M.; Wawrzyniak, P.; Kieryl, P.; Plucienniczak, A.; Bartosik, D. Diversity and Global Distribution of IncL/M Plasmids Enabling Horizontal Dissemination of β-Lactam Resistance Genes among the Enterobacteriaceae. Biomed. Res. Int. 2015, 2015, 414681. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, É.M.; Beltrão, E.M.B.; Scavuzzi, A.M.L.; Barros, J.F.; Lopes, A.C.S. High plasmid variability, and the presence of IncFIB, IncQ, IncA/C, IncHI1B, and IncL/M in clinical isolates of Klebsiella pneumoniae with blaKPC and blaNDM from patients at a public hospital in Brazil. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200397. [Google Scholar] [CrossRef]
- Rada, A.M.; De La Cadena, E.; Agudelo, C.; Capataz, C.; Orozco, N.; Pallares, C.; Dinh, A.Q.; Panesso, D.; Ríos, R.; Diaz, L.; et al. Dynamics of bla(KPC-2) Dissemination from Non-CG258 Klebsiella pneumoniae to Other Enterobacterales via IncN Plasmids in an Area of High Endemicity. Antimicrob. Agents Chemother. 2020, 64, e01743-20. [Google Scholar] [CrossRef]
- Aires-de-Sousa, M.; Ortiz de la Rosa, J.M.; Gonçalves, M.L.; Pereira, A.L.; Nordmann, P.; Poirel, L. Epidemiology of Carbapenemase-Producing Klebsiella pneumoniae in a Hospital, Portugal. Emerg. Infect. Dis. 2019, 25, 1632–1638. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Higgins, C.H.; Rehman, I.; Galvao, K.N.; Brito, I.L.; Bicalho, M.L.; Song, J.; Wang, H.; Bicalho, R.C. Genomic Diversity, Virulence, and Antimicrobial Resistance of Klebsiella pneumoniae Strains from Cows and Humans. Appl. Environ. Microbiol. 2019, 85, e02654-18. [Google Scholar] [CrossRef] [Green Version]
- Hao, M.; He, Y.; Zhang, H.; Liao, X.P.; Liu, Y.H.; Sun, J.; Du, H.; Kreiswirth, B.N.; Chen, L. CRISPR-Cas9-Mediated Carbapenemase Gene and Plasmid Curing in Carbapenem-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2020, 64, e00843-20. [Google Scholar] [CrossRef]
- García-Fernández, A.; Villa, L.; Moodley, A.; Hasman, H.; Miriagou, V.; Guardabassi, L.; Carattoli, A. Multilocus sequence typing of IncN plasmids. J. Antimicrob. Chemother. 2011, 66, 1987–1991. [Google Scholar] [CrossRef]
- Jing, Y.; Jiang, X.; Yin, Z.; Hu, L.; Zhang, Y.; Yang, W.; Yang, H.; Gao, B.; Zhao, Y.; Zhou, D.; et al. Genomic diversification of IncR plasmids from China. J. Glob. Antimicrob. Resist. 2019, 19, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Liu, L.; Zhang, R.; Chen, K.; Xie, M.; Chan, E.W.C.; Chen, S. An IncR Plasmid Harbored by a Hypervirulent Carbapenem-Resistant Klebsiella pneumoniae Strain Possesses Five Tandem Repeats of the bla (KPC-2)::NTE(KPC)-Id Fragment. Antimicrob. Agents Chemother. 2019, 63, e01775-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocsis, E.; Gužvinec, M.; Butić, I.; Krešić, S.; Crnek, S.Š.; Tambić, A.; Cornaglia, G.; Mazzariol, A. blaNDM-1 Carriage on IncR Plasmid in Enterobacteriaceae Strains. Microb. Drug Resist. 2016, 22, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Gamal, D.; Fernández-Martínez, M.; Salem, D.; El-Defrawy, I.; Montes, L.Á.; Ocampo-Sosa, A.A.; Martínez-Martínez, L. Carbapenem-resistant Klebsiella pneumoniae isolates from Egypt containing blaNDM-1 on IncR plasmids and its association with rmtF. Int. J. Infect. Dis. 2016, 43, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, C.; Zhu, X.; Lu, J.; Shen, K.; Chen, Q.; Zhou, W.; Liu, H.; Lu, W.; Zhou, D.; Sun, Z.; et al. Characterization of an IncR Plasmid with Two Copies of ISCR-Linked qnrB6 from ST968 Klebsiella pneumoniae. Int. J. Genom. 2020, 2020, 3484328. [Google Scholar] [CrossRef]
