
Antimicrobial Resistance Profile of Staphylococcus hyicus Strains Isolated from Brazilian Swine Herds

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Culture Conditions
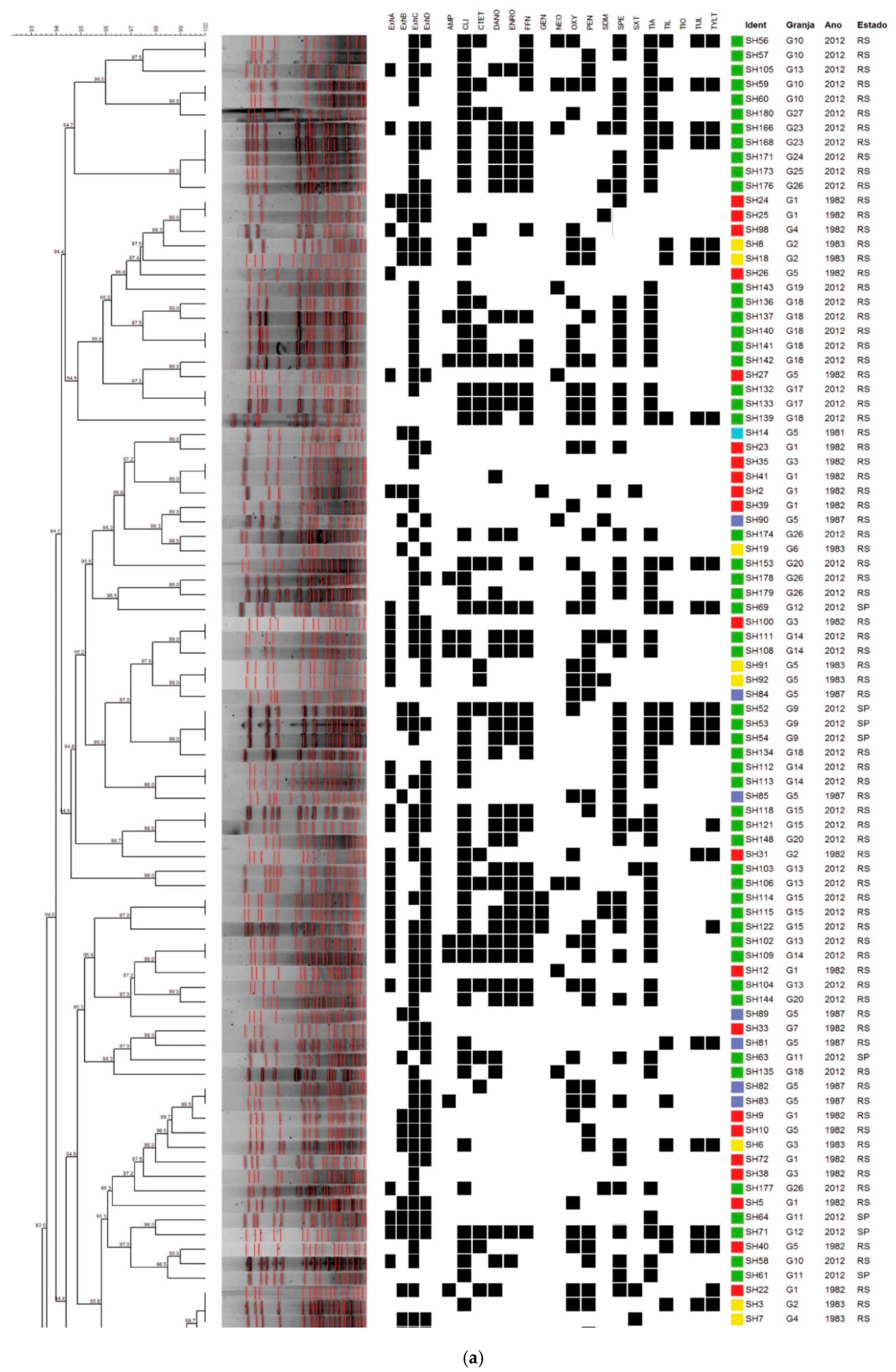
2.2. Detection of Genes Encoding Superoxide Dismutase A and Toxins SHETB, ExhA, ExhB, ExhC, and ExhD
2.3. Molecular Typing by PFGE
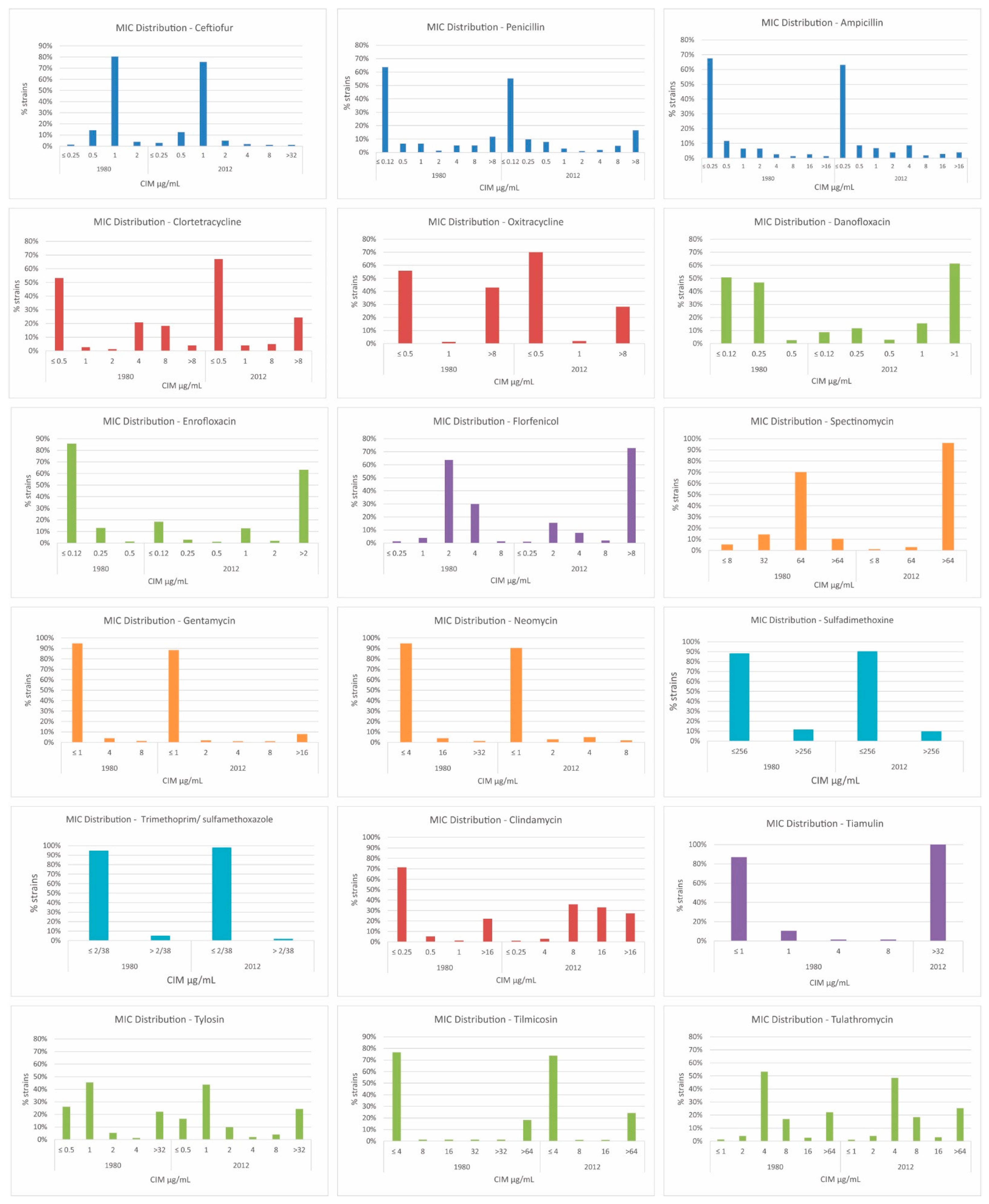
2.4. Broth Microdilution
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Aarestrup, F.M.; Jensen, L.B. Trends in antimicrobial susceptibility in relation to antimicrobial usage and presence of resistance genes in Staphylococcus hyicus isolated from exudative epidermitis in pigs. Vet. Microbiol. 2002, 89, 83–94. [Google Scholar] [CrossRef]
- Foster, A.P. Staphylococcal skin disease in livestock. Vet. Dermatol. 2012, 23, 342–351.e63. [Google Scholar] [CrossRef] [PubMed]
- Leekitcharoenphon, P.; Pamp, S.J.; Andresen, L.O.; Aarestrup, F.M. Comparative genomics of toxigenic and non-toxigenic Staphylococcus hyicus. Vet. Microbiol. 2016, 185, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, H.; Watanabe, T.; Higuchi, K.; Teruya, K.; Ohtake, A.; Murata, Y.; Saito, H.; Aizawa, C.; Danbara, H.; Maehara, N. Chromosomal and Extrachromosomal Synthesis of Exfoliative Toxin from Staphylococcus hyicus. J. Bacteriol. 2000, 182, 4096–4100. [Google Scholar] [CrossRef] [Green Version]
- Andresen, L.; Ahrens, P. A multiplex PCR for detection of genes encoding exfoliative toxins from Staphylococcus hyicus. J. Appl. Microbiol. 2004, 96, 1265–1270. [Google Scholar] [CrossRef]
- Fudaba, Y.; Nishifuji, K.; Andresen, L.O.; Yamaguchi, T.; Komatsuzawa, H.; Amagai, M.; Sugai, M. Staphylococcus hyicus exfoliative toxins selectively digest porcine desmoglein 1. Microb. Pathog. 2005, 39, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Nishifuji, K.; Fudaba, Y.; Yamaguchi, T.; Iwasaki, T.; Sugai, M.; Amagai, M. Cloning of swine desmoglein 1 and its direct proteolysis by Staphylococcus hyicus exfoliative toxins isolated from pigs with exudative epidermitis. Vet. Dermatol. 2005, 16, 315–323. [Google Scholar] [CrossRef]
