
Antimicrobial Evaluation of Various Honey Types against Carbapenemase-Producing Gram-Negative Clinical Isolates

 , ,
, ,  and
and
Abstract
:1. Introduction
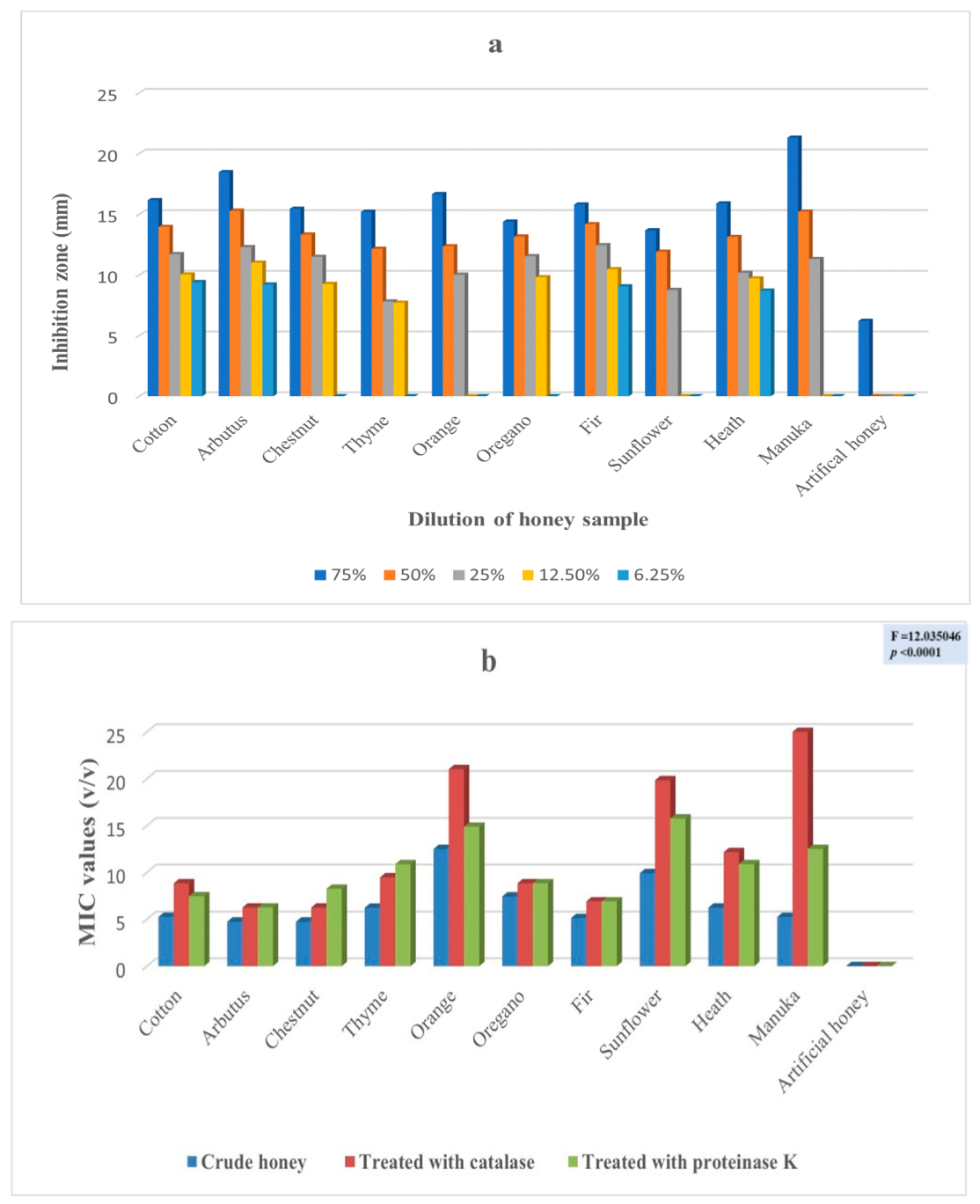
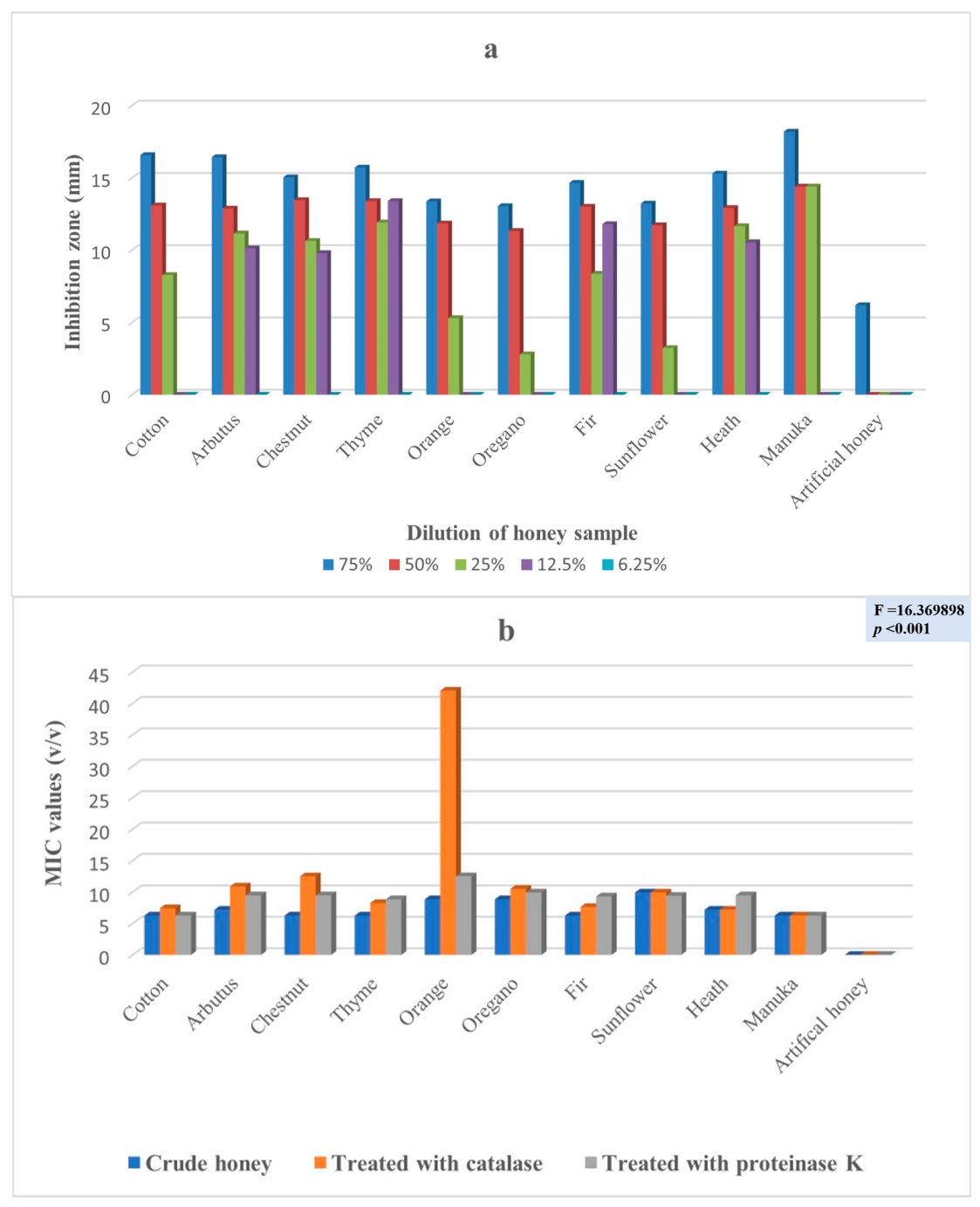
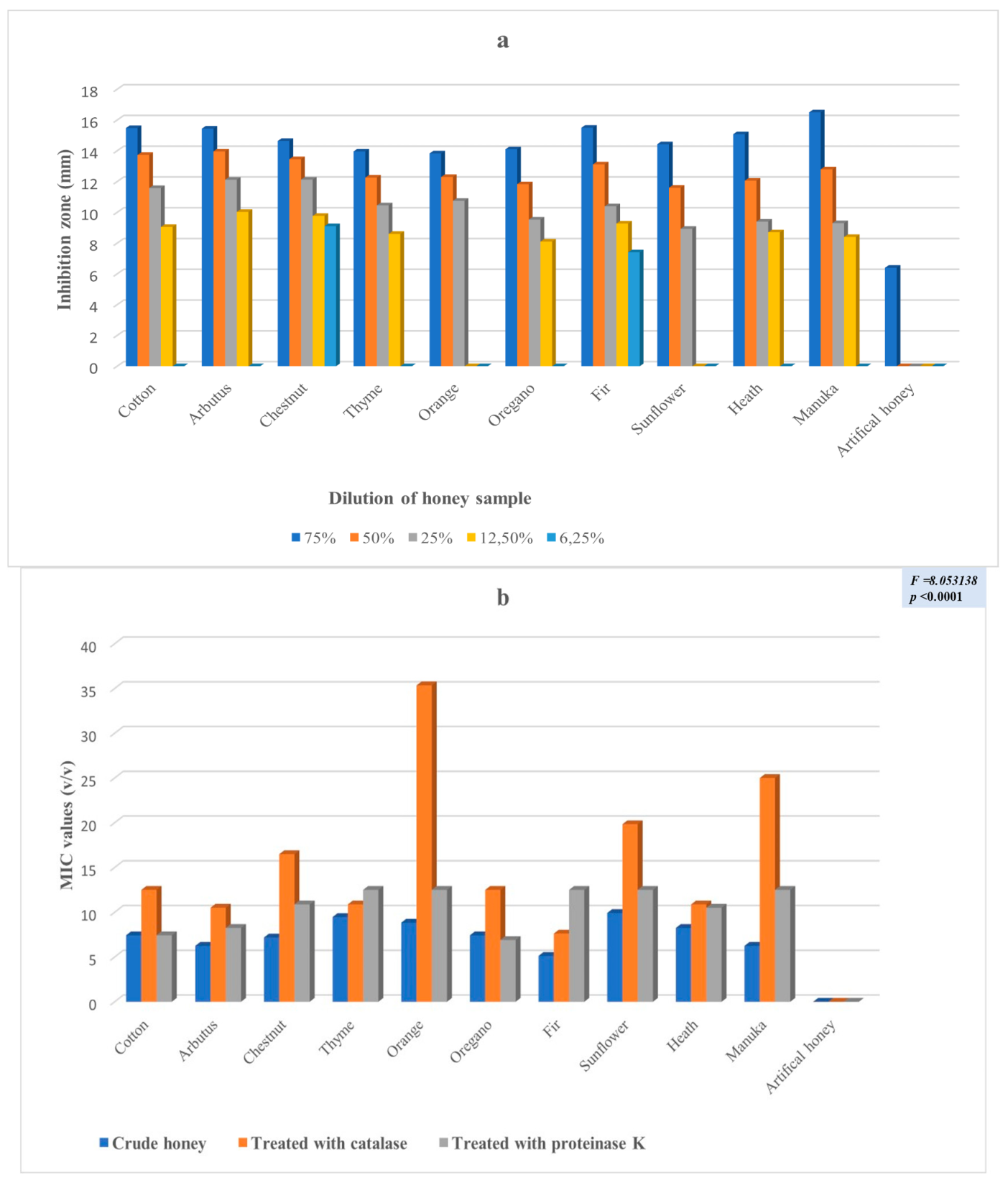
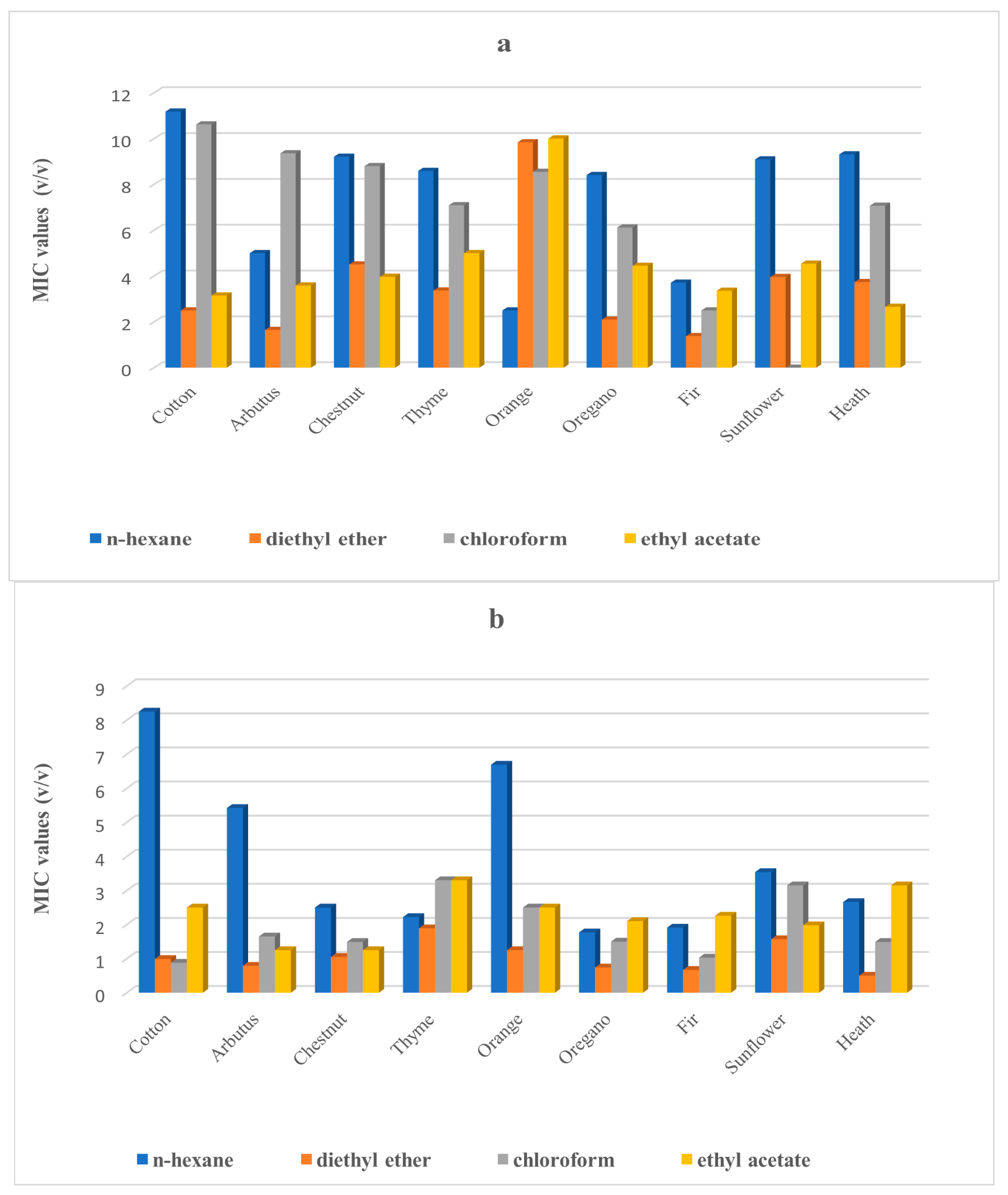
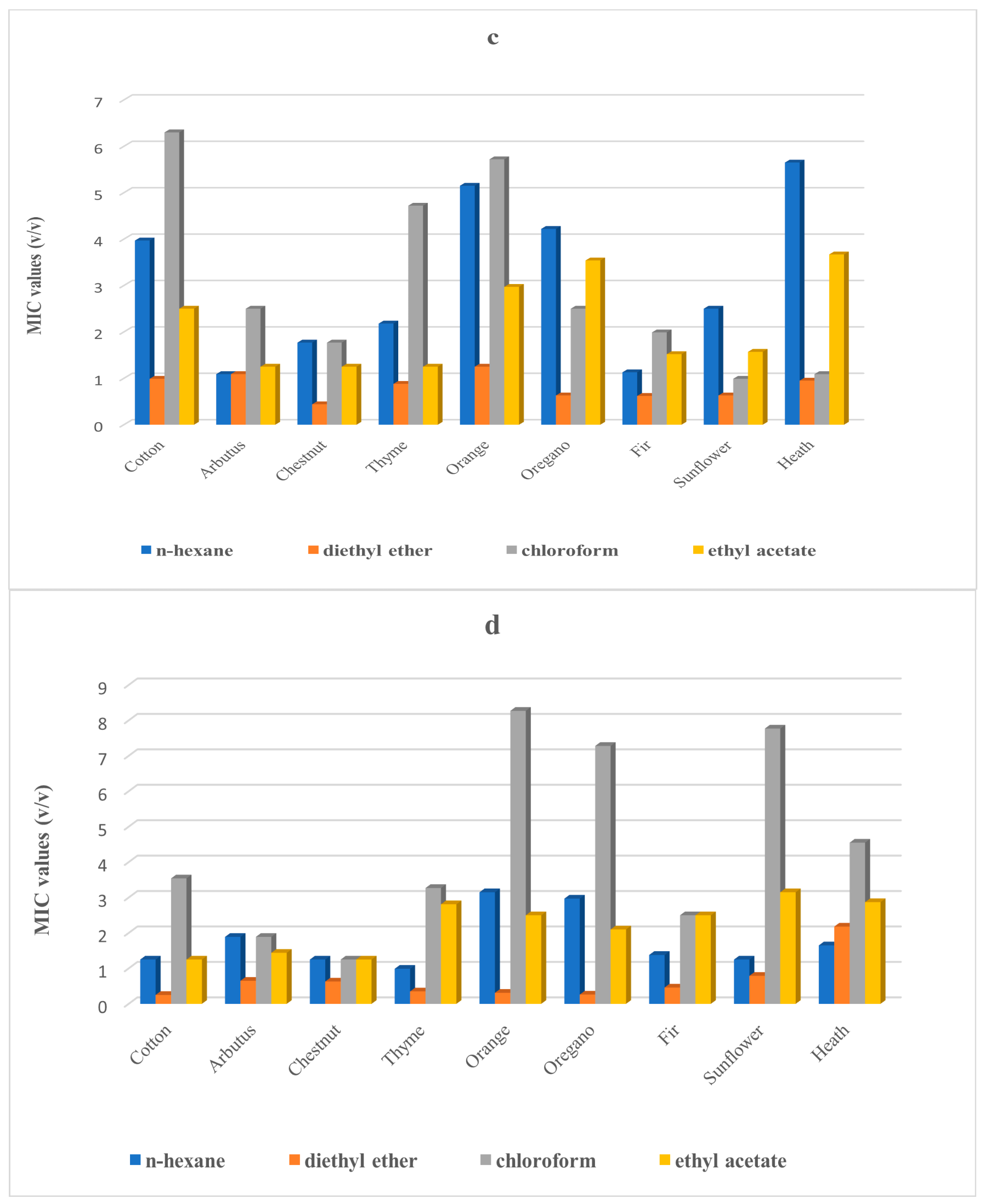
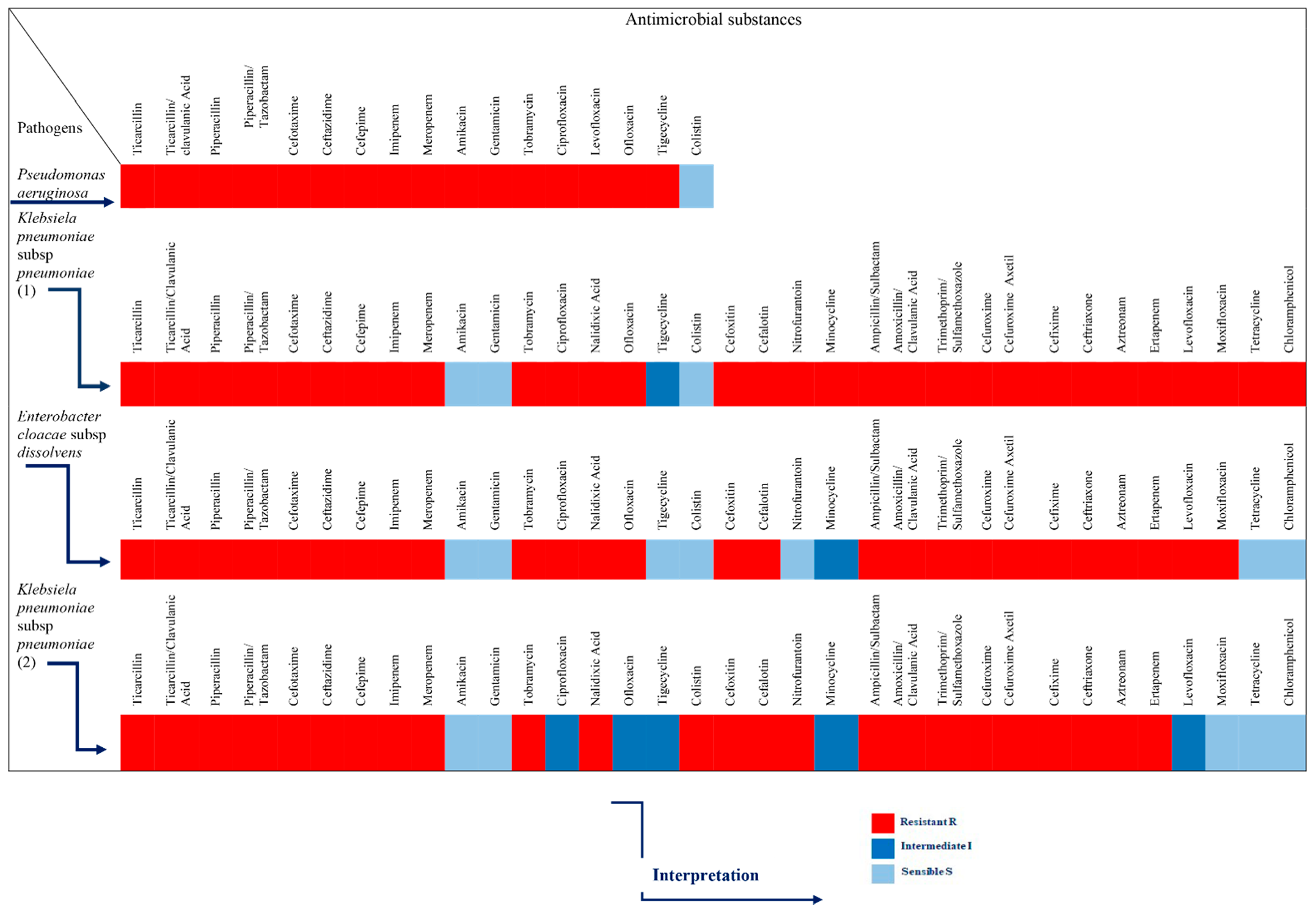
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Area—Honey Samples
4.1.1. Control Indexes of the Experimental Design
4.1.2. Determination of Physicochemical Parameters
Determination of pH
Determination of H2O2 Content
Determination of the Total Phenolic Content (TPC) (Measurement of Level of Phenolic Compounds, Which Contribute to the Antibacterial Activity of Honey)
