
Combined Effect of Phage phT4A and Pressure-Based Strategies in the Inhibition of Escherichia coli

 , , and
, , and 
Abstract
:1. Introduction
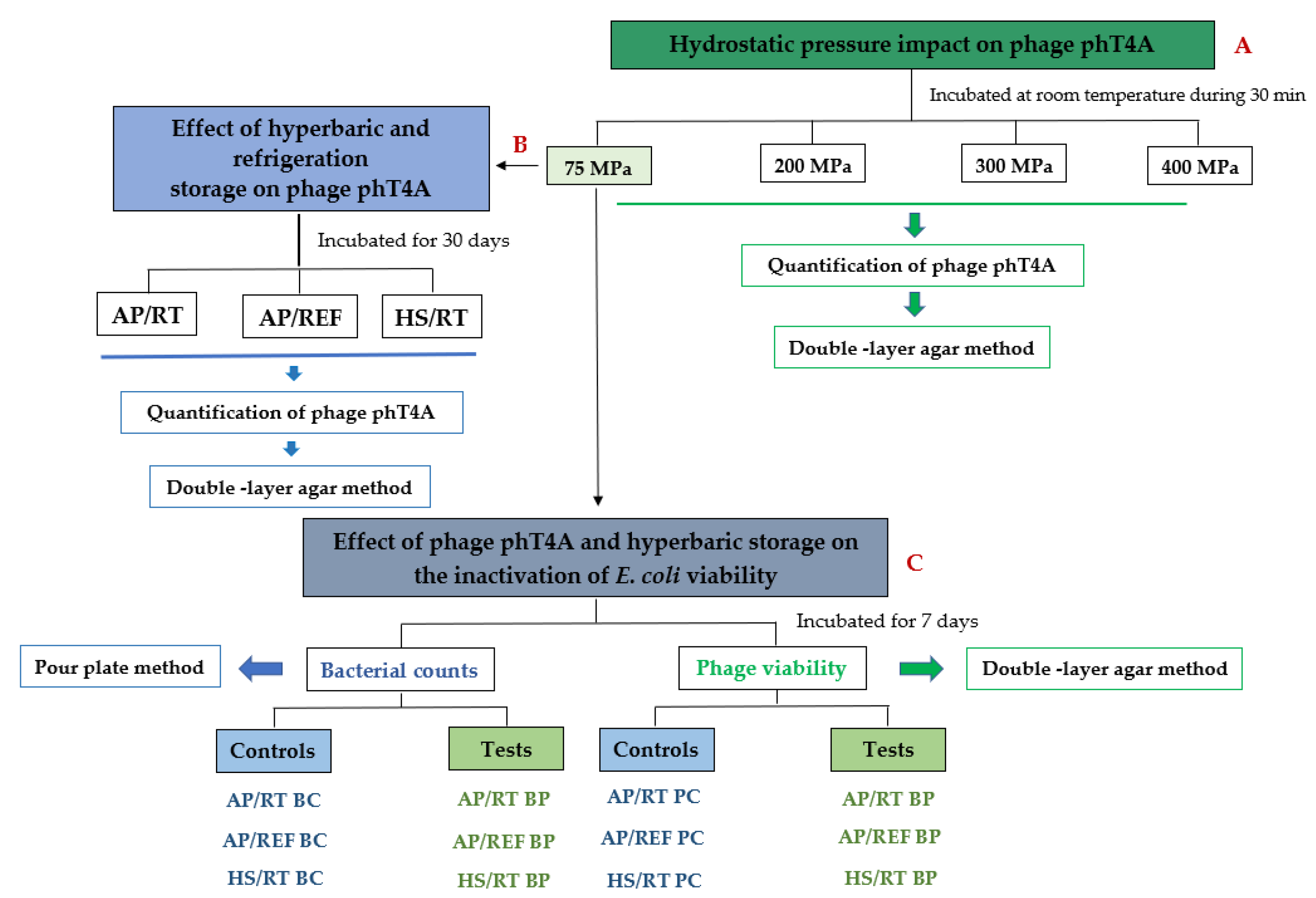
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Preparation of Phage phT4A and Enrichment
2.3. Hydrostatic Pressure Impact on Phage phT4A Viability
2.4. Effect of Hyperbaric and Refrigeration Storage on Phage phT4A Viability
2.5. Hyperbaric Storage Experiments
2.6. Statistical Analysis
3. Results
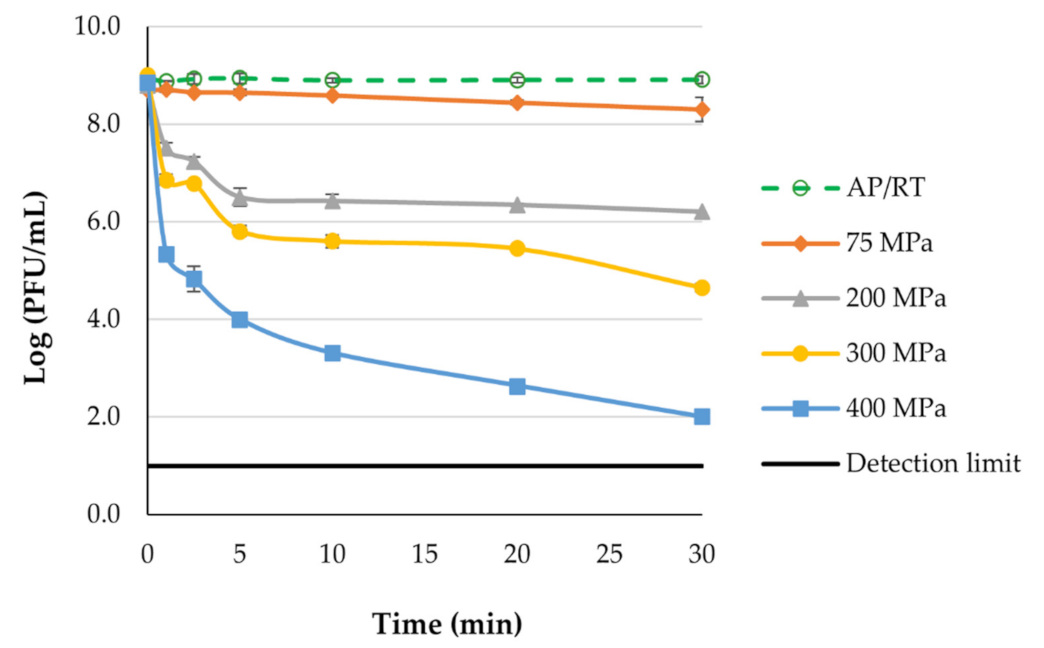
3.1. Hydrostatic Pressure Impact on Phage phT4A Viability
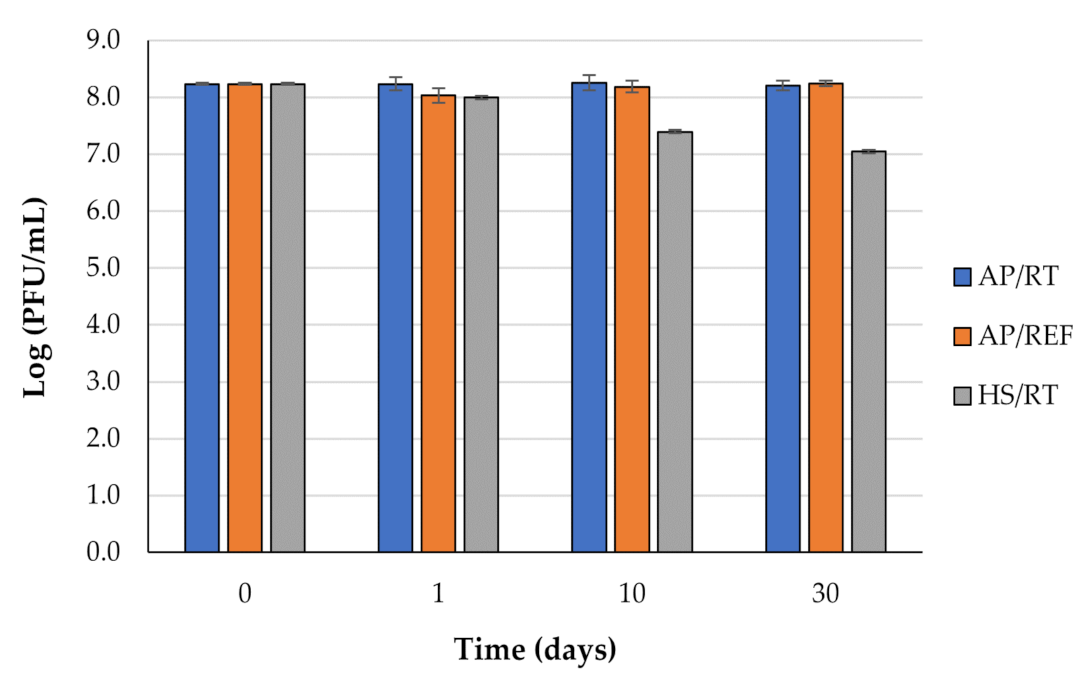
3.2. Effect of Hyperbaric and Refrigeration Storage on Phage phT4A Viability
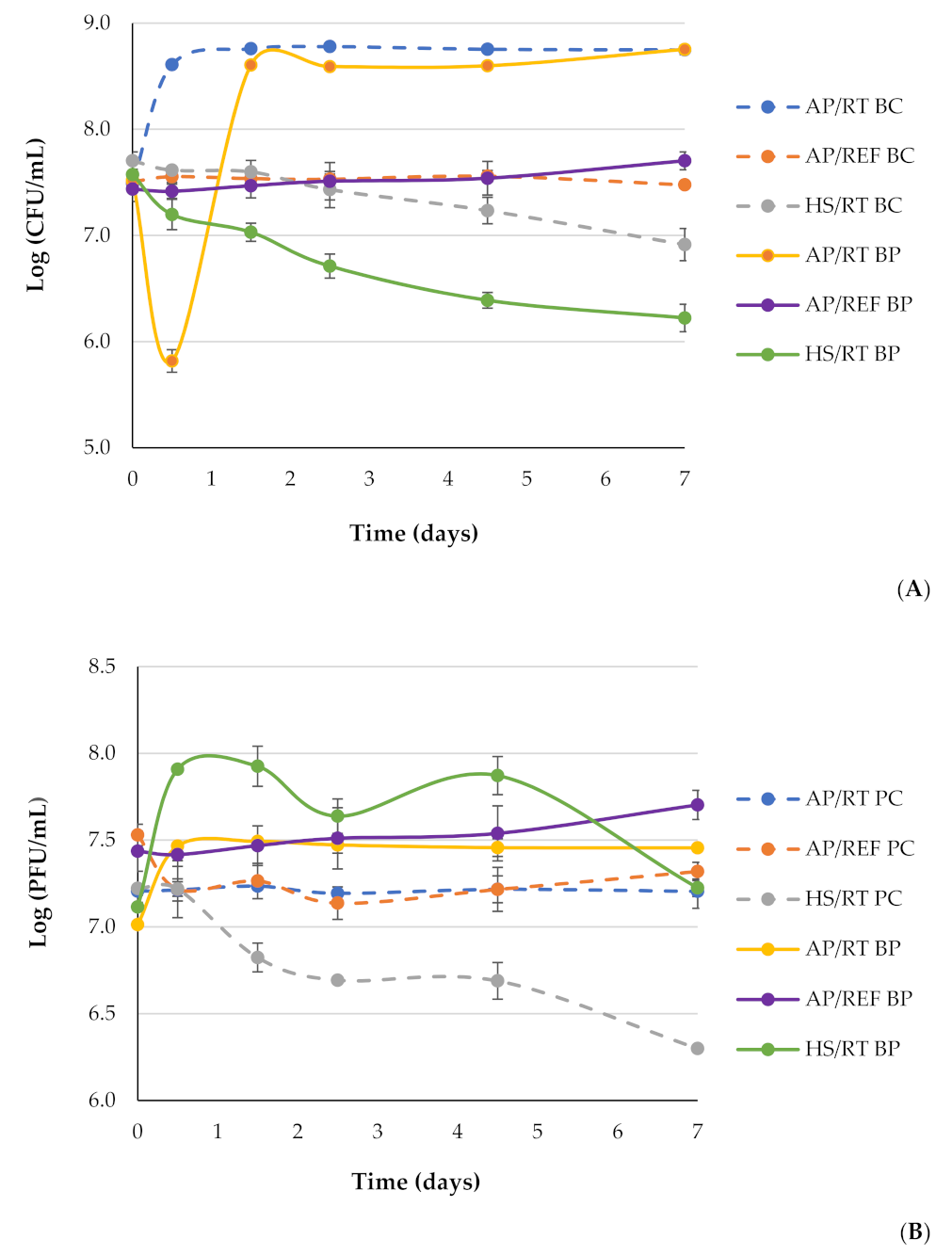
3.3. Effect of Phage phT4A and Hyperbaric Storage on the Inactivation of E. coli
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529–563. [Google Scholar] [CrossRef] [PubMed]
- Makinde, O.; Ayeni, K.; Sulyok, M.; Krska, R.; Adeleke, R.; Ezekiel, C. Microbiological safety of ready-to-eat foods in low- and middle-income countries: A comprehensive 10-year (2009 to 2018) review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 703–732. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, T.Y.; Ye, C.; Chen, L.; Liang, Y.; Wang, K.; Liu, J. Formation and control of the viable but non-culturable state of foodborne pathogen Escherichia coli O157:H7. Front. Microbiol. 2020, 11, 1202. [Google Scholar] [CrossRef]
- Rodrigues, C.; da Silva, A.L.B.R.; Dunn, L.L. Factors impacting the prevalence of foodborne pathogens in agricultural water sources in the southeastern United States. Water 2020, 12, 51. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.; Pereira, C.; Almeida, A. Sequential combined effect of phages and antibiotics on the inactivation of E. coli. Microorganisms 2018, 6, 125. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Attributing Illness Caused by Shiga Toxin-Producing Escherichia Coli (STEC) to Specific Foods; World Health Organization: Rome, Italy, 2019. [Google Scholar]
- Puño-Sarmiento, J.; Anderson, E.M.; Park, A.J.; Khursigara, C.M.; Barnett Foster, D.E. Potentiation of antibiotics by a novel antimicrobial peptide against Shiga toxin producing E. coli O157:H7. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage applications for food production and processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention (CDC). Estimates of Foodborne Illness in the United States. Available online: https://www.cdc.gov/foodborneburden/2011-foodborne-estimates.html (accessed on 3 December 2021).