- Guo, Q.; Spychala, C.N.; McElheny, C.L.; Doi, Y. Comparative analysis of an IncR plasmid carrying armA, blaDHA-1 and qnrB4 from Klebsiella pneumoniae ST37 isolates. J. Antimicrob. Chemother. 2016, 71, 882–886. [Google Scholar] [CrossRef] [Green Version]
- Compain, F.; Frangeul, L.; Drieux, L.; Verdet, C.; Brisse, S.; Arlet, G.; Decré, D. Complete nucleotide sequence of two multidrug-resistant IncR plasmids from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2014, 58, 4207–4210. [Google Scholar] [CrossRef] [Green Version]
- Qu, D.; Shen, Y.; Hu, L.; Jiang, X.; Yin, Z.; Gao, B.; Zhao, Y.; Yang, W.; Yang, H.; Han, J.; et al. Comparative analysis of KPC-2-encoding chimera plasmids with multi-replicon IncR:Inc(pA1763-KPC):IncN1 or IncFII(pHN7A8):Inc(pA1763-KPC):IncN1. Infect. Drug Resist. 2019, 12, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- De Maio, N.; Shaw, L.P.; Hubbard, A.; George, S.; Sanderson, N.D.; Swann, J.; Wick, R.; AbuOun, M.; Stubberfield, E.; Hoosdally, S.J.; et al. Comparison of long-read sequencing technologies in the hybrid assembly of complex bacterial genomes. Microb. Genom. 2019, 5, e000294. [Google Scholar] [CrossRef]
- Suzuki, Y.; Nishijima, S.; Furuta, Y.; Yoshimura, J.; Suda, W.; Oshima, K.; Hattori, M.; Morishita, S. Long-read metagenomic exploration of extrachromosomal mobile genetic elements in the human gut. Microbiome 2019, 7, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perine, P.L.; Totten, P.A.; Knapp, J.S.; Holmes, K.K.; Bentsi, C.; Klufio, C.A. Diversity of gonococcal plasmids, auxotypes, and serogroups in Ghana. Lancet 1983, 1, 1051–1052. [Google Scholar] [CrossRef]
- Labar, A.S.; Millman, J.S.; Ruebush, E.; Opintan, J.A.; Bishar, R.A.; Aboderin, A.O.; Newman, M.J.; Lamikanra, A.; Okeke, I.N. Regional dissemination of a trimethoprim-resistance gene cassette via a successful transposable element. PLoS ONE 2012, 7, e38142. [Google Scholar] [CrossRef] [PubMed]
- Oduro-Mensah, D.; Obeng-Nkrumah, N.; Bonney, E.Y.; Oduro-Mensah, E.; Twum-Danso, K.; Osei, Y.D.; Sackey, S.T. Genetic characterization of TEM-type ESBL-associated antibacterial resistance in Enterobacteriaceae in a tertiary hospital in Ghana. Ann. Clin. Microbiol Antimicrob. 2016, 15, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudirkiene, E.; Andoh, L.A.; Ahmed, S.; Herrero-Fresno, A.; Dalsgaard, A.; Obiri-Danso, K.; Olsen, J.E. The Use of a Combined Bioinformatics Approach to Locate Antibiotic Resistance Genes on Plasmids from Whole Genome Sequences of Salmonella enterica Serovars from Humans in Ghana. Front. Microbiol. 2018, 9, 1010. [Google Scholar] [CrossRef]
- Ayibieke, A.; Sato, W.; Mahazu, S.; Prah, I.; Addow-Thompson, J.; Ohashi, M.; Suzuki, T.; Iwanaga, S.; Ablordey, A.; Saito, R. Molecular characterisation of the NDM-1-encoding plasmid p2189-NDM in an Escherichia coli ST410 clinical isolate from Ghana. PLoS ONE 2018, 13, e0209623. [Google Scholar] [CrossRef] [Green Version]