Determination of Free, Lactonic and Total Acidity (Determination of the Acidity Factors with May Have a Potential Effect on the Expression of the Antimicrobial Activity of Honey)
Determination of the Total Flavonoid Content (TFC) (Due to the Contribution of Flavonoids in Antimicrobial Activity of Honey)
Determination of the DPPH Free Radical Scavenging Activity (Screening the Antioxidant Activity of Honey Samples through Investigation of the Overall Hydrogen or Electron Donating Activity of Single Antioxidants)
Determination of the Anti-Tyrosinase Activity (Study of Tyrosinase inhibition by Honey Samples, Factor That Potentially Enhances the Antimicrobial Activity)
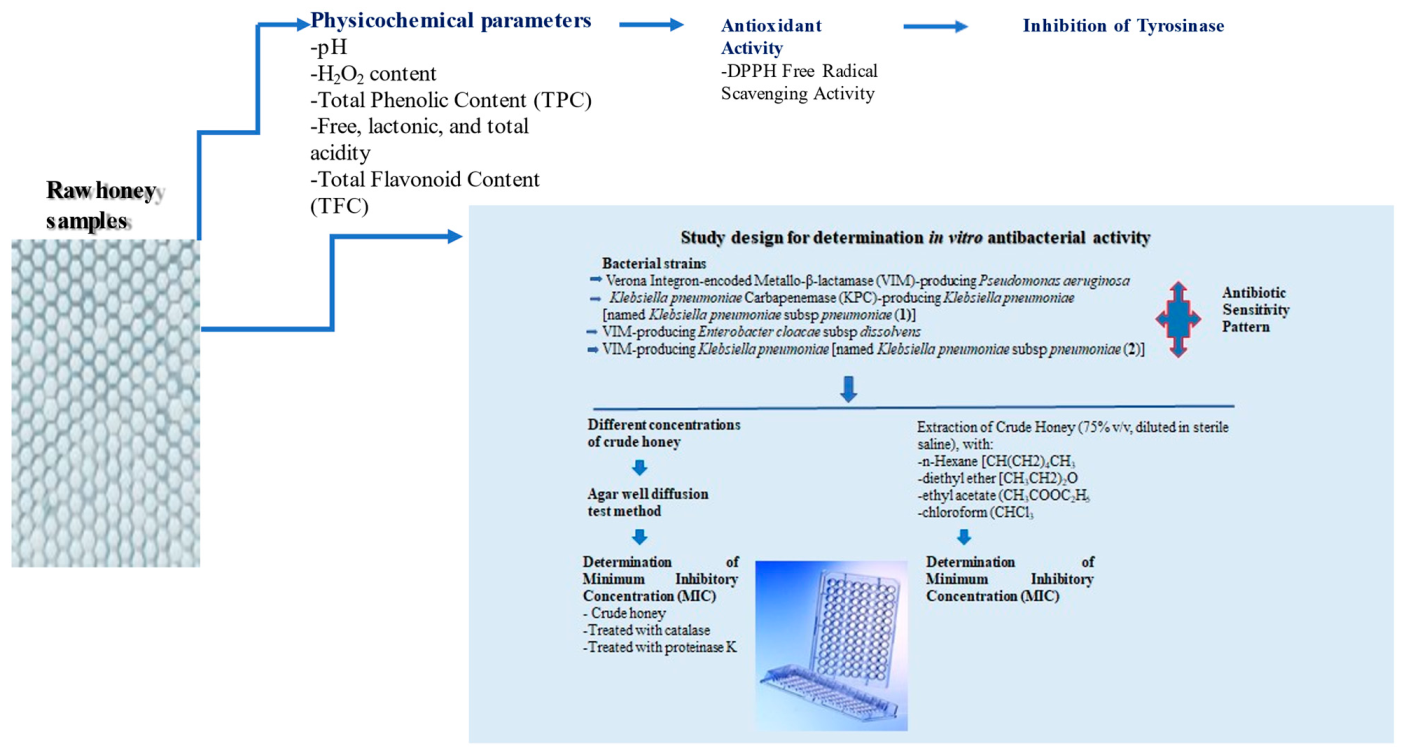
4.2. Study Design for Determination In Vitro Antibacterial Activity of Honey Samples
4.2.1. Bacterial Strains and Antibiotic Sensitivity Pattern
4.2.2. Used Solvents
4.2.3. Extraction of Crude Honey
4.2.4. Determination of the Antibacterial Activity of Crude Honey Samples
Determination of Minimum Inhibitory Concentration (MIC)
Determination of Minimum Inhibitory Concentration (MIC) Performed after Enzymatic Treatment of Honey Samples with Catalase and Proteinase K
4.2.5. Screening the Antibacterial Efficacy of Honey Extracts Antimicrobial Assay
Preliminary Assessment of Antimicrobial Activity
Minimum Inhibitory Concentration (MIC)
- -
- Points needs clarification
- (i)
- Crude honey: Honey sample (10 g) was extracted in a condenser with 50 mL of distilled water at 60 °C for more than 6 h. The obtained extract was filtered to remove particles and volume was adjusted with ultra-pure water.
