
Pathogenic E. coli from Cattle as a Reservoir of Resistance Genes to Various Groups of Antibiotics

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Prevalence of Pathogenic E. coli Strains
2.2. Resistance Profiles of Pathogenic E. coli Strains
2.3. Resistance Genes Presence in Pathogenic E. coli Strains
3. Discussion
4. Materials and Methods
4.1. Samples and Isolates
4.2. Bacterial Genomic DNA Extraction
4.3. Virulence Gene Identification
4.4. Susceptibility Testing
4.5. PCR Method for Identification of Resistance Genes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO (World Health Organization). Antimicrobial Resistance. 17 November 2021. Available online: https://www.who.int/ (accessed on 2 November 2021).
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol Spectr. 2016, 4, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Raymond, B. Five rules for resistance management in the antibiotic apocalypse, a road map for integrated microbial management. Evol. Appl. 2019, 12, 1079–1091. [Google Scholar] [CrossRef]
- Bae, W.K.; Lee, Y.K.; Cho, M.S.; Ma, S.K.; Kim, S.W.; Kim, N.H.; Choi, K.C. A case of hemolytic uremic syndrome caused by Escherichia coli O104:H4. Yonsei Med. J. 2006, 47, 437–439. [Google Scholar] [CrossRef] [PubMed]
- Keir, L.S.; Marks, S.D.; Kim, J.J. Shigatoxin-associated hemolytic uremic syndrome: Current molecular mechanisms and future therapies. Drug Des. Devel. Ther. 2012, 6, 195–208. [Google Scholar]
- Fagan, P.K.; Hornitzky, M.A.; Bettelheim, K.A.; Djordjevic, S.P. Detection of shiga-like toxin (stx1 and stx2), intimin (eaeA), and enterohemorrhagic Escherichia coli (EHEC) hemolysin (EHEC hlyA) genes in animal feces by multiplex PCR. Appl. Environ. Microbiol. 1999, 65, 868–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paton, J.C.; Paton, A.W. Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli infections. Clin. Microbiol. Rev. 1998, 11, 450–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, A.R.; Sperandio, V. Shiga toxin in enterohemorrhagic E.coli: Regulation and novel anti-virulence strategies. Front. Cell. Infect. Microbiol. 2012, 2, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Rodríguez, N.E.; Barba-León, J.; Navarro-Ocaña, A.; Vega-Sánchez, V.; Gómez De Anda, F.R.; Talavera-González, J.M.; Talavera-Rojas, M. Serotypes and Stx2 subtyping of Shiga toxin producing Escherichia coli isolates from cattle carcasses and feces. Rev. Mex. Cienc. Pecu. 2020, 11, 1030–1044. [Google Scholar]