- Agyepong, N.; Govinden, U.; Owusu-Ofori, A.; Amoako, D.G.; Allam, M.; Janice, J.; Pedersen, T.; Sundsfjord, A.; Essack, S. Genomic characterization of multidrug-resistant ESBL-producing Klebsiella pneumoniae isolated from a Ghanaian teaching hospital. Int. J. Infect. Dis. 2019, 85, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Moirongo, R.M.; Lorenz, E.; Ntinginya, N.E.; Dekker, D.; Fernandes, J.; Held, J.; Lamshöft, M.; Schaumburg, F.; Mangu, C.; Sudi, L.; et al. Regional Variation of Extended-Spectrum Beta-Lactamase (ESBL)-Producing Enterobacterales, Fluoroquinolone-Resistant Salmonella enterica and Methicillin-Resistant Staphylococcus aureus Among Febrile Patients in Sub-Saharan Africa. Front. Microbiol. 2020, 11, 567235. [Google Scholar] [CrossRef]
- Acheampong, G.; Owusu, M.; Owusu-Ofori, A.; Osei, I.; Sarpong, N.; Sylverken, A.; Kung, H.J.; Cho, S.T.; Kuo, C.H.; Park, S.E.; et al. Chromosomal and plasmid-mediated fluoroquinolone resistance in human Salmonella enterica infection in Ghana. BMC Infect. Dis. 2019, 19, 898. [Google Scholar] [CrossRef] [Green Version]
- Eibach, D.; Nagel, M.; Lorenzen, S.; Hogan, B.; Belmar Campos, C.; Aepfelbacher, M.; Sarpong, N.; May, J. Extended-spectrum β-lactamase-producing Enterobacteriaceae among geckos (Hemidactylus brookii) in a Ghanaian hospital. Clin. Microbiol. Infect. 2019, 25, 1048–1050. [Google Scholar] [CrossRef]
- Adzitey, F.; Assoah-Peprah, P.; Teye, G.A. Whole-genome sequencing of Escherichia coli isolated from contaminated meat samples collected from the Northern Region of Ghana reveals the presence of multiple antimicrobial resistance genes. J. Glob. Antimicrob. Resist. 2019, 18, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Imirzalioglu, C.; Oppong, K.; Akenten, C.W.; Hogan, B.; Krumkamp, R.; Poppert, S.; Levermann, V.; Schwengers, O.; Sarpong, N.; et al. Detection and Characterization of ESBL-Producing Escherichia coli from Humans and Poultry in Ghana. Front. Microbiol. 2019, 9, 3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayibieke, A.; Kobayashi, A.; Suzuki, M.; Sato, W.; Mahazu, S.; Prah, I.; Mizoguchi, M.; Moriya, K.; Hayashi, T.; Suzuki, T.; et al. Prevalence and Characterization of Carbapenem-Hydrolyzing Class D β-Lactamase-Producing Acinetobacter Isolates from Ghana. Front. Microbiol. 2020, 11, 587398. [Google Scholar] [CrossRef] [PubMed]
- Krumkamp, R.; Oppong, K.; Hogan, B.; Strauss, R.; Frickmann, H.; Wiafe-Akenten, C.; Boahen, K.G.; Rickerts, V.; McCormick Smith, I.; Groß, U.; et al. Spectrum of antibiotic resistant bacteria and fungi isolated from chronically infected wounds in a rural district hospital in Ghana. PLoS ONE 2020, 15, e0237263. [Google Scholar] [CrossRef] [PubMed]
- Codjoe, F.S.; Donkor, E.S.; Smith, T.J.; Miller, K. Phenotypic and Genotypic Characterization of Carbapenem-Resistant Gram-Negative Bacilli Pathogens from Hospitals in Ghana. Microb. Drug Resist. 2019, 25, 1449–1457. [Google Scholar] [CrossRef]