- (ii)
- Dried honey: After extraction with diethyl ether, dried honey was applied in the MIC assay as following; weights extracts were diluted (400 mg/mL honey/ultrapure water solution) for 24 h, vortexed at 1500 rpm for 3 min and filtered through a 0.45 Whatman TM syringe filter (Merck, Germany).Then, serial dilutions were prepared at the following concentrations (200 mg/mL, 100 mg/mL, 50 mg/mL, 25 mg/mL, 12.5 mg/mL, 6.25 mg/mL, 3.125 mg/mL, 1.56 mg/mL, 0.78 mg/mL and 0.39 mg/mL).
Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bezirtzoglou, E.; Alexopoulos, A.; Voidarou, C. Apparent antibiotic misuse in environmental ecosystems and food. Microb. Ecol. Health Dis. 2008, 20, 197–198. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- WEF Global Risks Report 2013.pdf. Available online: https://www3.weforum.org/docs/WEF_GlobalRisks_Report_2013.pdf (accessed on 16 December 2021).
- WEF_GlobalRisks_Report_2014.pdf. Available online: https://www3.weforum.org/docs/WEF_GlobalRisks_Report_2014.pdf (accessed on 16 December 2021).
- Multidrug Antibiotic Resistance Increasing in Europe. European Centre for Disease Prevention and Control. 2012. Available online: https://www.ecdc.europa.eu/en/news-events/multidrug-antibiotic-resistance-increasing-europe (accessed on 16 December 2021).
- European Antimicrobial Resistance Surveillance Network (EARS-Net). Available online: https://www.ecdc.europa.eu/en/about-us/partnerships-and-networks/disease-and-laboratory-networks/ears-net (accessed on 16 December 2021).
- Surveillance of Antimicrobial Resistance in Europe. 2018. Available online: https://www.ecdc.europa.eu/en/publications-data/surveillance-antimicrobial-resistance-europe-2018 (accessed on 16 December 2021).
- Alternatives to Antibiotics: Why and How—National Academy of Medicine. Available online: https://nam.edu/alternatives-to-antibiotics-why-and-how/ (accessed on 16 December 2021).
- Kohler, P.; Tijet, N.; Kim, H.C.; Johnstone, J.; Edge, T.; Patel, S.N.; Seah, C.; Willey, B.; Coleman, B.; Green, K.; et al. Dissemination of Verona Integron-encoded Metallo-β-lactamase among clinical and environmental Enterobacteriaceae isolates in Ontario, Canada. Sci. Rep. 2020, 10, 18580. [Google Scholar] [CrossRef] [PubMed]
- Palacios, A.R.; Rossi, M.-A.; Mahler, G.S.; Vila, A.J. Metallo-β-Lactamase Inhibitors Inspired on Snapshots from the Catalytic Mechanism. Biomolecules 2020, 10, 854. [Google Scholar] [CrossRef]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 26, 26717. [Google Scholar] [CrossRef] [Green Version]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Genet. 2012, 11, 95–105. [Google Scholar] [CrossRef]
- Martin, K.W.; Ernst, E. Herbal medicines for treatment of fungal infections: A systematic review of controlled clinical trials. Phytomedizin zur Behandlung von Pilzinfektionen: Ubersicht und Bewertung kontrollierter klinischer Studien. Mycoses 2004, 47, 87–92. [Google Scholar] [CrossRef]
- Honey in History. Prehistory, Ancient Egypt, Ancient China. 2015. Available online: https://healthywithhoney.com/honey-in-history-prehistory-ancient-egypt-ancient-china/ (accessed on 16 December 2021).
- Mandal, M.; Mandal, S. Honey: Its medicinal property and antibacterial activity. Asian Pac. J. Trop. Biomed. 2011, 1, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Sanz, M.; Gonzalez, M.; De Lorenzo, C.; Sanz, J.; Martĺnez-Castro, I. A contribution to the differentiation between nectar honey and honeydew honey. Food Chem. 2005, 91, 313–317. [Google Scholar] [CrossRef]
- Nolan, V.C.; Harrison, J.; Cox, J.A. Dissecting the antimicrobial composition of honey. Antibiotics 2019, 8, 251. [Google Scholar] [CrossRef] [Green Version]
- Voidarou, C.; Alexopoulos, A.; Plessas, S.; Karapanou, A.; Mantzourani, I.; Stavropoulou, E.; Fotou, K.; Tzora, A.; Skoufos, I.; Bezirtzoglou, E. Antibacterial activity of different honeys against pathogenic bacteria. Anaerobe 2011, 17, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Combarros-Fuertes, P.; Fresno, J.M.; Estevinho, M.M.; Sousa-Pimenta, M.; Tornadijo, M.E.; Estevinho, L.M. Honey: Another Alternative in the Fight against Antibiotic-Resistant Bacteria? Antibiotics 2020, 9, 774. [Google Scholar] [CrossRef] [PubMed]
- Molan, P.C. The Evidence Supporting the Use of Honey as a Wound Dressing. Int. J. Low. Extrem. Wounds 2006, 5, 40–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, W.-J.; Sit, N.-W.; Ooi, P.A.-C.; Ee, K.-Y.; Lim, T.-M. The Antibacterial Potential of Honeydew Honey Produced by Stingless Bee (Heterotrigona itama) against Antibiotic Resistant Bacteria. Antibiotics 2020, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Voidarou, C.; Rozos, G.; Alexopoulos, A.; Plessas, S.; Mantzourani, I.; Stavropoulou, E.; Tzora, A.; Bezirtzoglou, E. In Vitro Screening Potential Antibacterial Properties of the Greek Oregano Honey against Clinical Isolates of Helicobacter pylori. Foods 2021, 10, 1568. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, S.; Jurendic, T.; Sieber, R.; Gallmann, P. Honey for Nutrition and Health: A Review. J. Am. Coll. Nutr. 2008, 27, 677–689. [Google Scholar] [CrossRef]
- John-Isa, J.F.; Adebolu, T.T.; Oyetayo, V.O. Antibacterial Effects of Honey in Nigeria on Selected Diarrhoeagenic Bacteria. South Asian J. Res. Microbiol. 2019, 3, 1–11. [Google Scholar] [CrossRef]
- Escuredo, O.; Silva, L.R.; Valentão, P.; Seijo, M.C.; Andrade, P.B. Assessing Rubus honey value: Pollen and phenolic compounds content and antibacterial capacity. Food Chem. 2012, 130, 671–678. [Google Scholar] [CrossRef]
- Matzen, R.D.; Leth-Espensen, J.Z.; Jansson, T.; Nielsen, D.S.; Lund, M.; Matzen, S. The Antibacterial Effect In Vitro of Honey Derived from Various Danish Flora. Dermatol. Res. Pract. 2018, 2018, 7021713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, M.; McBride, M.; Dahiya, D.; Owusu-Apenten, R.; Nigam, P.S. Antibacterial activity of Manuka honey and its components: An overview. AIMS Microbiol. 2018, 4, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Molan, P.C. The antibacterial activity of honey: 1. The nature of the antibacterial activity. Bee World 2015, 73, 5–28. [Google Scholar] [CrossRef]
- Makena, A.; Düzgün, A.Ö.; Brem, J.; McDonough, M.A.; Rydzik, A.M.; Abboud, M.I.; Saral, A.; Çiçek, A.Ç.; Sandalli, C.; Schofield, C.J. Comparison of Verona Integron-Borne Metallo-β-Lactamase (VIM) Variants Reveals Differences in Stability and Inhibition Profiles. Antimicrob. Agents Chemother. 2016, 60, 1377–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, G.T. Continuous Evolution: Perspective on the Epidemiology of Carbapenemase Resistance among Enterobacterales and Other Gram-Negative Bacteria. Infect. Dis. Ther. 2021, 10, 75–92. [Google Scholar] [CrossRef]