- Devriese, L.A. Isolation and identification of Staphylococcus hyicus. Am. J. Vet. Res. 1977, 38, 787–792. [Google Scholar]
- Quinn, P.J.; Carter, M.E.; Markey, B.; Carter, G.R. Clinical Veterinary Microbiology; Wolfe: London, UK, 1994; pp. 137–143. [Google Scholar]
- Voytenko, A.; Kanbar, T.; Alber, J.; Lammler, C.; Weiss, R.; Prenger-Berninghoff, E.; Zschöck, M.; Akineden, O.; Hassan, A.; Dmitrenko, O. Identification of Staphylococcus hyicus by polymerase chain reaction mediated amplification of species specific sequences of superoxide dismutase A encoding gene sodA. Vet. Microbiol. 2006, 116, 211–216. [Google Scholar] [CrossRef]
- Kanbar, T.; Voytenko, A.V.; Alber, J.; Lämmler, C.; Weiss, R.; Skvortzov, V.N. Distribution of the putative virulence factor encoding gene sheta in Staphylococcus hyicus strains of various origins. J. Vet. Sci. 2008, 9, 327–329. [Google Scholar] [CrossRef]
- Boom, R.; Sol, C.J.; Salimans, M.M.; Jansen, C.L.; Dillen, P.M.W.-V.; van der Noordaa, J. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassler, C.; Nitzsche, S.; Iversen, C.; Zweifel, C.; Stephan, R. Characteristics of Staphylococcus hyicus strains isolated from pig carcasses in two different slaughterhouses. Meat Sci. 2008, 80, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI Supplement VET08. CLSI Document VET08; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Document M100S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Hu, Y.; Liu, L.; Zhang, X.; Feng, Y.; Zong, Z. In Vitro Activity of Neomycin, Streptomycin, Paromomycin and Apramycin against Carbapenem-Resistant Enterobacteriaceae Clinical Strains. Front. Microbiol. 2017, 8, 2275. [Google Scholar] [CrossRef] [Green Version]
- Rønne, H.; Szancer, J. In vitro susceptibility of Danish field isolates of Treponema hyodysenteriae to chemotherapeutics in swine dysentery (SD) therapy. Interpretation of MIC results based on the pharmacokinetic properties of the antibacterial agents. In Proceedings of the 11th International Pig Veterinary Society Congress, Swiss Association of Swine Medicine, Berne, Switzerland, 1–5 July 1990; p. 1126. [Google Scholar]