- Pereira, C.; Costa, P.; Duarte, J.; Balcão, V.M.; Almeida, A. Phage therapy as a potential approach in the biocontrol of pathogenic bacteria associated with shellfish consumption. Int. J. Food Microbiol. 2021, 338, 108995. [Google Scholar] [CrossRef] [PubMed]
- Endersen, L.; O’Mahony, J.; Hill, C.; Ross, R.P.; McAuliffe, O.; Coffey, A. Phage therapy in the food industry. Annu. Rev. Food Sci. Technol. 2014, 5, 327–349. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Costa, P.; Pinheiro, L.; Balcão, V.M.; Almeida, A. Kiwifruit bacterial canker: An integrative view focused on biocontrol strategies. Planta 2021, 253, 49. [Google Scholar] [CrossRef]
- O’Flynn, G.; Ross, R.P.; Fitzgerald, G.F.; Coffey, A. Evaluation of a cocktail of three bacteriophages for biocontrol of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2004, 70, 3417–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, J.; Billington, C.; Wilson, T.; On, S. Effect of phage and host concentration on the inactivation of Escherichia coli O157:H7 on cooked and raw beef. Food Sci. Technol. Int. 2013, 21, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Abuladze, T.; Li, M.; Menetrez, M.Y.; Dean, T.; Senecal, A.; Sulakvelidze, A. Bacteriophages reduce experimental contamination of hard surfaces, tomato, spinach, broccoli, and ground beef by Escherichia coli O157:H7. Appl. Environ. Microbiol. 2008, 74, 6230–6238. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Patel, J.R.; Conway, W.S.; Ferguson, S.; Sulakvelidze, A. Effectiveness of bacteriophages in reducing Escherichia coli O157:H7 on fresh-cut cantaloupes and lettuce. J. Food Prot. 2009, 72, 1481–1485. [Google Scholar] [CrossRef] [Green Version]
- Carter, C.D.; Parks, A.; Abuladze, T.; Li, M.; Woolston, J.; Magnone, J.; Senecal, A.; Kropinski, A.M.; Sulakvelidze, A. Bacteriophage cocktail significantly reduces Escherichia coli O157: H7 contamination of lettuce and beef, but does not protect against recontamination. Bacteriophage 2012, 2, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.; Roberts, C.; Handy, E.; Sharma, M. Lytic bacteriophages reduce Escherichia coli O157. Bacteriophage 2013, 3, e24323. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Moreirinha, C.; Teles, L.; Rocha, R.J.M.; Calado, R.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Application of phage therapy during bivalve depuration improves Escherichia coli decontamination. Food Microbiol. 2017, 61, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Moreirinha, C.; Rocha, R.J.M.; Calado, R.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Application of bacteriophages during depuration reduces the load of Salmonella Typhimurium in cockles. Food Res. Int. 2016, 90, 73–84. [Google Scholar] [CrossRef]
- McLean, S.; Dunn, L.; Palombo, E. Phage inhibition of Escherichia coli in ultrahigh-temperature-treated and raw milk. Foodborne Pathog. Dis. 2013, 10, 956–962. [Google Scholar] [CrossRef]
- Tomat, D.; Mercanti, D.; Balagué, C.; Quiberoni, A. Phage biocontrol of enteropathogenic and Shiga toxin-producing Escherichia coli during milk fermentation. Lett. Appl. Microbiol. 2013, 57, 3–10. [Google Scholar] [CrossRef]
- Bach, S.; Mcallister, T.; Veira, D.; Gannon, V.; Holley, R. Effect of bacteriophage DC22 on Escherichia coli O157: H7 in an artificial rumen system (Rusitec) and inoculated sheep. Anim. Res. 2003, 52, 89–101. [Google Scholar] [CrossRef]
- Gill, J.J.; Hyman, P. Phage choice, isolation, and preparation for phage therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef]
- Deveau, H.; Garneau, J.E.; Moineau, S. CRISPR/Cas system and its role in phage-bacteria interactions. Annu. Rev. Microbiol. 2010, 64, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Vegge, C.S.; Schmerer, M.; Chaudhry, W.N.; Levin, B.R. Phenotypic resistance and the dynamics of bacterial escape from phage control. PLoS ONE 2014, 9, e94690. [Google Scholar] [CrossRef] [PubMed]
- Lou, F.; Neetoo, H.; Chen, H.; Li, J. High hydrostatic pressure processing: A promising nonthermal technology to inactivate viruses in high-risk foods. Annu. Rev. Food Sci. Technol. 2015, 6, 389–409. [Google Scholar] [CrossRef]
- Picart-Palmade, L.; Cunault, C.; Chevalier-Lucia, D.; Belleville, M.P.; Marchesseau, S. Potentialities and limits of some non-thermal technologies to improve sustainability of food processing. Front. Nutr. 2019, 5, 130. [Google Scholar] [CrossRef] [PubMed]