- Dekker, D.; Pankok, F.; Thye, T.; Taudien, S.; Oppong, K.; Wiafe Akenten, C.; Lamshöft, M.; Jaeger, A.; Kaase, M.; Scheithauer, S.; et al. Clonal Clusters, Molecular Resistance Mechanisms and Virulence Factors of Gram-Negative Bacteria Isolated from Chronic Wounds in Ghana. Antibiotics 2021, 10, 339. [Google Scholar] [CrossRef]
- Founou, L.L.; Founou, R.C.; Allam, M.; Ismail, A.; Djoko, C.F.; Essack, S.Y. Genome Sequencing of Extended-Spectrum β-Lactamase (ESBL)-Producing Klebsiella pneumoniae Isolated from Pigs and Abattoir Workers in Cameroon. Front. Microbiol. 2018, 9, 188. [Google Scholar] [CrossRef]
- Minja, C.A.; Shirima, G.; Mshana, S.E. Conjugative Plasmids Disseminating CTX-M-15 among Human, Animals and the Environment in Mwanza Tanzania: A Need to Intensify One Health Approach. Antibiotics 2021, 10, 836. [Google Scholar] [CrossRef]
- Pedersen, T.; Tellevik, M.G.; Kommedal, Ø.; Lindemann, P.C.; Moyo, S.J.; Janice, J.; Blomberg, B.; Samuelsen, Ø.; Langeland, N. Horizontal Plasmid Transfer among Klebsiella pneumoniae Isolates Is the Key Factor for Dissemination of Extended-Spectrum β-Lactamases among Children in Tanzania. mSphere 2020, 5, e00428-20. [Google Scholar] [CrossRef]
- Bevan, E.R.; Powell, M.J.; Toleman, M.A.; Thomas, C.M.; Piddock, L.J.V.; Hawkey, P.M. Molecular characterization of plasmids encoding blaCTX-M from faecal Escherichia coli in travellers returning to the UK from South Asia. J. Hosp. Infect. 2021, 114, 134–143. [Google Scholar] [CrossRef]
- Schaufler, K.; Semmler, T.; Wieler, L.H.; Trott, D.J.; Pitout, J.; Peirano, G.; Bonnedahl, J.; Dolejska, M.; Literak, I.; Fuchs, S.; et al. Genomic and Functional Analysis of Emerging Virulent and Multidrug-Resistant Escherichia coli Lineage Sequence Type 648. Antimicrob. Agents Chemother. 2019, 63, e00243-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnedahl, J.; Järhult, J.D. Antibiotic resistance in wild birds. Upsala J. Med. Sci. 2014, 119, 113–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, C.A.; Zarazaga, M.; Ben Sallem, R.; Jouini, A.; Ben Slama, K.; Torres, C. Antibiotic resistance in Escherichia coli in husbandry animals: The African perspective. Lett. Appl. Microbiol. 2017, 64, 318–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutai, W.C.; Waiyaki, P.G.; Kariuki, S.; Muigai, A.W.T. Plasmid profiling and incompatibility grouping of multidrug resistant Salmonella enterica serovar Typhi isolates in Nairobi, Kenya. BMC Res. Notes 2019, 12, 422. [Google Scholar] [CrossRef]
- Inouye, M.; Dashnow, H.; Raven, L.A.; Schultz, M.B.; Pope, B.J.; Tomita, T.; Zobel, J.; Holt, K.E. SRST2: Rapid genomic surveillance for public health and hospital microbiology labs. Genome Med. 2014, 6, 90. [Google Scholar] [CrossRef] [Green Version]
- Delahaye, C.; Nicolas, J. Sequencing DNA with nanopores: Troubles and biases. PLoS ONE 2021, 16, e0257521. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Hasman, H. PlasmidFinder and In Silico pMLST: Identification and Typing of Plasmid Replicons in Whole-Genome Sequencing (WGS). Methods Mol. Biol. 2020, 2075, 285–294. [Google Scholar]
- Johnson, T.J.; Nolan, L.K. Plasmid replicon typing. Methods Mol. Biol. 2009, 551, 27–35. [Google Scholar]
- Seemann, T. Abricate. Available online: https://github.com/tseemann/abricate (accessed on 11 May 2022).