- Oporto, B.; Ocejo, M.; Alkorta, M.; Marimón, J.M.; Montes, M.; Hurtado, A. Zoonotic approach to Shiga toxin-producing Escherichia coli: Integrated analysis of virulence and antimicrobial resistance in ruminants and humans. Epidemiol. Infect. 2019, 147, e164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianca, A.; Amézquita-López, M.S.B.; Bertram, G.L.; Jaszemyn, C.Y. Isolation, genotyping and antimicrobial resistance of Shiga toxin-producing Escherichia coli. J. Microbiol. Immunol. Infect. 2018, 51, 425–434. [Google Scholar]
- Okeke, I.N.; Aboderin, O.A.; Byarugaba, D.K.; Ojo, K.K.; Opintan, J.A. Growing problem of multidrug-resistant enteric pathogens in Africa. Emerg. Infect. Dis. 2007, 13, 1640–1646. [Google Scholar] [CrossRef]
- Imre, K.; Herman, V.; Morar, A. Scientific Achievements in the Study of the Occurrence and Antimicrobial Susceptibility Profile of Major Foodborne Pathogenic Bacteria in Foods and Food Processing Environments in Romania: Review of the Last Decade. BioMed Res. Int. 2020, 5, 5134764. [Google Scholar] [CrossRef]
- Armstrong, G.L.; Hollingsworth, J.; Glenn Morris, J. Emerging Foodborne Pathogens: Escherichia coli O157:H7 as a Model of Entry of a New Pathogen into the Food Supply of the Developed World. Epidemiol. Rev. 1996, 18, 29–51. [Google Scholar] [CrossRef]
- Duffell, E.; Espié, E.; Nichols, T.; Adak, G.K.; De Valk, H.; Anderson, K.; Stuart, J.M. Investigation of an outbreak of E. coli O157 infections associated with a trip to France of schoolchildren from Somerset, England. Eurosurveillance 2003, 8, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Gill, A.; Gill, C.O. Non-O157 verotoxigenic Escherichia coli and beef: A Canadian perspective. Can. J. Vet. Res. 2010, 74, 161–169. [Google Scholar] [PubMed]
- Wieler, L.H.; Vieler, E.; Erpenstein, C.; Schlapp, T.; Steinrück, H.; Bauerfeind, R.; Byomi, A.; Baljer, G. Shiga toxin-producing Escherichia coli strains from bovines: Association of adhesion with carriage of eae and other genes. J. Clin. Microbiol. 1996, 34, 2980–2984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eklund, M.; Leino, K.; Siitonen, A. Clinical Escherichia coli strains carrying stx genes: Stx variants and stx-positive virulence profiles. J. Clin. Microbiol. 2002, 40, 4585–4593. [Google Scholar] [CrossRef] [Green Version]
- Pradel, N.; Livrelli, V.; De Champs, C.; Palcoux, J.B.; Reynaud, A.; Scheutz, F.; Sirot, J.; Joly, B.; Forestier, C. Prevalence and characterization of Shiga toxin-producing Escherichia coli isolated from cattle, food, and children during a one-year prospective study in France. J. Clin. Microbiol. 2000, 38, 1023–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirila, F.; Tabaran, A.; Fit, N.; Nadas, G.; Mihaiu, M.; Tabaran, A.F.; Cătoi, C.; Reget, O.L.; Dan, S.D. Concerning increase in antimicrobial resistance in Shiga toxin-producing Escherichia coli isolated from young animals during 1980–2016. Microbes Environ. 2017, 32, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vázquez, A.V.; Vázquez-Villanueva, J.; Leyva-Zapata, L.M.; Barrios-García, H.; Rivera, G.; Bocanegra-García, V. Multidrug Resistance of Escherichia coli Strains Isolated from Bovine Feces and Carcasses in Northeast Mexico. Front. Vet. Sci. 2021, 8, 643802. [Google Scholar] [CrossRef]