- van Belkum, A.; Tassios, P.T.; Dijkshoorn, L.; Haeggman, S.; Cookson, B.; Fry, N.; Fussing, V.; Green, J.; Feil, E.; Gerner-Smidt, P.; et al. Guidelines for the validation and application of typing methods for use in bacterial epidemiology. Clin. Microbiol. Infect. 2007, 13 (Suppl. 3), 1–46. [Google Scholar] [CrossRef]
- Futagawa-Saito, K.; Ba-Thein, W.; Fukuyasu, T. Antimicrobial Susceptibilities of Exfoliative Toxigenic and Non-Toxigenic Staphylococcus hyicus Strains in Japan. J. Vet. Med. Sci./Jpn. Soc. Vet. Sci. 2009, 71, 681–684. [Google Scholar] [CrossRef] [Green Version]
- Andresen, L.O. Production of exfoliative toxin by isolates of Staphylococcus hyicus from different countries. Vet. Rec. 2005, 157, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Dutra, M.; Moreno, L.; Dias, R.; Moreno, A. Antimicrobial Use in Brazilian Swine Herds: Assessment of Use and Reduction Examples. Microorganisms 2021, 9, 881. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Friendship, R.M.; Weese, J.S.; Poljak, Z.; Dewey, C.E. An investigation of resistance to β-lactam antimicrobials among staphylococci isolated from pigs with exudative epidermitis. BMC Vet. Res. 2013, 9, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegener, H.C.; Watts, J.L.; Salmon, S.A.; Yancey, R.J. Antimicrobial Susceptibility of Staphylococcus hyicus Isolated from Exudative Epidermitis in Pigs. J. Clin. Microbiol. 1994, 32, 793–795. Available online: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=263126&tool=pmcentrez&rendertype=abstract (accessed on 26 January 2022). [CrossRef] [Green Version]
- Aarestrup, F.M.; Duran, C.O.; Burch, D.G.S. Antimicrobial resistance in swine production. Anim. Health Res. Rev./Conf. Res. Work. Anim. Dis. 2008, 9, 135–148. [Google Scholar] [CrossRef]
- Wegener, H.C.; Schwarz, S. Antibiotic-resistance and plasmids in Staphylococcus hyicus isolated from pigs with exudative epidermitis and from healthy pigs. Vet. Microbiol. 1993, 34, 363–372. [Google Scholar] [CrossRef]
