
Modeling the Directed Evolution of Broad Host Range Phages

Abstract
:1. Introduction
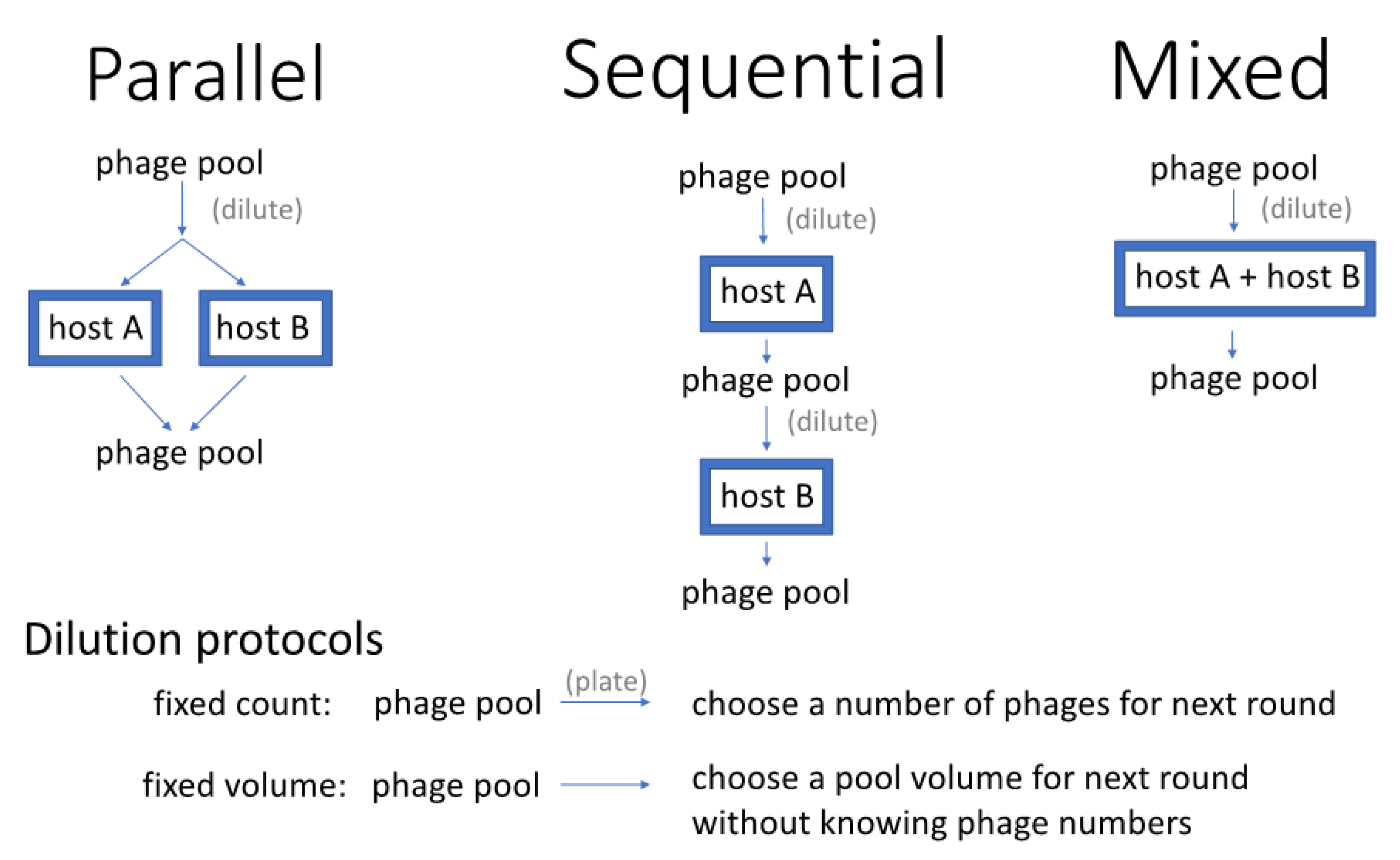
- Sequential presentation. Phages are grown on one host, then the pool of phages from that host is transferred to another host, and so on [20]. The process continues by rotating through each host and is then repeated. In one variation of this protocol, actual plaques are chosen for propagation from one host to the next [20].
- Mixed presentation. Phages are grown in a mix of all hosts in a single culture. The bacterial mix is periodically or continually refreshed to ensure the continued presence of all hosts while maintaining the supernatant containing phage. This method originates with d’Herelle [7] using one non-permissive and one permissive host [as attributed by [11], p. 129].