- Otero, L. Hyperbaric storage at room temperature for fruit juice preservation. Beverages 2019, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Pinto, C.; Moreira, S.A.; Fidalgo, L.G.; Santos, M.D.; Vidal, M.; Delgadillo, I.; Saraiva, J.A. Impact of different hyperbaric storage conditions on microbial, physicochemical and enzymatic parameters of watermelon juice. Food Res. Int. 2017, 99, 123–132. [Google Scholar] [CrossRef]
- Moreira, S.A.; Duarte, R.V.; Fernandes, P.A.R.; Alves, S.P.; Bessa, R.J.; Delgadillo, I.; Saraiva, J.A. Hyperbaric storage preservation at room temperature using an industrial-scale equipment: Case of two commercial ready-to-eat pre-cooked foods. Innov. Food Sci. Emerg. Technol. 2015, 32, 29–36. [Google Scholar] [CrossRef]
- Pinto, C.; Moreira, S.A.; Fidalgo, L.G.; Santos, M.D.; Delgadillo, I.; Saraiva, J.A. Shelf-life extension of watermelon juice preserved by hyperbaric storage at room temperature compared to refrigeration. LWT Food Sci. Technol. 2016, 72, 78–80. [Google Scholar] [CrossRef]
- Segovia-Bravo, K.A.; Guignon, B.; Bermejo-Prada, A.; Sanz, P.D.; Otero, L. Hyperbaric storage at room temperature for food preservation: A study in strawberry juice. Innov. Food Sci. Emerg. Technol. 2012, 15, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Freitas, P.; Pereira, S.A.; Santos, M.D.; Alves, S.P.; Bessa, R.J.B.; Delgadillo, I.; Saraiva, J.A. Performance of raw bovine meat preservation by hyperbaric storage (quasi energetically costless) compared to refrigeration. Meat Sci. 2016, 121, 64–72. [Google Scholar] [CrossRef]
- Moreira, S.A.; Fernandes, P.A.R.; Duarte, R.; Santos, D.I.; Fidalgo, L.G.; Santos, M.D.; Queirós, R.P.; Delgadillo, I.; Saraiva, J.A. A first study comparing preservation of a ready-to-eat soup under pressure (hyperbaric storage) at 25 °C and 30 °C with refrigeration. Food Sci. Nutr. 2015, 3, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Prada, A.; López-Caballero, M.E.; Otero, L. Hyperbaric storage at room temperature: Effect of pressure level and storage time on the natural microbiota of strawberry juice. Innov. Food Sci. Emerg. Technol. 2016, 33, 154–161. [Google Scholar] [CrossRef]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeidab, P.; Clemente, C.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Characterization and in vitro evaluation of new bacteriophages for the biocontrol of Escherichia coli. Virus Res. 2016, 227, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.H. Bacteriophages; Interscience Publishers: New York, NY, USA, 1959. [Google Scholar]