- Li, B.; Wendlandt, S.; Yao, J.; Liu, Y.; Zhang, Q.; Shi, Z.; Wei, J.; Shao, D.; Schwarz, S.; Wang, S.; et al. Detection and new genetic environment of the pleuromutilin-lincosamide-streptogramin A resistance gene lsa(E) in methicillin-resistant Staphylococcus aureus of swine origin. J. Antimicrob. Chemother. 2013, 68, 1251–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witte, W.; Cuny, C. Emergence and spread of cfr-mediated multiresistance in staphylococci: An interdisciplinary challenge. Future Microbiol. 2011, 6, 925–931. [Google Scholar] [CrossRef] [PubMed]
- MAPA. Ministério da Agricultura Pecuária e Abastecimento. Instrução Normativa Nº 1, de 13 de Janeiro de 2020. Available online: https://pesquisa.in.gov.br/imprensa/jsp/visualiza/index.jsp?jornal=515&pagina=6&data=23/01/2020 (accessed on 23 January 2021).




{kind=link}
{kind=link}
{kind=link}
| Antimicrobial | MIC Range (µg/mL) | MIC Breakpoints | ||
|---|---|---|---|---|
| Susceptible | Intermediary | Resistant | ||
| Ampicillin | ≤0.25–1.0 | ≤0.25 | 0.5 | ≥1 |
| Ceftiofur | ≤0.25–2.0 | ≤2 | 4 | ≥8 |
| Penicillin | ≤0.12–2.0 | ≤0.12 | - | ≥0.25 |
| Chlortetracycline | ≤0.5–>8.0 | ≤0.5 | 1 | ≥2 |
| Oxitetracycline | ≤0.5–>8.0 | ≤0.5 | 1 | ≥2 |
| Danofloxacin | 0.5–>1.0 | ≤0.25 | 0.5 | ≥1 |
| Enrofloxacin | 0.5–>2.0 | ≤0.5 | 1 | ≥2 |
| Florfenicol | 1.0–>8.0 | ≤2 | 4 | ≥8 |
| Spectinomycin | 16.0–>64.0 | ≤32 | 64 | ≥128 |
| Gentamycin | ≤1.0–>16.0 | ≤2 | 4 | ≥8 |
| Neomycin | ≤4.0–>32.0 | ≤8 | - | - |
| Sulfadimethoxine | >256.0 | ≤256 | - | ≥512 |
| Trimethoprim/sulfamethoxazole | >2/38 | ≤2/38 | - | ≥4/76 |
| Clindamycin | ≤0.25–>16.0 | ≤0.5 | 1–2 | ≥4 |
| Tylosin | ≤0.5–>32.0 | ≤1 | 2–4 | ˃4 |
| Tilmicosin | ≤4.0–>64.0 | ≤16 | - | ≥32 |
| Tulathromycin | ≤1.0–>64.0 | ≤16 | 32 | ≥64 |
| Tiamulin | 1.0–>32.0 | ≤16 | - | ≥32 |
| Toxins | 1980 | 2012 | p | ||
|---|---|---|---|---|---|
| N | % | N | % | ||
| ExhA | 17 | 22.08 | 45 | 43.68 | <0.001 |
| ExhB | 31 | 40.26 | 13 | 12.62 | <0.001 |
| ExhC | 55 | 71.40 | 82 | 79.61 | 0.150 |
| ExhD | 48 | 62.30 | 50 | 48.54 | 0.087 |
| Class | Antimicrobial | 1980 | 2012 | p | ||
|---|---|---|---|---|---|---|
| N | (%) | N | (%) | |||
| Beta-lactams | Ampicillin | 16 | 20.77 | 29 | 28.15 | 0.258 |
| Ceftiofur | 0 | 0.00 | 1 | 0.97 | 0.386 | |
| Penicillin | 28 | 36.36 | 46 | 44.66 | 0.263 | |