2. Methods
2.1. Growth Dynamics
2.2. Dilution Protocols
2.3. A Historical Note
3. Results
3.1. Heuristic Models
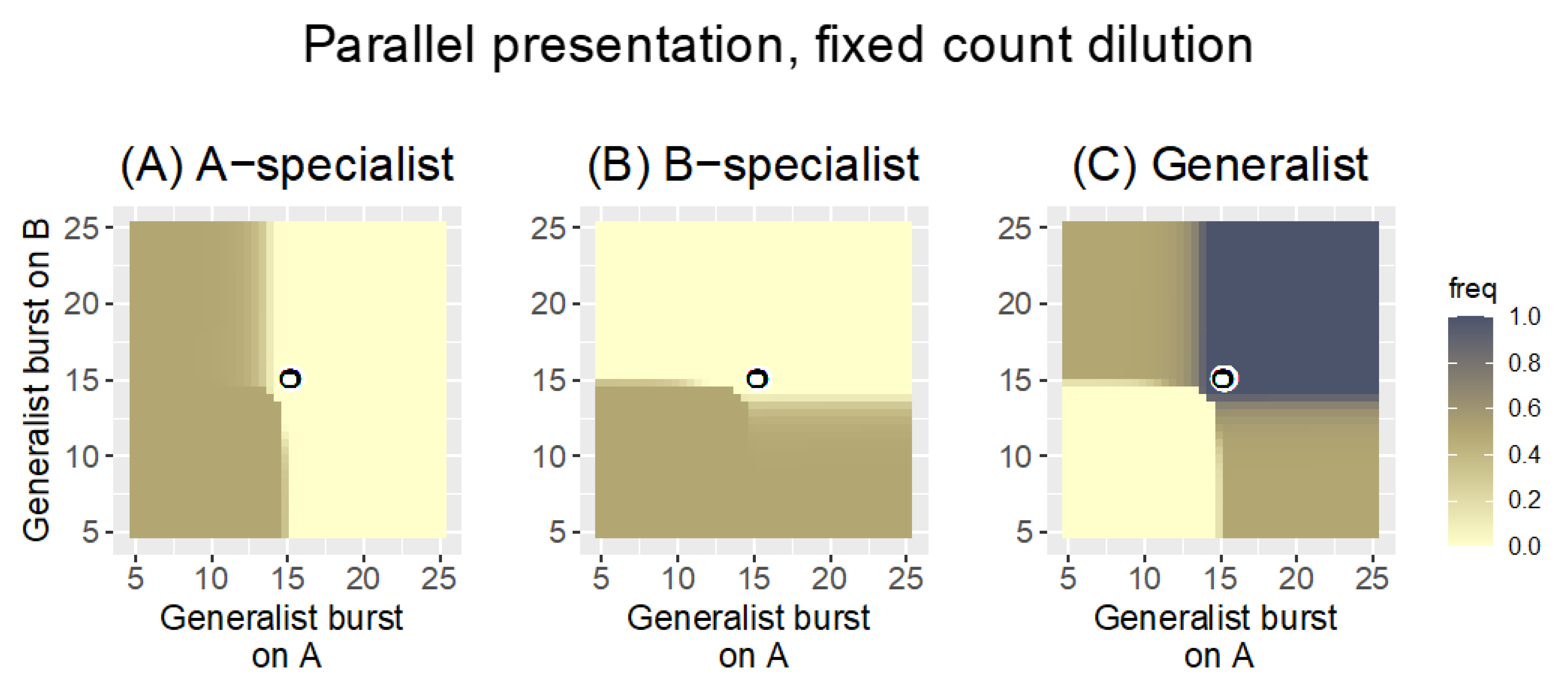
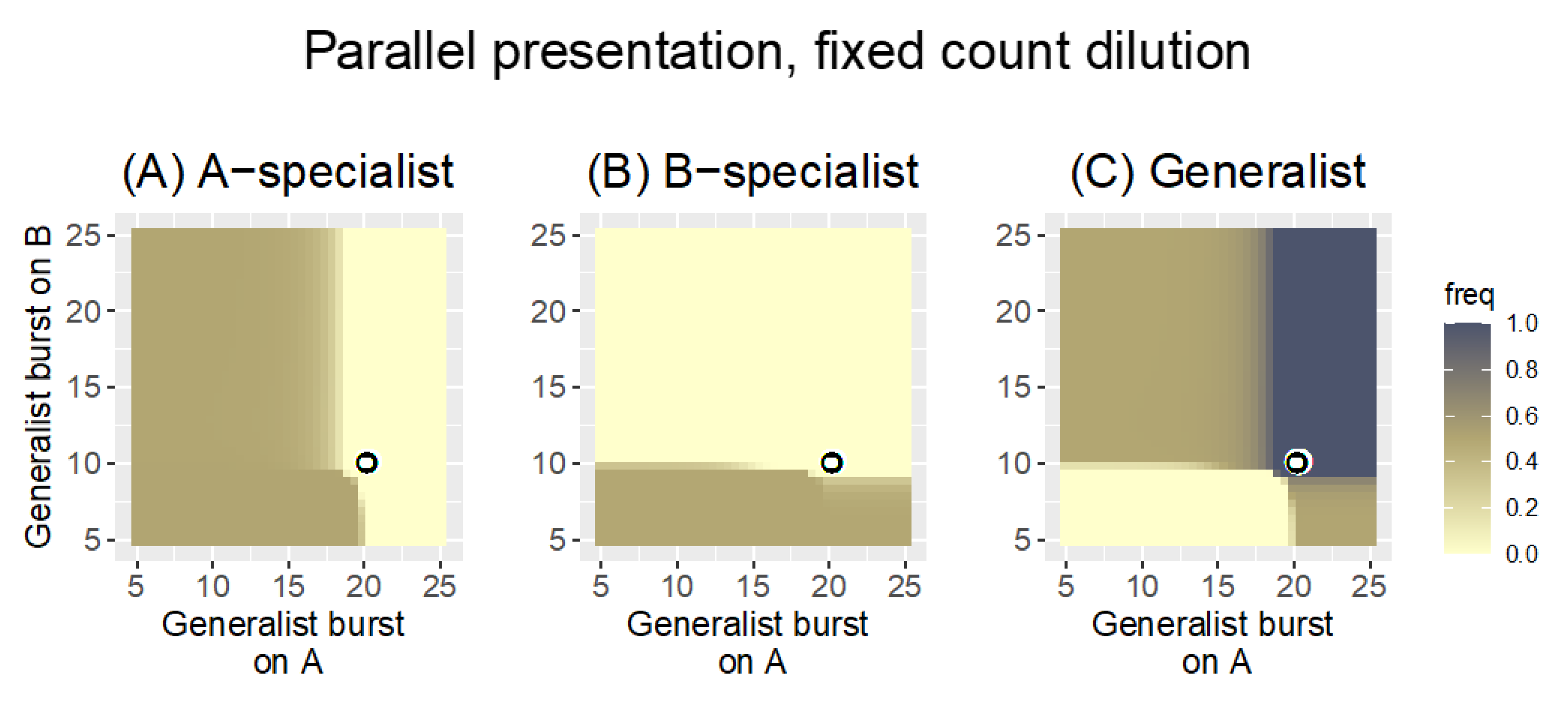
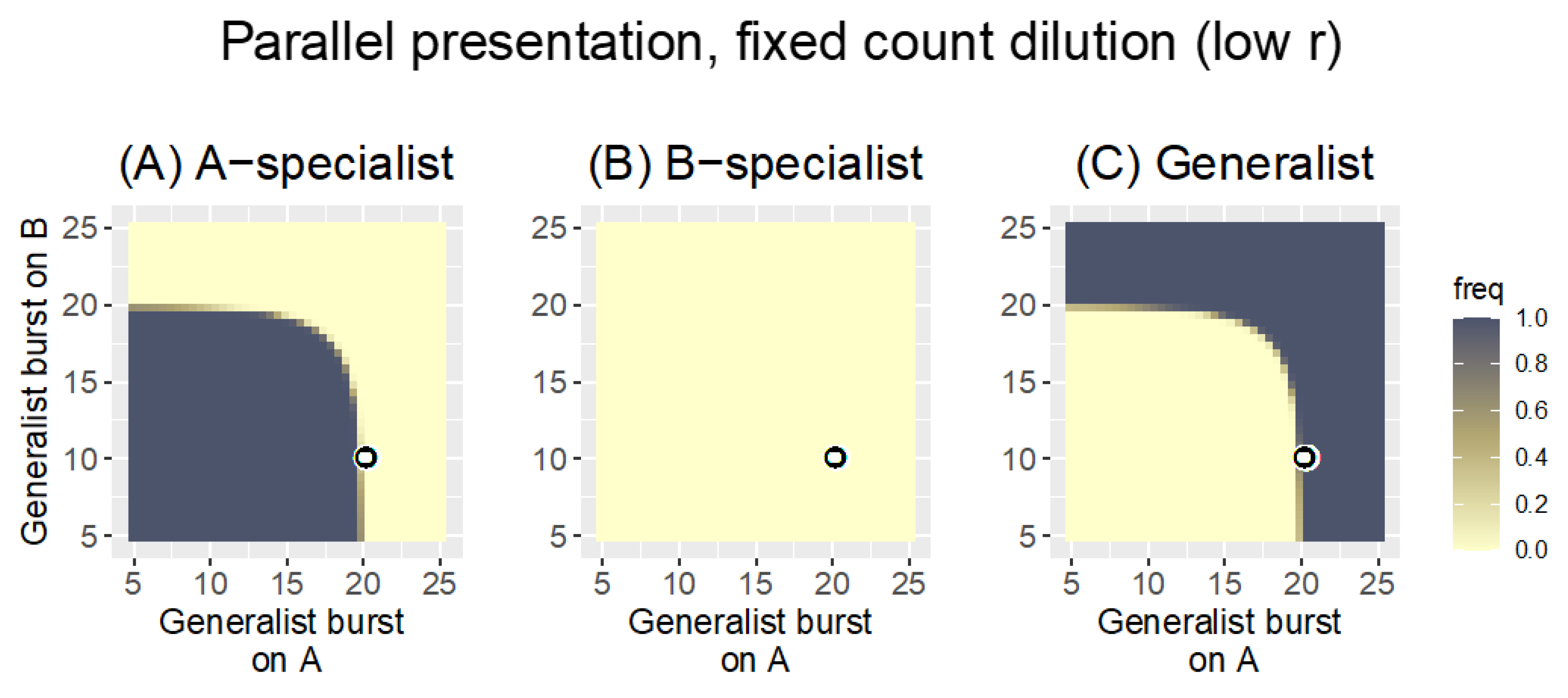
3.1.1. Parallel Presentation