- Duarte, J.; Pereira, C.; Costa, P.; Almeida, A. Bacteriophages with potential to inactivate Aeromonas hydrophila in cockles: In vitro and in vivo preliminary studies. Antibiotics 2021, 10, 710. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeida, P.; Clemente, C.; Cunha, Â.; Delgadillo, I.; Romalde, J.; Nunes, M.L.; Almeida, A. Bacteriophages with potential to inactivate Salmonella Typhimurium: Use of single phage suspensions and phage cocktails. Virus Res. 2016, 220, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Valério, N.; Oliveira, C.; Jesus, V.; Branco, T.; Pereira, C.; Moreirinha, C.; Almeida, A. Effects of single and combined use of bacteriophages and antibiotics to inactivate Escherichia coli. Virus Res. 2017, 240, 8–17. [Google Scholar] [CrossRef]
- Komora, N.; Maciel, C.; Pinto, C.A.; Ferreira, V.; Brandão, T.R.S.; Saraiva, J.M.A.; Castro, S.M.; Teixeira, P. Non-thermal approach to Listeria monocytogenes inactivation in milk: The combined effect of high pressure, pediocin PA-1 and bacteriophage P100. Food Microbiol. 2020, 86, 103315. [Google Scholar] [CrossRef]
- Khadre, M.A.; Yousef, A.E. Susceptibility of human rotavirus to ozone, high pressure and pulsed electric field. J. Food Prot. 2002, 65, 1441–1446. [Google Scholar] [CrossRef]
- Tang, Q.; Li, D.; Xu, J.; Wang, J.; Zhao, Y.; Li, Z.; Xue, C. Mechanism of inactivation of murine norovirus-1 by high pressure processing. Int. J. Food Microbiol. 2010, 137, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tang, Q.; Wang, J.; Wang, Y.; Zhao, Q.; Xue, C. Effects of high-pressure processing on murine norovirus-1 in oysters (Crassostrea gigas) in situ. Food Control 2009, 20, 992–996. [Google Scholar] [CrossRef]
- Kingsley, D.H.; Chen, H. Influence of pH, salt, and temperature on pressure inactivation of hepatitis A virus. Int. J. Food Microbiol. 2009, 130, 61–64. [Google Scholar] [CrossRef]
- Lou, F.; Neetoo, H.; Chen, H.; Li, J. Inactivation of a human norovirus surrogate by high-pressure processing: Effectiveness, mechanism, and potential application in the fresh produce industry. Appl. Environ. Microbiol. 2011, 77, 1862–1871. [Google Scholar] [CrossRef] [Green Version]