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carver, T.; Thomson, N.; Bleasby, A.; Berriman, M.; Parkhill, J. DNAPlotter: Circular and linear interactive genome visualization. Bioinformatics 2009, 25, 119–120. [Google Scholar] [CrossRef] [PubMed]
- Shelenkov, A.; Mikhaylova, Y.; Yanushevich, Y.; Samoilov, A.; Petrova, L.; Fomina, V.; Gusarov, V.; Zamyatin, M.; Shagin, D.; Akimkin, V. Molecular Typing, Characterization of Antimicrobial Resistance, Virulence Profiling and Analysis of Whole-Genome Sequence of Clinical Klebsiella pneumoniae Isolates. Antibiotics 2020, 9, 261. [Google Scholar] [CrossRef]

| Species and Isolate Number/MLST Type of the Isolate/GenBank Accession Number of Genomic DNA | Plasmid Id and GenBank Accession Number | Plasmid Size | Typing Results Based on PlasmidFinder-2.0 and Mob-Typer | Predicted Mobility Based on Mob-Typer | Resistance Genes on the Plasmid |
|---|---|---|---|---|---|
| K. pneumoniae Iso00073/ST39/CP095150 | pIso00073_01; CP095151 | 219,384 | IncFIB, IncFII, rep_cluster_2183 | conjugative | aac(3)-IIa, blaTEM-1B *, blaCTX-M-15 |
| pIso00073_02; CP095152 | 92,025 | IncFIA, IncR | mobilizable | sul2, aph(3″)-Ib, aph(3″)-Ib, aph(6)-Id, blaTEM-1B *, catA2-like, tet(D), aac(6′)-Ib-cr, ARR-3, dfrA27, aadA16, qacE, sul1, qnrB2 | |
| pIso00073_03; CP095153 | 82,442 | IncFII, IncFIA, IncFIC | conjugative | erm(B), mph(A), blaTEM-1B * | |
| pIso00073_04; CP095154 | 4350 | ColRNAI_rep_cluster_1987 | non-mobilizable | - | |
| K. pneumoniae Iso00100/ST152/CP095145 | pIso00100_01; CP095146 | 224,675 | IncFIB, IncFII, rep_cluster_2183 | conjugative | dfrA1, aadA1, qacE, sul1, tet(D), blaSHV-187 *, catA1 |
| pIso00100_02; CP095147 | 109,388 | FIA, rep_cluster_1418 | mobilizable | mph(A), aph(3’)-Ia, sul1, qacE *, qnrB6, aadA16, dfrA27, ARR-3, aac(6’)-Ib-cr *, sul2, aph(3’’)-Ib, aph(6)-Id, blaTEM-1B, blaCTX-M-15, blaOXA-1, catB3, aac(3)-IIa | |
| pIso00100_03; CP095148 | 8282 | ColRNAI_rep_cluster_1857 | mobilizable | - | |
| pIso00100_04; CP095149 | 4642 | Col440I, ColRNAI_rep_cluster_1987 | non-mobilizable | - | |
| K. pneumoniae Iso00199/ST17/CP095140 | pIso00199_01; CP095141 | 221,581 | IncFIB, IncFII, rep_cluster_2183 | conjugative | aac(3)-IIa, blaTEM-1B *, blaCTX-M-15 |
| pIso00199_02; CP095142 | 52,096 | IncP1 | conjugative | - | |
| pIso00199_03; CP095143 | 4204 | rep_cluster_2358 | non-mobilizable | - | |
| pIso00199_04; CP095144 | 3643 | ColRNAI_rep_cluster_1987 | non-mobilizable | - | |
| K. pneumoniae Iso00267/ST36/CP095132 | pIso00267_01; CP095133 | 194,916 | IncFIB, IncFII, rep_cluster_2183 | conjugative | aac(3)-IIa, blaTEM-1B, blaCTX-M-15 |
| pIso00267_02; CP095134 | 70,936 | IncFIA, rep_cluster_1418 | mobilizable | tet(D), catA2-like, aph(6)-Id *, aph(3’’)-Ib *, sul2, aac(6’)-Ib-cr, ARR-3, dfrA27, aadA16, qacE, sul1 | |
| pIso00267_03; CP095135 | 9294 | ColRNAI_rep_cluster_1857 | mobilizable | - | |
| pIso00267_04; CP095136 | 5835 | Col(Ye4449) | mobilizable | - | |
| E. coli Iso00041/ST2/CP095155 | pIso00041_01; CP095156 | 174,508 | IncFIA, IncFIB, IncFIC | conjugative | catB3, blaOXA-1, aac(6′)-Ib-cr, sitABCD, blaCTX-M-15, tet(B), catA1, mph(A), sul1, qacE, aadA5, dfrA17, aac(3)-IId, blaTEM-1B |