- Economou, V.; Gousia, P. Agriculture and Food Animals as a Source of Antimicrobial-Resistant Bacteria. Infect. Drug Resist. 2015, 1, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Mesa Varona, O.; Chaintarli, K.; Muller-Pebody, B.; Anjum, M.F.; Eckmanns, T.; Norström, M.; Boone, I.; Tenhagen, B.A. Monitoring Antimicrobial Resistance and Drug Usage in the Human and Livestock Sector and Foodborne Antimicrobial Resistance in Six European Countries. Infect. Drug Resist. 2020, 13, 957–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramadan, H.; Jackson, C.R.; Frye, J.G.; Hiott, L.M.; Samir, M.; Awad, A. Antimicrobial resistance, genetic diversity and multilocus sequence typing of Escherichia coli from humans, retail chicken and ground beef in Egypt. Pathogens 2020, 9, 357. [Google Scholar] [CrossRef] [PubMed]
- Bumunangg, E.W.; McAllister, T.A.; Zaheer, R.; Ortega, R.; Stanford, K.; King, R. Characterization of non-O157 Escherichia coli from cattle faecal samples in the north-west province of South Africa. Microorganisms 2019, 7, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obaidat, M.M.; Bani, A.E.; Davis, M.A.; Roess, A.A. Major diseases, extensive misuse, and high antimicrobial resistance of Escherichia coli in large-and small- scale dairy cattle farms in Jordan. J. Dairy Sci. 2018, 101, 2324–2334. [Google Scholar] [CrossRef] [Green Version]
- Cavalcanti, A.M.F.; Hernandes, R.T.; Takagi, E.H. Virulence Profiling and Molecular Typing of Shiga Toxin-Producing E. coli (STEC) from Human Sources in Brazil. Microorganisms 2020, 8, 171. [Google Scholar] [CrossRef] [Green Version]
- Tahamtan, Y.; Hayati, M.; Namavari, M. Prevalence and distribution of the stx, stx genes in Shiga toxin producing E. coli (STEC) isolates from cattle. Iran. J. Microbiol. 2010, 2, 8–13. [Google Scholar] [PubMed]
- SO 16654/2001. Microbiology-Horizontal Method for the Detection of Escherichia coli O157, 1st ed.; International Organization for Standardization: Geneve, Switzerland, 2001. [Google Scholar]
- Mihaiu, L.; Lăpuşan, A.; Tănăsuică, R.; Sobolu, R.; Mihaiu, R.; Oniga, O.; Mihaiu, M. First study of Salmonella in meat in Romania. J. Infect. Dev. Ctries. 2014, 8, 50–58. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; Clinical and Laboratory Standards Institute: Wayne, NY, USA, 2022. [Google Scholar]