- Kingsley, D.H.; Chen, H.Q. Aqueous matrix compositions and pH influence feline calicivirus inactivation by high pressure processing. J. Food Prot. 2008, 71, 1598–1603. [Google Scholar] [CrossRef]
- Kingsley, D.H.; Holliman, D.R.; Calci, K.R.; Chen, H.; Flick, G.J.; Al, K.E.T. Inactivation of a norovirus by high-pressure processing. Appl. Environ. Microbiol. 2007, 73, 581–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, P.; Pereira, C.; Gomes, A.; Almeida, A. Efficiency of single phage suspensions and phage cocktail in the inactivation of Escherichia coli and Salmonella Typhimurium: An in vitro preliminary study. Microorganisms 2019, 7, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, J.; Sharma, M.; Millner, P.; Calaway, T.; Singh, M. Inactivation of Escherichia coli O157:H7 attached to spinach harvester blade using bacteriophage. Foodborne Pathog. Dis. 2011, 8, 541–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Fernandes, P.; Moreira, S.; Duarte, R.; Santos, D.; Queirós, R.; Fidalgo, L.; Santos, M.; Delgadillo, I.; Saraiva, J. Preservation of sliced cooked ham at 25, 30 and 37 °C under moderated pressure (hyperbaric storage) and comparison with refrigerated storage. Food Bioprod. Process 2015, 95, 200–207. [Google Scholar] [CrossRef]
- Fernandes, P.; Moreira, S.; Santos, M.; Duarte, R.; Santos, D.; Inacio, R.; Alves, S.; Bessa, R.; Delgadillo, I.; Saraiva, J. Hyperbaric storage at variable room temperature-a new preservation methodology for minced meat compared to refrigeration. J. Sci. Food Agric. 2019, 99, 3276–3282. [Google Scholar] [CrossRef] [PubMed]
- Fidalgo, L.G.; Simões, M.M.Q.; Casal, S.; Lopes-da-Silva, J.A.; Delgadillo, I.; Saraiva, J.A. Enhanced preservation of vacuum-packaged Atlantic salmon by hyperbaric storage at room temperature versus refrigeration. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Duarte, R.V.; Moreira, S.A.; Fernandes, P.A.R.; Fidalgo, L.G.; Santos, M.D.; Queirós, R.P.; Santos, D.I.; Delgadillo, I.; Saraiva, J.A. Preservation under pressure (hyperbaric storage) at 25 °C, 30 °C and 37 °C of a highly perishable dairy food and comparison with refrigeration. CYTA J. Food 2015, 13, 321–328. [Google Scholar] [CrossRef]