3.1.2. Sequential Presentation
3.1.3. Mixed Presentation
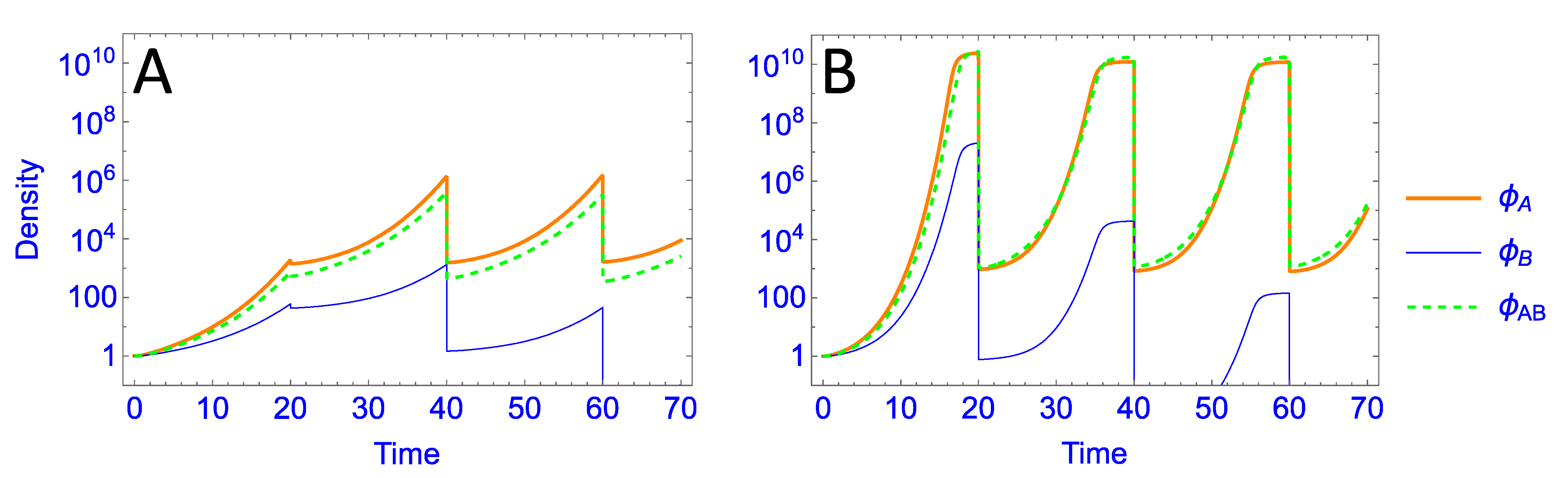
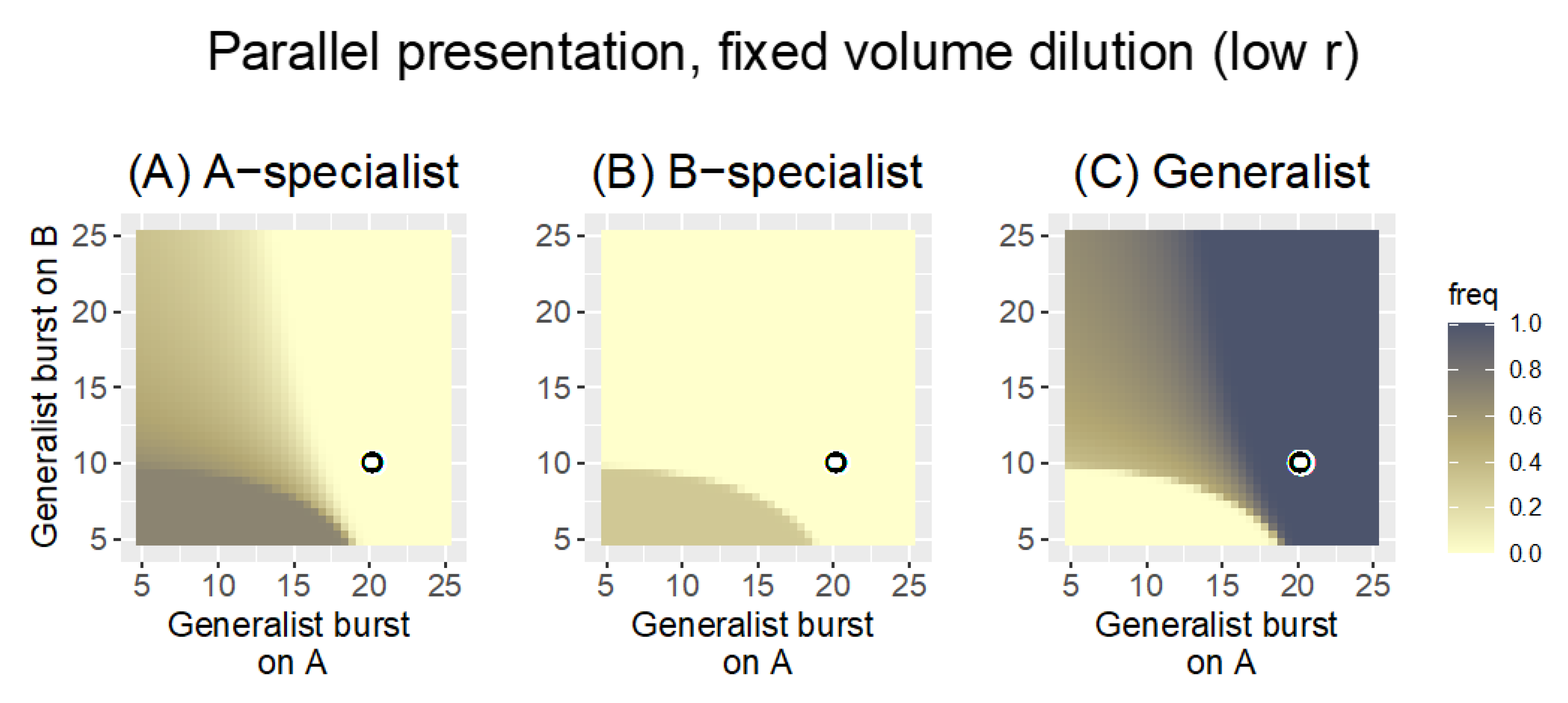
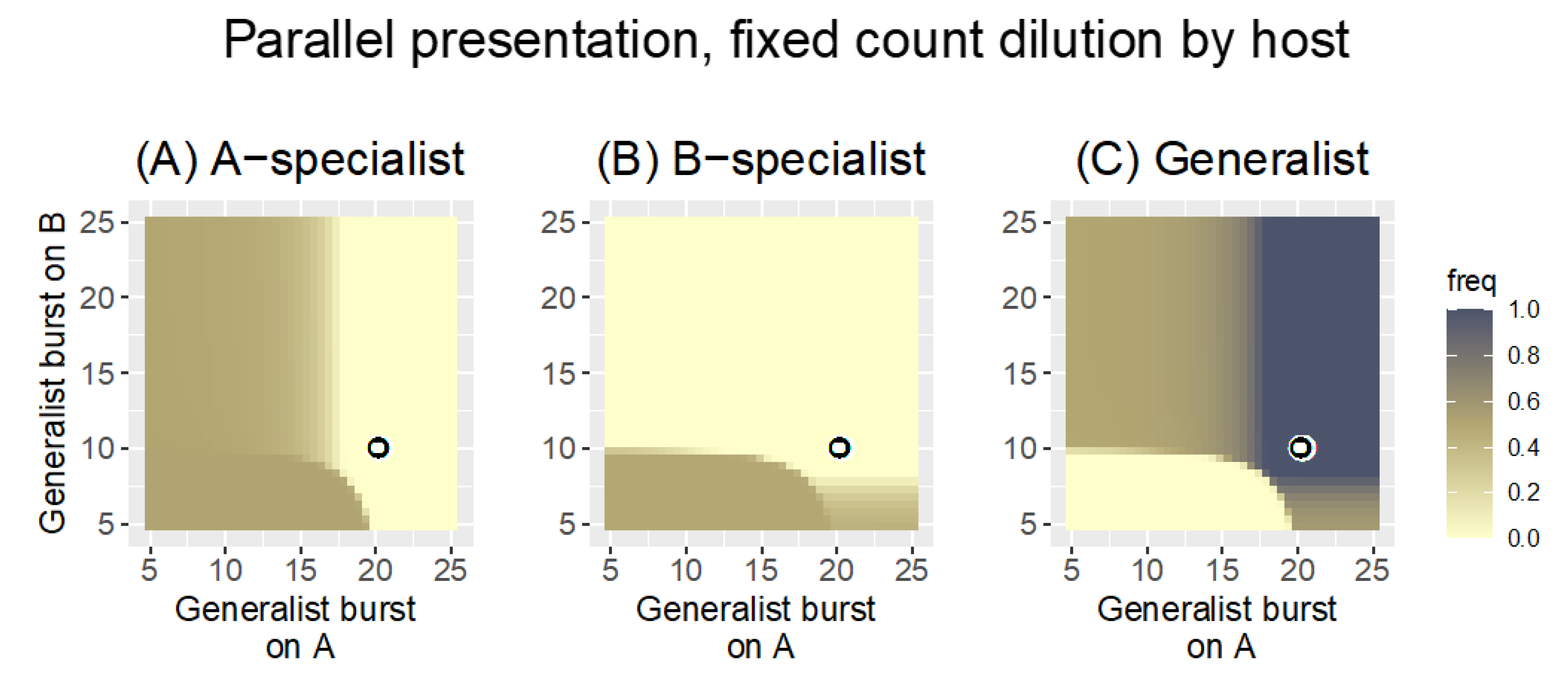
3.2. Numerical Analyses
3.2.1. Parallel Presentation
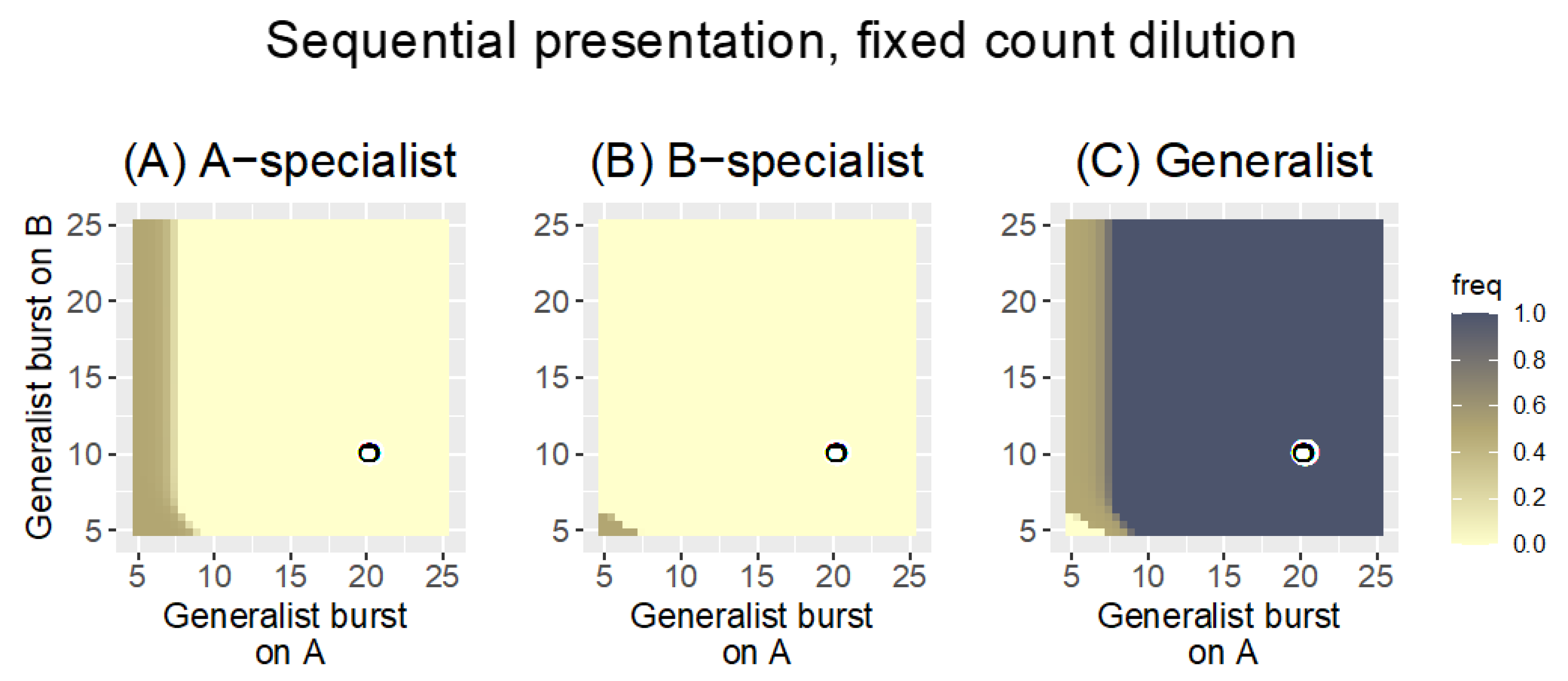