- Suay-García, B.; Pérez-Gracia, M.T. Present and Future of Carbapenem-resistant Enterobacteriaceae (CRE) Infections. Antibiotics 2019, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Wasihun, A.G.; Kasa, B.G. Evaluation of antibacterial activity of honey against multidrug resistant bacteria in Ayder Referral and Teaching Hospital, Northern Ethiopia. SpringerPlus 2016, 5, 842–850. [Google Scholar] [CrossRef] [Green Version]
- Scepankova, H.; Combarros-Fuertes, P.; Fresno, J.M.; Tornadijo, M.E.; Dias, M.S.; Pinto, C.A.; Saraiva, J.A.; Estevinho, L.M. Role of Honey in Advanced Wound Care. Molecules 2021, 26, 4784. [Google Scholar] [CrossRef]
- Hayes, G.; Wright, N.; Gardner, S.L.; Telzrow, C.; Wommack, A.J.; Vigueira, P.A. Manuka honey and methylglyoxal increase the sensitivity of Staphylococcus aureus to linezolid. Lett. Appl. Microbiol. 2018, 66, 491–495. [Google Scholar] [CrossRef]
- Cooper, R.A.; Molan, P.C.; Harding, K.G. The sensitivity to honey of Gram-positive cocci of clinical significance isolated from wounds. J. Appl. Microbiol. 2002, 93, 857–863. [Google Scholar] [CrossRef]
- Almasaudi, S. The antibacterial activities of honey. Saudi J. Biol. Sci. 2021, 28, 2188–2196. [Google Scholar] [CrossRef]
- Frydman, G.H.; Olaleye, D.; Annamalai, D.; Layne, K.; Yang, I.; Kaafarani, H.M.A.; Fox, J.G. Manuka honey microneedles for enhanced wound healing and the prevention and/or treatment of Methicillin-resistant Staphylococcus aureus (MRSA) surgical site infection. Sci. Rep. 2020, 10, 13229. [Google Scholar] [CrossRef]
- Al Refaey, H.R.; Newairy, A.-S.A.; Wahby, M.M.; Albanese, C.; Elkewedi, M.; Choudhry, M.U.; Sultan, A.S. Manuka honey enhanced sensitivity of HepG2, hepatocellular carcinoma cells, for Doxorubicin and induced apoptosis through inhibition of Wnt/β-catenin and ERK1/2. Biol. Res. 2021, 54, 16. [Google Scholar] [CrossRef]
- Liu, M.Y.; Cokcetin, N.N.; Lu, J.; Turnbull, L.; Carter, D.; Whitchurch, C.B.; Harry, E. Rifampicin-Manuka Honey Combinations Are Superior to Other Antibiotic-Manuka Honey Combinations in Eradicating Staphylococcus aureus Biofilms. Front. Microbiol. 2018, 8, 2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, R.; Burton, N.; Cooper, R. Manuka honey inhibits cell division in methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2011, 66, 2536–2542. [Google Scholar] [CrossRef] [Green Version]
- Girma, A.; Seo, W.; She, R.C. Antibacterial activity of varying UMF-graded Manuka honeys. PLoS ONE 2019, 14, e0224495. [Google Scholar] [CrossRef] [Green Version]
- Cremers, N.; Belas, A.; Costa, S.S.; Couto, I.; De Rooster, H.; Pomba, C. In vitro antimicrobial efficacy of two medical grade honey formulations against common high-risk meticillin-resistant staphylococci and Pseudomonas spp. pathogens. Vet. Dermatol. 2019, 31, 90–96. [Google Scholar] [CrossRef]
- Idris, A.R.; Afegbua, S.L. Single and joint antibacterial activity of aqueous garlic extract and Manuka honey on extended-spectrum beta-lactamase-producing Escherichia coli. Trans. R. Soc. Trop. Med. Hyg. 2017, 111, 472–478. [Google Scholar] [CrossRef]
- O’Grady, F.; Lambert, H.P.; Finch, R.G.; Greenwood, D. Antibiotic, and Chemotherapy: Anti-Infective Agents and Their Use in Therapy, 7th ed.; Churchill Livingstone: London, UK, 1997. [Google Scholar]
- Khan, F.R.; Abadin, Z.U.; Rauf, N. Honey: Nutritional and medicinal value. Int. J. Clin. Pract. 2007, 61, 1705–1707. [Google Scholar] [CrossRef]
- Al Somal, N.; Coley, K.E.; Molan, P.C.; Hancock, B.M. Susceptibility of Helicobacter pylori to the antibacterial activity of manuka honey. J. R. Soc. Med. 1994, 87, 9–12. [Google Scholar]
- Oelschlaegel, S.; Gruner, M.; Wang, P.-N.; Boettcher, A.; Koelling-Speer, I.; Speer, K. Classification and Characterization of Manuka Honeys Based on Phenolic Compounds and Methylglyoxal. J. Agric. Food Chem. 2012, 60, 7229–7237. [Google Scholar] [CrossRef] [PubMed]
- Israili, Z.H. Antimicrobial Properties of Honey. Am. J. Ther. 2014, 21, 304–323. [Google Scholar] [CrossRef] [PubMed]
- Estevinho, L.; Pereira, A.P.; Moreira, L.F.; Dias, L.; Pereira, E.L. Antioxidant and antimicrobial effects of phenolic compounds extracts of Northeast Portugal honey. Food Chem. Toxicol. 2008, 46, 3774–3779. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Scholz, M.B.; Júnior, A.Q.; Delamuta, B.H.; Nakamura, J.M.; Baudraz, M.C.; Reis, M.O.; Kato, T.; Pedrão, M.R.; Dias, L.F.; dos Santos, D.T.R.; et al. Indication of the geographical origin of honey using its physicochemical characteristics and multivariate analysis. J. Food Sci. Technol. 2020, 57, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- Ananias, K.R.; De-Melo, A.A.M.; De Moura, C.J. Analysis of moisture content, acidity and contamination by yeast and molds in Apis mellifera L. honey from central Brazil. Braz. J. Microbiol. 2013, 44, 679–683. [Google Scholar] [CrossRef] [Green Version]
- Barra, M.P.G.; Ponce-Díaz, M.C.; Venegas-Gallegos, C. Volatile Compounds in Honey Produced in the Central Valley of Ñuble Province, Chile. Chil. J. Agric. Res. 2010, 70, 75–84. [Google Scholar] [CrossRef]
- Mondragón-Cortez, P.; Ulloa, J.A.; Rosas-Ulloa, P.; Rodríguez-Rodríguez, R.; Resendiz Vázquez, J.A. Physicochemical characterization of honey from the West region of México. CyTA J. Food 2013, 11, 7–13. [Google Scholar] [CrossRef]
- Samarghandian, S.; Farkhondeh, T.; Samini, F. Honey and health: A review of recent clinical research. Pharmacogn. Res. 2017, 9, 121–127. [Google Scholar] [CrossRef]
- Richter, G.; Heinecker, H. Conversion of Glucose into Gluconic Acid by Means of Immobilized Glucose Oxidase. Starch-Stärke 1979, 31, 418–422. [Google Scholar] [CrossRef]
- Weston, R.J. The contribution of catalase and other natural products to the antibacterial activity of honey: A review. Food Chem. 2000, 71, 235–239. [Google Scholar] [CrossRef]
- Brudzynski, K.; Abubaker, K.; Miotto, D. Unraveling a mechanism of honey antibacterial action: Polyphenol/H2O2-induced oxidative effect on bacterial cell growth and on DNA degradation. Food Chem. 2012, 133, 329–336. [Google Scholar] [CrossRef]
- Mato, I.; Huidobro, J.F.; Simal-Lozano, J.; Sancho, M. Significance of Nonaromatic Organic Acids in Honey. J. Food Prot. 2003, 66, 2371–2376. [Google Scholar] [CrossRef]
- Bang, L.M.; Buntting, C.; Molan, P. The Effect of Dilution on the Rate of Hydrogen Peroxide Production in Honey and Its Implications for Wound Healing. J. Altern. Complement. Med. 2003, 9, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, T.; Rao, P.V. Free acidity measurement—A review. Talanta 2014, 118, 162–171. [Google Scholar] [CrossRef]