- Duarte, R.V.; Moreira, S.A.; Fernandes, P.A.R.; Santos, D.I.; Inácio, R.S.; Alves, S.P.; Bessa, R.J.B.; Saraiva, J.A. Whey cheese longer shelf-life achievement at variable uncontrolled room temperature and comparison to refrigeration. J. Food Process. Preserv. 2017, 41, e13307. [Google Scholar] [CrossRef]
- Chibeu, A.; Agius, L.; Gao, A.; Sabour, P.M.; Kropinski, A.M.; Balamurugan, S. Efficacy of bacteriophage LISTEXTMP100 combined with chemical antimicrobials in reducing Listeria monocytogenes in cooked turkey and roast beef. Int. J. Food Microbiol. 2013, 167, 208–214. [Google Scholar] [CrossRef]
- Mojica, K.D.A.; Brussaard, C.P.D. Factors affecting virus dynamics and microbial host-virus interactions in marine environments. FEMS Microbiol. Ecol. 2014, 89, 495–515. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.R.; Axler, R.P.; Hicks, R.E. Effects of freezing and storage temperature on MS2 viability. J. Virol. Methods 2004, 122, 147–152. [Google Scholar] [CrossRef]
- Pinheiro, L.A.M.; Pereira, C.; Frazão, C.; Balcão, V.M.; Almeida, A. Efficiency of phage φ6 for biocontrol of Pseudomonas syringae pv. syringae: An in vitro preliminary study. Microorganisms 2019, 7, 286–309. [Google Scholar] [CrossRef] [Green Version]
- Silva, Y.; Costa, L.; Pereira, C.; Cunha, A.; Calado, R.; Gomes, N.; Almeida, A. Influence of environmental variables in the efficiency of phage therapy in aquaculture. Microb. Biotechnol. 2014, 7, 401–413. [Google Scholar] [CrossRef]
- Duarte, J.; Pereira, C.; Moreirinha, C.; Salvio, R.; Lopes, A.; Wang, D.; Almeida, A. New insights on phage efficacy to control Aeromonas salmonicida in aquaculture systems: An in vitro preliminary study. Aquaculture 2018, 495, 970–982. [Google Scholar] [CrossRef]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The influence of external factors on bacteriophages—Review. Folia Microbiol. 2011, 56, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y. Phage and phage lysins: New era of bio-preservatives and food safety agents. J. Food Sci. 2021, 86, 3349–3373. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Zhang, P.; Zhang, H.; Zhou, Y.; Zhang, L.; Wang, R. Bio-control of Salmonella Enteritidis in foods using bacteriophages. Viruses 2015, 7, 4836–4853. [Google Scholar] [CrossRef] [PubMed]