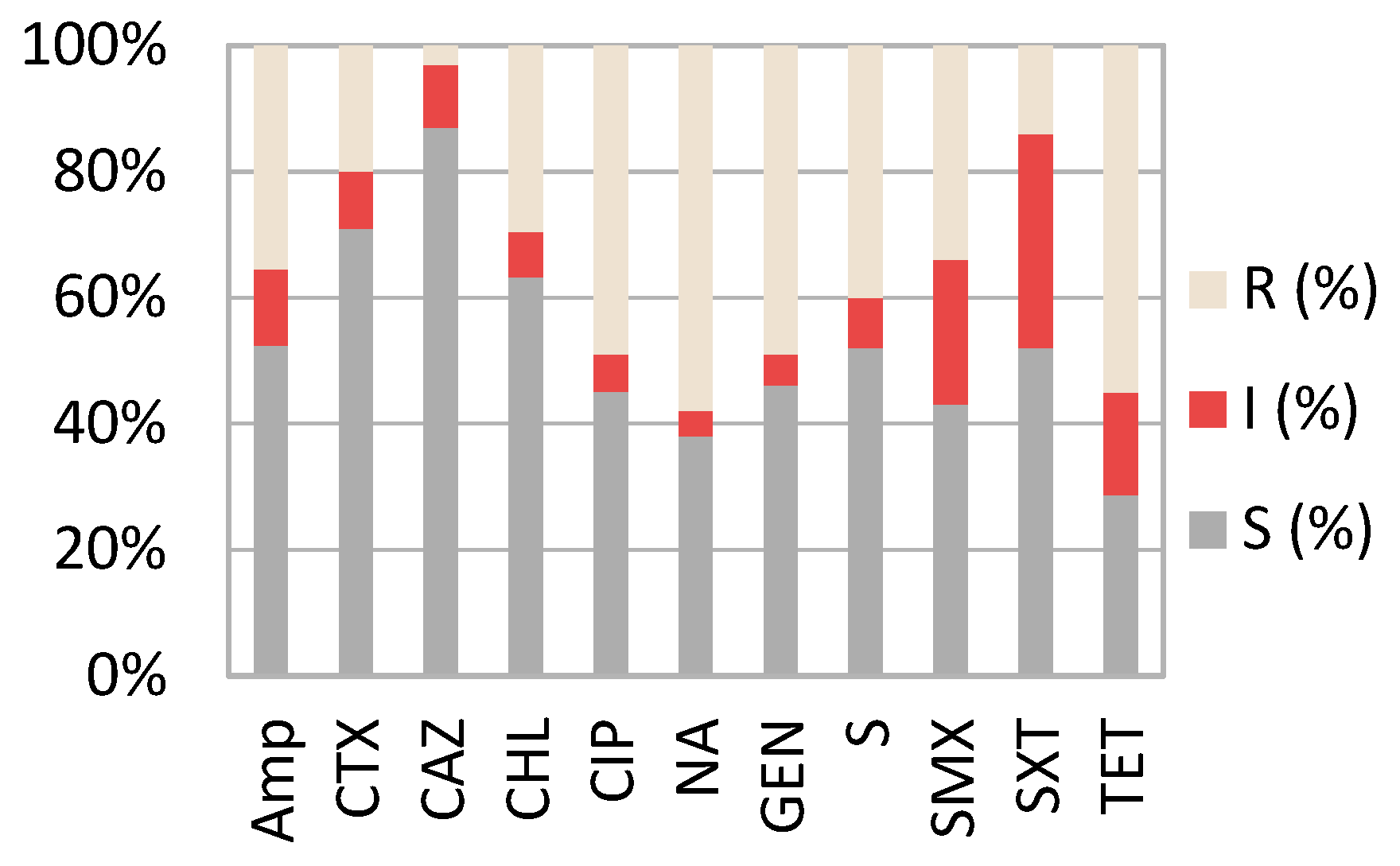{kind=link}
| Country | No of Samples Tested | Shiga Toxin Genes | Total (%) | ||
|---|---|---|---|---|---|
| Stx 1 | Stx 2 | eaeA | |||
| Romania | 164 | + | − | − | 23 (14) |
| + | − | + | 2 (1) | ||
| − | + | + | 0 | ||
| − | − | + | 1 (1) | ||
| + | + | − | 3 (2) | ||
| Total no of samples positive to at least one virulence gene | 29 (17) | ||||
| France | 78 | + | − | − | 8 (10) |
| + | − | + | 2 (3) | ||
| − | + | + | 1 (1) | ||
| − | − | + | 0 | ||
| + | + | − | 2 (3) | ||
| Total no of samples positive to at least one virulence gene | 13 (16) | ||||
| E. coli Strain/Country | Phenotype of Resistance | Resistance Genes | Toxinogenic Genes/Sample No. |
|---|---|---|---|
| 1–2/Romania | AMP, TET, SMX, SXT, CTX, NA | blaTEM, tetA, sul1, dfrA1,ere(A) | stx1 (1,2) |
| 3–5/Romania 6/France | TET, SMX, SXT, CIP, NA, AMP | blaTEM, tetA, tetB, sul1, dfrA1,ere(A) | stx1 (4,6) stx1, eaeA (3) stx1, Stx2 (5) |
| 7/Romania 8/France | AMP, TET, SMX, SXT, NA, CHL | blaTEM, tetA, tetB, sul1, dfrA1 | stx1 (7,8) |
| 9/Romania | TET, SMX, SXT, NA, CAZ, S | blaTEM, blaSHV, blaCMY, tetA, tetB, aadA1, dfrA1 | stx1, eaeA |
| 10,11/Romania 12/France | TET, SMX, SXT, CTX, S | tet A, tetB, sul1,ere(A) | stx1 (10,11) stx1, eaeA (12) |
| 13/Romania 14/France | AMP, CIP, TET, GEN | tetA, tetB,ere(A) | stx1, Stx2 (13) stx2, eaeA (14) |
| 15–17/Romania 18/France | TET, NA, AMP, GEN | tetA,ere(A) | stx1, stx2, (16,17,18) stx 1 (15) |
| 19/Romania | TET, NA, GEN, CTX, CIP | tetA, tetB | stx1, stx2 |
| 20/Romania | TET, NA, S, | tetA, aadA1 | stx1 |
| 21/Romania | TET, NA, CAZ, SMX, AMP | tetA, sul1,ere(A) | stx1 |
| 22/Romania | NA, SMX, SXT, CTX | sul1 | stx1 |
| 23/Romania | NA, SMX, SXT, CIP, AMP | tetA, sul1,ere(A) | stx1 |
| 24–25/Romania | TET, NA, GEN, CIP, S, AMP | tetA, sul1, aadA1,ere(A) | stx1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabaran, A.; Soulageon, V.; Chirila, F.; Reget, O.L.; Mihaiu, M.; Borzan, M.; Dan, S.D. Pathogenic E. coli from Cattle as a Reservoir of Resistance Genes to Various Groups of Antibiotics. Antibiotics 2022, 11, 404. https://doi.org/10.3390/antibiotics11030404
Tabaran A, Soulageon V, Chirila F, Reget OL, Mihaiu M, Borzan M, Dan SD. Pathogenic E. coli from Cattle as a Reservoir of Resistance Genes to Various Groups of Antibiotics. Antibiotics. 2022; 11(3):404. https://doi.org/10.3390/antibiotics11030404
Chicago/Turabian StyleTabaran, Alexandra, Virginie Soulageon, Flore Chirila, Oana Lucia Reget, Marian Mihaiu, Mihai Borzan, and Sorin Daniel Dan. 2022. "Pathogenic E. coli from Cattle as a Reservoir of Resistance Genes to Various Groups of Antibiotics" Antibiotics 11, no. 3: 404. https://doi.org/10.3390/antibiotics11030404