- Ribeiro, I.A.C.; Bronze, M.; Castro, M.F.; Ribeiro, M.H.L. Selective recovery of acidic and lactonic sophorolipids from culture broths towards the improvement of their therapeutic potential. Bioprocess Biosyst. Eng. 2016, 39, 1825–1837. [Google Scholar] [CrossRef]
- Zhang, X.; Ashby, R.D.; Solaiman, D.K.; Liu, Y.; Fan, X. Antimicrobial activity and inactivation mechanism of lactonic and free acid sophorolipids against Escherichia coli O157:H7. Biocatal. Agric. Biotechnol. 2017, 11, 176–182. [Google Scholar] [CrossRef]
- Tewari, J.; Irudayaraj, J. Quantification of Saccharides in Multiple Floral Honeys Using Fourier Transform Infrared Microattenuated Total Reflectance Spectroscopy. J. Agric. Food Chem. 2004, 52, 3237–3243. [Google Scholar] [CrossRef]
- Tseng, J.-M.; Huang, J.-R.; Huang, H.-C.; Tzen, J.T.C.; Chou, W.-M.; Peng, C.-C. Facilitative production of an antimicrobial peptide royalisin and its antibody via an artificial oil-body system. Biotechnol. Prog. 2010, 27, 153–161. [Google Scholar] [CrossRef]
- Lachman, J.; Orsák, M.; Hejtmánková, A.; Kovářová, E. Evaluation of antioxidant activity and total phenolics of selected Czech honeys. LWT-Food Sci. Technol. 2010, 43, 52–58. [Google Scholar] [CrossRef]
- Albaridi, N.A. Antibacterial Potency of Honey. Int. J. Microbiol. 2019, 2019, 2464507. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Manna, P.P.; Zhang, J.; Bravo Lamas, L.; Martínez Flórez, S.; Agudo Toyos, P.; et al. Phenolic Compounds in Honey and Their Associated Health Benefits: A Review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, H.; Maeda, M.; Okubo, S.; Shimamura, T. Role of Hydrogen Peroxide in Bactericidal Action of Catechin. Biol. Pharm. Bull. 2004, 27, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Hu, J.; Hu, D.; Yang, X. A Role of Gallic Acid in Oxidative Damage Diseases: A Comprehensive Review. Nat. Prod. Commun. 2019, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Collins, W.; Lowen, N.; Blake, D.J. Caffeic Acid Esters Are Effective Bactericidal Compounds against Paenibacillus larvae by Altering Intracellular Oxidant and Antioxidant Levels. Biomolecules 2019, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial Activity and Mechanism of Action of Chlorogenic Acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef]
- Kwakman, P.H.S.; te Velde, A.A.; de Boer, L.; Speijer, D.; Vandenbroucke-Grauls, M.J.C.; Zaat, S.A.J. How honey kills bacteria. FASEB J. 2010, 24, 2576–2582. [Google Scholar] [CrossRef] [Green Version]
- DellaGreca, M.; De Tommaso, G.; Salvatore, M.M.; Nicoletti, R.; Becchimanzi, A.; Iuliano, M.; Andolfi, A. The Issue of Misidentification of Kojic Acid with Flufuran in Aspergillus flavus. Molecules 2019, 24, 1709. [Google Scholar] [CrossRef] [Green Version]
- Szweda, P. Honeybee products as an alternative or complement to classical antibiotics (Editorial). Antibiotics 2021, 10, 234. [Google Scholar] [CrossRef]
- Şöhretoğlu, D.; Sari, S.; Barut, B.; Özel, A. Tyrosinase inhibition by some flavonoids: Inhibitory activity, mechanism by in vitro and in silico studies. Bioorganic Chem. 2018, 81, 168–174. [Google Scholar] [CrossRef]
- Kim, D.; Park, J.; Kim, J.; Han, C.; Yoon, J.; Kim, N.; Seo, J.; Lee, C. Flavonoids as Mushroom Tyrosinase Inhibitors: A Fluorescence Quenching Study. J. Agric. Food Chem. 2006, 54, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Mira, L.; Fernandez, M.T.; Santos, M.; Rocha, R.; Florêncio, M.H.; Jennings, K.R. Interactions of Flavonoids with Iron and Copper Ions: A Mechanism for their Antioxidant Activity. Free Radic. Res. 2002, 36, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Říha, M.; Karlíčková, J.; Filipský, T.; Jahodář, L.; Hrdina, R.; Mladěnka, P. In vitro copper-chelating properties of flavonoids. Free Radic. Biol. Med. 2014, 75, S46. [Google Scholar] [CrossRef] [PubMed]
- Weston, R.J.; Brocklebank, L.K.; Lu, Y. Identification, and quantitative levels of antibacterial components of some New Zealand manuka honeys. Food Chem. 2000, 70, 427–435. [Google Scholar] [CrossRef]
- Combarros-Fuertes, P.; Estevinho, L.M.; Teixeira-Santos, R.; Rodrigues, A.G.; Pina-Vaz, C.; Fresno, J.M.; Tornadijo, M.E. Antibacterial Action Mechanisms of Honey: Physiological Effects of Avocado, Chestnut, and Polyfloral Honey upon Staphylococcus aureus and Escherichia coli. Molecules 2020, 25, 1252. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.-T.T.; Rahman, R.A.; Gan, S.H.; Halim, A.S.; Hassan, S.A.; Sulaiman, S.A.; Bs, K.-K. The antibacterial properties of Malaysian tualang honey against wound and enteric microorganisms in comparison to manuka honey. BMC Complement. Altern. Med. 2009, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef]
- Saeedi, M.; Eslamifar, M.; Khezri, K. Kojic acid applications in cosmetic and pharmaceutical preparations. Biomed. Pharmacother. 2019, 110, 582–593. [Google Scholar] [CrossRef]
- Majtan, J.; Bucekova, M.; Kafantaris, I.; Szweda, P.; Hammer, K.; Mossialos, D. Honey antibacterial activity: A neglected aspect of honey quality assurance as functional food. Trends Food Sci. Technol. 2021, 118, 870–886. [Google Scholar] [CrossRef]
- Grecka, K.; Xiong, Z.R.; Chen, H.; Pełka, K.; Worobo, R.W.; Szweda, P. Effect of Ethanol Extracts of Propolis (EEPs) against Staphylococcal Biofilm—Microscopic Studies. Pathogens 2020, 9, 646. [Google Scholar] [CrossRef]
- Grecka, K.; Szweda, P. Synergistic Effects of Propolis Combined with 2-Phenoxyethanol and Antipyretics on the Growth of Staphylococcus aureus. Pharmaceutics 2021, 13, 215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-L.; Guo, Y.-S.; Wang, C.-H.; Li, G.-Q.; Xu, J.-J.; Chung, H.Y.; Wang, G.C. Phenolic compounds from Origanum vulgare and their antioxidant and antiviral activities. Food Chem. 2014, 152, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Lusby, P.E.; Coombes, A.L.; Wilkinson, J.M. Bactericidal Activity of Different Honeys against Pathogenic Bacteria. Arch. Med. Res. 2005, 36, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, M.K.; Sonter, S.; Mishra, S.; Patel, D.K.; Singh, P.K. Antioxidant, antibacterial activity, and phytochemical characterization of Carica papaya flowers. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 23. [Google Scholar] [CrossRef]
- Brudzynski, K. Honey as an Ecological Reservoir of Antibacterial Compounds Produced by Antagonistic Microbial Interactions in Plant Nectars, Honey and Honey Bee. Antibiotics 2021, 10, 551. [Google Scholar] [CrossRef]