- Zinno, P.; Devirgiliis, C.; Ercolini, D.; Ongeng, D.; Mauriello, G. Bacteriophage P22 to challenge Salmonella in foods. Int. J. Food Microbiol. 2014, 191, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Guenther, S.; Huwyler, D.; Richard, S.; Loessner, M.J. Virulent bacteriophage for efficient biocontrol of Listeria monocytogenes in ready-to-eat foods. Appl. Environ. Microbiol. 2009, 75, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Bueno, E.; García, P.; Martínez, B.; Rodríguez, A. Phage inactivation of Staphylococcus aureus in fresh and hard-type cheeses. Int. J. Food Microbiol. 2012, 158, 23–27. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; García, P. Bacteriophages as weapons against bacterial biofilms in the food industry. Front. Microbiol. 2016, 7, 825. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples Conditions | Log PFU/mL | Reduction Effectiveness (Log PFU/mL) |
|---|---|---|
| AP/RT/5 min | 8.94 ± 0.11 | - |
| AP/RT/20 min | 8.90 ± 0.05 | - |
| AP/RT/30 min | 8.91 ± 0.08 | - |
| 75 MPa/5 min | 8.65 ± 0.06 | 0.29 a,d,e,f |
| 75 MPa/20 min | 8.44 ± 0.05 | 0.46 a,d,e,f |
| 75 MPa/30 min | 8.30 ± 0.24 | 0.61 a,d,e,f |
| 200 MPa/5 min | 6.50 ± 0.18 | 2.44 b,c,e,f |
| 200 MPa/20 min | 6.34 ± 0.03 | 2.56 b,c,e,f |
| 200 MPa/30 min | 6.20 ± 0.02 | 2.71 b,c,e,f |
| 300 MPa/5 min | 5.81 ± 0.04 | 3.13 b,c,d,f |
| 300 MPa/20 min | 5.45 ± 0.03 | 3.45 b,c,d,f |
| 300 MPa/30 min | 4.65 ± 0.08 | 4.26 b,c,d,f |
| 400 MPa/5 min | 4.00 ± 0.12 | 4.94 b,c,d,e |
| 400 MPa/20 min | 2.64 ± 0.08 | 6.26 b,c,d,e |
| 400 MPa/30 min | ND | 7.91 b,c,d,e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, C.; Marques, J.F.; Reis, S.; Costa, P.; Martins, A.P.; Pinto, C.A.; Saraiva, J.A.; Almeida, A. Combined Effect of Phage phT4A and Pressure-Based Strategies in the Inhibition of Escherichia coli. Antibiotics 2022, 11, 211. https://doi.org/10.3390/antibiotics11020211
Pereira C, Marques JF, Reis S, Costa P, Martins AP, Pinto CA, Saraiva JA, Almeida A. Combined Effect of Phage phT4A and Pressure-Based Strategies in the Inhibition of Escherichia coli. Antibiotics. 2022; 11(2):211. https://doi.org/10.3390/antibiotics11020211
Chicago/Turabian StylePereira, Carla, João F. Marques, Sílvia Reis, Pedro Costa, Ana P. Martins, Carlos A. Pinto, Jorge A. Saraiva, and Adelaide Almeida. 2022. "Combined Effect of Phage phT4A and Pressure-Based Strategies in the Inhibition of Escherichia coli" Antibiotics 11, no. 2: 211. https://doi.org/10.3390/antibiotics11020211