3.2.2. Sequential Presentation
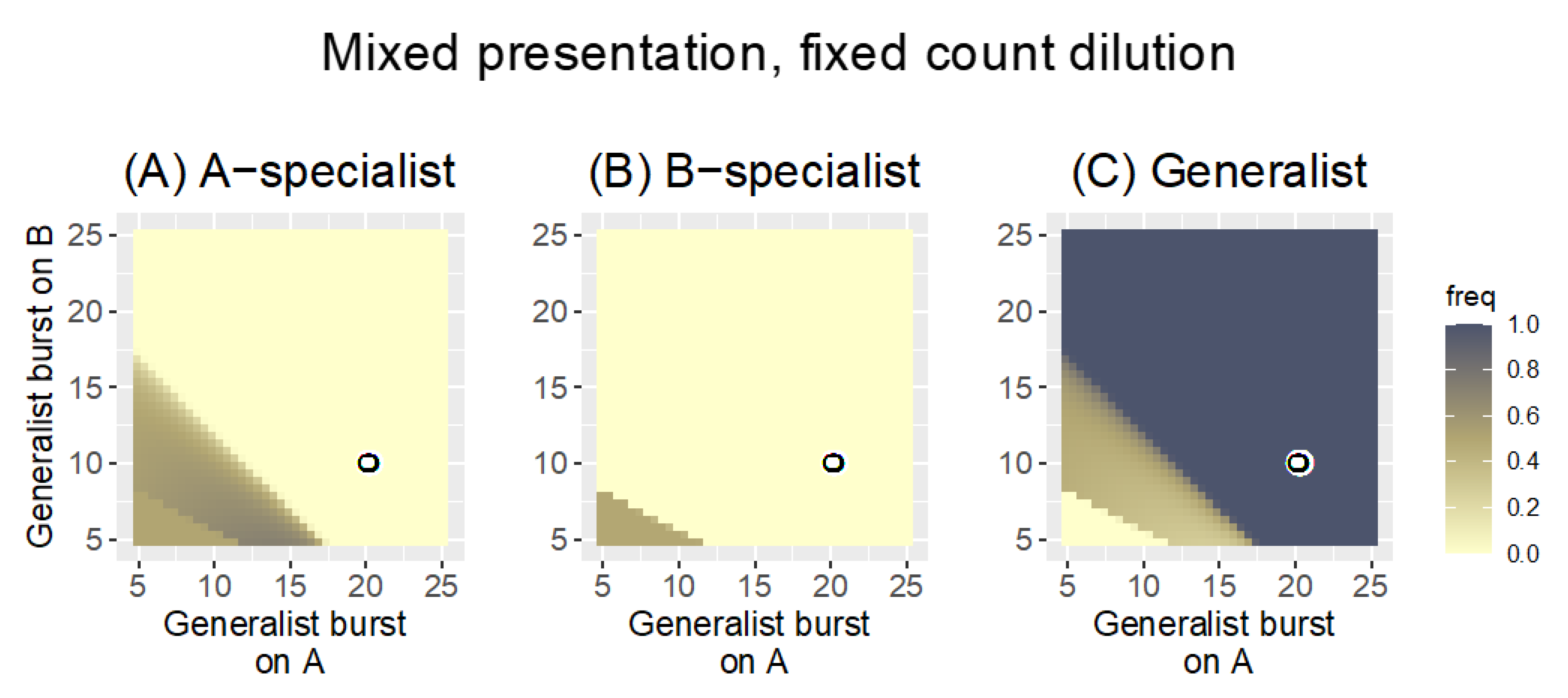
3.2.3. Mixed Presentation
3.3. Summary
4. Discussion and Conclusions
4.1. Other Effects of Protocol on Evolution
4.1.1. Recombination
4.1.2. Rare Mutations Are Likely to Be Lost
4.2. Recommendations
- (1)
- Choose an appropriate presentation protocol. Although the protocols vary in how deferential they are to generalist phages, the utility of generalist over specialist phages will depend on the application. To take advantage of the variety of host-presentation protocols available, the protocol could be tailored to the application. For example, sequential presentation of hosts is unlikely to exist in infections (except possibly if host resistance evolves following treatment), whereas both Parallel and Mixed protocols could directly match infections. In turn, this would suggest that a Sequential protocol was not the best choice for evolving phages to treat infections. Phages evolved under Mixed presentation might be better suited to treating a mixed infection than would phages evolved under other protocols, and so on.