| Tetracycline | Oxitetracycline | 33 | 42.86 | 29 | 28.16 | 0.040 |
| Chlortetracycline | 34 | 44.15 | 30 | 29.12 | 0.037 | |
| Fluoroquinolones | Danofloxacin | 2 | 2.60 | 81 | 78.64 | <0.001 |
| Enrofloxacin | 0 | 0.00 | 67 | 65.05 | <0.001 | |
| Aminoglycosides | Gentamycin | 1 | 1.30 | 9 | 8.74 | 0.045 |
| Neomycin | 4 | 5.19 | 10 | 9.71 | 0.263 | |
| Spectinomycin | 8 | 10.4 | 99 | 96.1 | <0.001 | |
| Fenicois | Florfenicol | 1 | 1.30 | 77 | 74.76 | <0.001 |
| Sulfas | Sulfadimethoxine | 9 | 11.69 | 10 | 9.71 | 0.669 |
| Trimethoprim/sulfamethoxazole | 4 | 5.19 | 2 | 1.94 | 0.229 | |
| Lincosamides | Clindamycin | 17 | 22.08 | 102 | 99.03 | <0.001 |
| Pleuromutilins | Tiamulin | 0 | 0.00 | 103 | 100.00 | <0.001 |
| Macrolides | Tilmicosin | 16 | 20.78 | 25 | 24.27 | 0.580 |
| Tylosin | 18 | 23.38 | 31 | 30.10 | 0.316 | |
| Tulathromycin | 17 | 22.08 | 26 | 25.24 | 0.622 | |
| Classification | 1980 | 2012 | p | ||
|---|---|---|---|---|---|
| N | % | N | % | ||
| Resistant to 2 classes or less | 47 | 61.0 | 1 | 1.0 | <0.001 |
| Multidrug resistant (3 classes or more) | 30 | 39.0 | 102 | 99.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, A.M.; Moreno, L.Z.; Poor, A.P.; Matajira, C.E.C.; Moreno, M.; Gomes, V.T.d.M.; da Silva, G.F.R.; Takeuti, K.L.; Barcellos, D.E. Antimicrobial Resistance Profile of Staphylococcus hyicus Strains Isolated from Brazilian Swine Herds. Antibiotics 2022, 11, 205. https://doi.org/10.3390/antibiotics11020205
Moreno AM, Moreno LZ, Poor AP, Matajira CEC, Moreno M, Gomes VTdM, da Silva GFR, Takeuti KL, Barcellos DE. Antimicrobial Resistance Profile of Staphylococcus hyicus Strains Isolated from Brazilian Swine Herds. Antibiotics. 2022; 11(2):205. https://doi.org/10.3390/antibiotics11020205
Chicago/Turabian StyleMoreno, Andrea Micke, Luisa Zanolli Moreno, André Pegoraro Poor, Carlos Emilio Cabrera Matajira, Marina Moreno, Vasco Túlio de Moura Gomes, Givago Faria Ribeiro da Silva, Karine Ludwig Takeuti, and David Emilio Barcellos. 2022. "Antimicrobial Resistance Profile of Staphylococcus hyicus Strains Isolated from Brazilian Swine Herds" Antibiotics 11, no. 2: 205. https://doi.org/10.3390/antibiotics11020205