- Pełka, K.; Worobo, R.; Walkusz, J.; Szweda, P. Bee Pollen and Bee Bread as a Source of Bacteria Producing Antimicrobials. Antibiotics 2021, 10, 713. [Google Scholar] [CrossRef]
- Pajor, M.; Worobo, R.W.; Milewski, S.; Szweda, P. The Antimicrobial Potential of Bacteria Isolated from Honey Samples Produced in the Apiaries Located in Pomeranian Voivodeship in Northern Poland. Int. J. Environ. Res. Public Health 2018, 15, 2002. [Google Scholar] [CrossRef] [Green Version]
- Pasupuleti, V.R.; Sammugam, L.; Ramesh, N.; Gan, S.H. Honey, Propolis, and Royal Jelly: A Comprehensive Review of Their Biological Actions and Health Benefits. Oxidative Med. Cell. Longev. 2017, 2017, 1259510. [Google Scholar] [CrossRef]
- Dixon, B. Bacteria can’t resist honey. Lancet Infect. Dis. 2003, 3, 116. [Google Scholar] [CrossRef]
- Bovo, S.; Utzeri, V.J.; Ribani, A.; Cabbri, R.; Fontanesi, L. Shotgun sequencing of honey DNA can describe honey bee derived environmental signatures and the honey bee hologenome complexity. Sci. Rep. 2020, 10, 9279. [Google Scholar] [CrossRef]
- Sabree, Z.L.; Hansen, A.; Moran, N.A. Independent Studies Using Deep Sequencing Resolve the Same Set of Core Bacterial Species Dominating Gut Communities of Honey Bees. PLoS ONE 2012, 7, e41250. [Google Scholar] [CrossRef] [Green Version]
- Brudzynski, K.; Sjaarda, C. Honey Glycoproteins Containing Antimicrobial Peptides, Jelleins of the Major Royal Jelly Protein 1, Are Responsible for the Cell Wall Lytic and Bactericidal Activities of Honey. PLoS ONE 2015, 10, e0120238. [Google Scholar] [CrossRef] [Green Version]
- Wakeel, A.; Jan, S.A.; Ullah, I.; Shinwari, Z.K.; Xu, M. Solvent polarity mediates phytochemical yield and antioxidant capacity of Isatis tinctoria. PeerJ 2019, 7, e7857. [Google Scholar] [CrossRef] [Green Version]
- Manyi-Loh, C.E.; Ndip, R.N.; Clarke, A.M. Volatile Compounds in Honey: A Review on Their Involvement in Aroma, Botanical Origin Determination and Potential Biomedical Activities. Int. J. Mol. Sci. 2011, 12, 9514–9532. [Google Scholar] [CrossRef] [Green Version]
- Al-Waili, N.; Al Ghamdi, A.; Ansari, M.J.; Al-Attal, Y.; Al-Mubarak, A.; Salom, K. Differences in Composition of Honey Samples and Their Impact on the Antimicrobial Activities against Drug Multiresistant Bacteria and Pathogenic Fungi. Arch. Med Res. 2013, 44, 307–316. [Google Scholar] [CrossRef]
- Stagos, D.; Soulitsiotis, N.; Tsadila, C.; Papaeconomou, S.; Arvanitis, C.; Ntontos, A.; Karkanta, F.; Adamou-Androulaki, S.; Petrotos, K.; Spandidos, D.; et al. Antibacterial and antioxidant activity of different types of honey derived from Mount Olympus in Greece. Int. J. Mol. Med. 2018, 42, 726–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voidarou, C.; Antoniadou, M.; Rozos, G.; Alexopoulos, A.; Giorgi, E.; Tzora, A.; Skoufos, I.; Varzakas, T.; Bezirtzoglou, E. An In Vitro Study of Different Types of Greek Honey as Potential Natural Antimicrobials against Dental Caries and Other Oral Pathogenic Microorganisms. Case Study Simulation of Oral Cavity Conditions. Appl. Sci. 2021, 11, 6318. [Google Scholar] [CrossRef]
- Gkoutzouvelidou, M.; Panos, G.; Xanthou, M.; Papachristoforou, A.; Giaouris, E. Comparing the Antimicrobial Actions of Greek Honeys from the Island of Lemnos and Manuka Honey from New Zealand against Clinically Important Bacteria. Foods 2021, 10, 1402. [Google Scholar] [CrossRef]
- Godocikova, J.; Bugarova, V.; Kast, C.; Majtan, V.; Majtan, J. Antibacterial potential of Swiss honeys and characterisation of their bee-derived bioactive compounds. J. Sci. Food Agric. 2020, 100, 335–342. [Google Scholar] [CrossRef]
- Download.pdf. Available online: https://www.fda.gov/media/73207/download (accessed on 16 January 2022).
- Bucekova, M.; Jardekova, L.; Juricova, V.; Bugarova, V.; Di Marco, G.; Gismondi, A.; Leonardi, D.; Farkasovska, J.; Godocikova, J.; Laho, M.; et al. Antibacterial Activity of Different Blossom Honeys: New Findings. Molecules 2019, 24, 1573. [Google Scholar] [CrossRef] [Green Version]
- Bucekova, M.; Bugarova, V.; Godocikova, J.; Majtan, J. Demanding New Honey Qualitative Standard Based on Antibacterial Activity. Foods 2020, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, H.A.; Alsabehi, R.; Boukraâ, L.; Abde-Llah, F.; Bellik, Y.; Bakhotmah, B.A. Antibacterial and Antioxidant Potency of Floral Honeys from Different Botanical and Geographical Origins. Molecules 2012, 17, 10540–10549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Farsi, M.; Al-Amri, A.; Al-Hadhrami, A.; Al-Belushi, S. Color, flavonoids, phenolics and antioxidants of Omani honey. Heliyon 2018, 4, e00874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AOAC International. AOAC: Official Methods of Analysis (Volume 1). 1990. Available online: http://archive.org/details/gov.law.aoac.methods.1.1990 (accessed on 16 January 2022).
- Terrab, A.; Recamales, A.F.; Hernanz, D.; Heredia, F.J. Characterisation of Spanish thyme honeys by their physicochemical characteristics and mineral contents. Food Chem. 2004, 88, 537–542. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Khalil, M.; Sulaiman, S.; Gan, S. Advances in the analytical methods for determining the antioxidant properties of honey: A review. Afr. J. Tradit. Complement. Altern. Med. 2011, 9, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Jantakee, K.; Tragoolpua, Y. Activities of different types of Thai honey on pathogenic bacteria causing skin diseases, tyrosinase enzyme and generating free radicals. Biol. Res. 2015, 48, 4. [Google Scholar] [CrossRef] [Green Version]
- Potduang, B.; Meeploy, M.; Giwanon, R.; Benmart, Y.; Kaewduang, M.; Supatanakul, W. Biological Activities of Asparagus racemosus. Afr. J. Tradit. Complement. Altern. Med. 2008, 5, 230–237. [Google Scholar] [CrossRef]
- Weldhagen, G.F.; Poirel, L.; Nordmann, P. Ambler Class A Extended-Spectrum β-Lactamases in Pseudomonas aeruginosa: Novel Developments and Clinical Impact. Antimicrob. Agents Chemother. 2003, 47, 2385–2392. [Google Scholar] [CrossRef] [Green Version]
- Tsakris, A.; Poulou, A.; Pournaras, S.; Voulgari, E.; Vrioni, G.; Themeli-Digalaki, K.; Petropoulou, D.; Sofianou, D. A simple phenotypic method for the differentiation of metallo-β-lactamases and class A KPC carbapenemases in Enterobacteriaceae clinical isolates. J. Antimicrob. Chemother. 2010, 65, 1664–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manyi-Loh, C.; Clarke, A.M.; Ndip, R.N. An overview of honey: Therapeutic properties and contribution in nutrition and human health. Afr. J. Microbiol. Res. 2011, 5, 844–852. [Google Scholar] [CrossRef]