| pIso00041_02; CP095157 | 5164 | IncFIB, rep_cluster_2131 | mobilizable | - | |
| pIso00041_03; CP095158 | 2348 | IncFII, Col(IRGK) | non-mobilizable | - | |
| pIso00041_04; CP095159 | 1538 | Col(MG828) | mobilizable | - | |
| E. coli Iso00225/ST506/CP095137 | pIso00225_01; CP095138 | 133,313 | Col156, IncFIA, IncFIB, IncFII, rep_cluster_2131 | conjugative | tet(A), aph(6)-Id, aph(3″)-Ib, sul2, mph(A), sul1, qacE, aadA5, dfrA17 |
| pIso00225_02; CP095139 | 110,850 | IncFIB, rep_cluster_488 | non-mobilizable | blaCTX-M-15 | |
| E. coli Iso00270/ST2/CP095125 | pIso00270_01; CP095126 | 99,933 | IncY | non-mobilizable | - |
| pIso00270_02; CP095127 | 94,223 | rep_cluster_1704 | non-mobilizable | - | |
| pIso00270_03; CP095128 | 64,118 | IncFIA, IncFIB, IncFII | mobilizable | catA1, blaCTX-M-15, tet(B) | |
| pIso00270_04; CP095129 | 5164 | rep_cluster_2131 | mobilizable | - | |
| pIso00270_05; CP095130 | 3007 | rep_cluster_2350 | mobilizable | - | |
| pIso00270_06; CP095131 | 2255 | Col(MG828) | non-mobilizable | - |
| Klebsiella pneumoniae | Escherichia coli | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Iso00073 | Iso00199 | Iso00267 | Iso00100 | Iso00041 | Iso00225 | Iso00270 | |||
| pIso00073_01 | pIso00199_01 | pIso00267_01 | pIso00100_02 | pIso00041_01 | pIso00225_02 | pIso00270_03 | |||
| Klebsiella pneumoniae | Iso00073 | pIso00073_01 | - | 100% | 83% | 9% | 11% | 1% | 7% |
| Iso00199 | pIso00199_01 | 99% | - | 83% | 9% | 11% | 1% | 7% | |
| Iso00267 | pIso00267_01 | 92% | 92% | - | 12% | 11% | 1% | 8% | |
| Iso00100 | pIso00100_02 | 21% | 21% | 25% | - | 35% | 2% | 14% | |
| Escherichia coli | Iso00041 | pIso00041_01 | 15% | 15% | 16% | 19% | - | 1% | 27% |
| Iso00225 | pIso00225_02 | 2% | 2% | 2% | 2% | 2% | - | 11% | |
| Iso00270 | pIso00270_03 | 23% | 23% | 24% | 18% | 65% | 3% | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pankok, F.; Taudien, S.; Dekker, D.; Thye, T.; Oppong, K.; Wiafe Akenten, C.; Lamshöft, M.; Jaeger, A.; Kaase, M.; Scheithauer, S.; et al. Epidemiology of Plasmids in Escherichia coli and Klebsiella pneumoniae with Acquired Extended Spectrum Beta-Lactamase Genes Isolated from Chronic Wounds in Ghana. Antibiotics 2022, 11, 689. https://doi.org/10.3390/antibiotics11050689
Pankok F, Taudien S, Dekker D, Thye T, Oppong K, Wiafe Akenten C, Lamshöft M, Jaeger A, Kaase M, Scheithauer S, et al. Epidemiology of Plasmids in Escherichia coli and Klebsiella pneumoniae with Acquired Extended Spectrum Beta-Lactamase Genes Isolated from Chronic Wounds in Ghana. Antibiotics. 2022; 11(5):689. https://doi.org/10.3390/antibiotics11050689
Chicago/Turabian StylePankok, Frederik, Stefan Taudien, Denise Dekker, Thorsten Thye, Kwabena Oppong, Charity Wiafe Akenten, Maike Lamshöft, Anna Jaeger, Martin Kaase, Simone Scheithauer, and et al. 2022. "Epidemiology of Plasmids in Escherichia coli and Klebsiella pneumoniae with Acquired Extended Spectrum Beta-Lactamase Genes Isolated from Chronic Wounds in Ghana" Antibiotics 11, no. 5: 689. https://doi.org/10.3390/antibiotics11050689