- Hamilton-Miller, J.M.; Gargan, R.A. Rapid screening for urease inhibitors. Investig. Urol. 1979, 16, 327–328. [Google Scholar]
- Patton, T.; Barrett, J.; Brennan, J.; Moran, N. Use of a spectrophotometric bioassay for determination of microbial sensitivity to manuka honey. J. Microbiol. Methods 2006, 64, 84–95. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Honey Samples Based on Botanical Origin | Physicochemical Parameters | ||||||
|---|---|---|---|---|---|---|---|
| pH | H2O2 mM in 40% Aqueous Honey Solution | Free Acidity | Lactonic Acidity | TPC (mg GAE/100 g of Honey) | TFC mg Quercetin Equivalents (CE)/100 g of Honey | DPPH (mg GAE/kg of Honey) | |
| Cotton | 3.73 ± 0.33 a | 0.84 ± 0.12 a | 17.75 ± 2.48 a | 8.30 ± 4.07 | 99.7 ± 39.89 a | 3.33 ± 0.93 a | 1.9 ± 1.22 a |
| Arbutus | 3.32 ± 0.22 b | 0.97 ± 0.15 b | 42.24 ± 6.25 a,b | 11.88 ± 4.08 | 52.32 ± 19.21 b | 4.22 ± 0.64 b | 2.78 ± 0.66 b |
| Chestnut | 3.66 ± 0.49 c | 1.16 ± 0.06 c | 42.80 ± 14.01 a,c | 15.44 ± 4.18 | 52.86 ± 23.91 c | 3.28 ± 1.95 c | 2.02 ± 1.33 c |
| Thyme | 3.48 ± 0.42 d | 0.59 ± 0.08 d | 37.60 ± 11.97 a,d | 11.14 ± 1.97 | 70.7 ± 25.18 | 2.06 ± 0.80 b,d | 1.5 ± 0.69 d |
| Orange | 3.70 ± 0.18 e | 0.36 ± 0.06 e | 30.48 ± 5.19 e | 9.23 ± 0.66 | 38.3 ± 7.16 a,d | 1.1 ± 0.18 a, b, c, e | 0.9 ± 0.25 b,e |
| Oregano | 3.23 ± 0.26 f | 1.39 ± 0.31 f | 42.18 ± 7.26 a, f | 12.45 ± 1.50 | 46.00 ± 14.80 a,e | 3.58 ± 0.77 e,f | 2.65 ± 0.94 e,f |
| Fir | 4.87 ± 0.21 a,b,c,d,e,f,g,j | 3.02 ± 1.78 a,b,c,d.e,f,g,j,k | 18.39 ± 4.66 b,c,d,f,g | 11.04 ± 3.54 | 99.1 ± 34.43 b,c,d,e,f | 4.03 ± 0.73 d,e,g | 3.19 ± 0.60 e,g |
| Sunflower | 3.93 ± 0.25 b,h,f | 0.36 ± 0.05 g | 36.93 ± 3.69 a,g,j | 9.70 ± 0.51 | 30.6 ± 1.71 a, f,g | 0.92 ± 0.14 a,b,c,f,g,h | 1.33 ± 0.48 d,g,h |
| Heath | 3.22 ± 0.13 j | 1.29 ± 0.11 j | 15.28 ± 2.52 b,c,d, e,f,j | 7.84 ± 2.14 | 43.66 ± 9.87 a,f | 1.36 ± 0.31 a,b,c,f,g,j | 1.98 ± 0.99 j |
| Manuka honey | 4.10 ± 0.15 b,d,f,j | 1.27 ± 0.74 k | 15.2 ± 0.20 b,c,d,e,f,j | 7.10 ± 0.20 | 88.71 ± 0.3 d,g | 4.1 ± 0.80 d,e,h,j | 5.1 ± 0.8 a,b,c,d, e,f,g,h,j |
| Honey Samples Based on Botanical Origin | Tyrosinase Inhibition | Kojic Acid |
|---|---|---|
| Cotton | 56.21 ± 3.02 a | 31.24 ± 1.91 a |
| Arbutus | 48.66 ± 9.08 b | 18.74 ± 4.56 b |
| Chestnut | 58.28 ± 4.61 c | 40.28 ± 2.64 b,c |
| Thyme | 50.04 ± 1.87 d | 14.31 ± 0.76 a,c,d |
| Orange | 44.39 ± 2.90 c,e | 7.84 ± 1.93 a,c,e |
| Oregano | 81.92 ± 4.90 a,b,c,d,e,f | 86.21 ± 14.48 a,b,c,e,f |
| Fir | 56.46 ± 12.56 f,g | 19.66 ± 9.42 c,f,g |
| Sunflower | 29.40 ± 2.85 a,b,c,d,e,f,g,h | 4.99 ± 1.12 a,b,c,f,g,h |
| Heath | 40.09 ± 2.96 a,c,f,g,j | 7.32 ± 2.54 a,c,f,g,j |
| Manuka honey | 85.11 ± 4.33 a,b,c,d,e,g,h,j | 210.15 ± 12.26 a,b,c,d,e,f,g,h,j |
| Physicochemical Parameters | Indexes of Correlation | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| pH | - | - | - | - | - | - | - | - | - |
| H2O2 | - | - | |||||||
| Free acidity | - | - | - | ||||||
| Lactonic acidity | - | - | r = 0.952 p < 0.00001 | - | |||||
| TPC | - | - | - | - | - | ||||
| TFC | - | - | - | - | - | - | |||
| DPPH | - | r = 0.802 p = 0.00521 | - | - | - | r = 0.89 p = 0.00054 | - | ||
| Tyrosinase inhibition | - | - | - | - | r = 0.636, p = 0.04791 | r = 0.872, p = 0.03304 | r = 0.696, p = 0.0251 | - | |
| Kojic acid | - | - | - | - | - | - | r = 0696, p = 0.0251 | r = 0.975, p < 0.0001 | - |
| pH | H2O2 | Free acidity | Lactonic acidity | TPC | TFC | DPPH | Tyrosinase inhibition | Kojic acid | |
| Bacterial Species | Number of Samples in Well Diffusion Assay | p-Value | ||||
|---|---|---|---|---|---|---|
| Concentration of Honey (% v/v) | ||||||
| 75% | 50% | 25% | 12.5% | 6.25% | ||
| Enterobacter cloacae subsp. dissolvens | 2 | 8 | 16 | 18 | 5 | <0.001 |
| Pseudomonas aeruginosa | 7 | 6 | 23 | 9 | 9 | <0.001 |
| Klebsiella pneumoniae subsp. pneumoniae (1) * | 6 | 11 | 30 | 10 | - | <0.001 |
| Klebsiella pneumoniae subsp. pneumoniae (2) ** | 5 | 19 | 31 | 12 | 2 | <0.001 |
| Bacterial Species | Number of Samples in the MIC95 Values (% v/v) Assessment | p-Value | ||
|---|---|---|---|---|
| Crude Samples | Catalase Addition | Protease Addition | ||
| Enterobacter cloacae subsp. dissolvens | 8 | 41 | 19 | <0.0001 |
| Pseudomonas aeruginosa | 5 | 2 | 1 | p = 0.178575 |
| Klebsiella pneumoniae subsp. pneumoniae (1) * | 1 | 31 | 18 | <0.0001 |
| Klebsiella pneumoniae subsp. pneumoniae (2) ** | 6 | 29 | 17 | <0.0001 |
| Honey Number | Botanical Source | Geographical Location | Honey Number | Botanical Source | Geographical Location |
|---|---|---|---|---|---|
| 1 | Cotton | Karditsa | 23 | Orange | Evros |
| 2 | Cotton | Evros | 24 | Oregano | Epirus |
| 3 | Cotton | Epirus | 25 | Oregano | Epirus |
| 4 | Cotton | Epirus | 26 | Oregano | Epirus |
| 5 | Arbutus | Arkadia | 27 | Oregano | Epirus |
| 6 | Arbutus | Epirus | 28 | Fir | Ftiotida |
| 7 | Arbutus | Epirus | 29 | Fir | Epirus |
| 8 | Arbutus | Evros | 30 | Fir | Epirus |
| 9 | Arbutus | Evros | 31 | Fir | Epirus |
| 10 | Chestnut | Epirus | 32 | Fir | Epirus |
| 11 | Chestnut | Epirus | 33 | Fir | Epirus |
| 12 | Chestnut | Epirus | 34 | Fir | Epirus |
| 13 | Chestnut | Evros | 35 | Sunflower | Evros |
| 14 | Chestnut | Evros | 36 | Sunflower | Evros |
| 15 | Thyme | Epirus | 37 | Cotton & Sunflower | Evros |
| 16 | Thyme | Attica (Laurion) | 38 | Heath | Epirus |
| 17 | Thyme | Epirus | 39 | Heath | Epirus |
| 18 | Thyme | Epirus | 40 | Fir & Heath | Arkadia |
| 19 | Thyme | Epirus | 41 | Heath | Epirus |
| 20 | Orange | Epirus | 42 | Heath | Epirus |
| 21 | Orange | Epirus | 43 | AM HEALTH Manuka Health MGO™550+ (25+) | Lower Hutt, New Zealand |
| 22 | Orange | Epirus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stavropoulou, E.; Voidarou, C.; Rozos, G.; Vaou, N.; Bardanis, M.; Konstantinidis, T.; Vrioni, G.; Tsakris, A. Antimicrobial Evaluation of Various Honey Types against Carbapenemase-Producing Gram-Negative Clinical Isolates. Antibiotics 2022, 11, 422. https://doi.org/10.3390/antibiotics11030422
Stavropoulou E, Voidarou C, Rozos G, Vaou N, Bardanis M, Konstantinidis T, Vrioni G, Tsakris A. Antimicrobial Evaluation of Various Honey Types against Carbapenemase-Producing Gram-Negative Clinical Isolates. Antibiotics. 2022; 11(3):422. https://doi.org/10.3390/antibiotics11030422
Chicago/Turabian StyleStavropoulou, Elisavet, Chrysoula (Chrysa) Voidarou, Georgios Rozos, Natalia Vaou, Michael Bardanis, Theodoros Konstantinidis, Georgia Vrioni, and Athanasios Tsakris. 2022. "Antimicrobial Evaluation of Various Honey Types against Carbapenemase-Producing Gram-Negative Clinical Isolates" Antibiotics 11, no. 3: 422. https://doi.org/10.3390/antibiotics11030422