- (2)
- Recombination. Although much remains to understand the general importance of recombination to host range evolution, there may be little lost by encouraging recombination in a protocol. Recombination is promoted by using multiple parental phages and ensuring they are maintained at moderate frequencies. However, including different phages at the start may offer only a transient boost to recombination. The propagation of different phages on a permissive host will typically result in numerical dominance by one phage [38], so the opportunities for recombination may dissipate quickly. Periodically supplementing the phage pool with the original phage stock will maintain opportunities for recombination. Furthermore, the phage pool itself may be created with recombination so that the pool is not only supplemented with parental phages but also supplemented with recombinants among the parents. Recombination rates also depend on the sequence similarities among starting phages.
- (3)
- Early losses. If mutations are limiting, it may be beneficial to modify the protocol to reduce phage dilution and avoid phages being trapped on an inappropriate host—the latter a greater problem with Parallel presentation than, say, Mixed presentation. In the Appelmans protocol, for example, this would involve reducing the number of non-permissive hosts. Concentrating phages at the end of a cycle would reduce dilution. Alternatively, the protocol may be modified to increase the number of recombinants and mutants (as with mutagens), so that early loss is inconsequential.
- (4)
- The initial pool. A diversity in the initial pool of phages will surely increase the range of possible outcomes. As noted above, the initial pool will affect opportunities for recombination. We yet can offer little insight on whether a diverse starting pool will affect the outcome toward generalists or specialists. Starting with a single phage might seem to ensure the evolution of a generalist, but selection is agnostic to phage origins, and a single phage might well diversify into a suite of specialists.
- (5)
- Dilution. There is a risk from too high dilution in a protocol—demographic extinction of phages that would otherwise evolve. However, low dilution has its own problems. First, low dilution slows evolution merely because there is less phage growth per cycle and thus less opportunity for fitness differences to manifest. Low dilution could thus greatly increase the empirical work associated with evolving phage host range. Second, low dilution results in most of the phage growth at high multiplicity. This can facilitate the evolution of phages that are good at within-host competition at the expense of independent growth [42,43]. High dilution will usually be desirable, and any demographic effects can be offset by extending the culture duration.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bielke, L.; Higgins, S.; Donoghue, A.; Donoghue, D.; Hargis, B.M. Salmonella host range of bacteriophages that infect multiple genera. Poult. Sci. 2007, 86, 2536–2540. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P.; Abedon, S.T. Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 2010, 70, 217–248. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, P.A.; Nobrega, F.L.; Brouns, S.J.J.; Dutilh, B.E. Molecular and Evolutionary Determinants of Bacteriophage Host Range. Trends Microbiol. 2019, 27, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Cazares, D.; Cazares, A.; Figueroa, W.; Guarneros, G.; Edwards, R.A.; Vinuesa, P. A novel group of promiscuous podophages infecting diverse gammaproteobacteria from river communities exhibits dynamic intergenus host adaptation. Msystems 2021, 6, e00773-20. [Google Scholar] [CrossRef] [PubMed]
- Fong, K.; Wong, C.W.Y.; Wang, S.; Delaquis, P. How Broad Is Enough: The Host Range of Bacteriophages and Its Impact on the Agri-Food Sector. Ther. Appl. Res. 2021, 2, 83–91. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- D’Herelle, F. The Bacteriophage and Its Behavior; Williams & Wilkins: Philadelphia, PA, USA, 1926. [Google Scholar]
- Dublanchet, A.; Fruciano, E. A short history of phage therapy. Med. Mal. Infect. 2008, 38, 415–420. [Google Scholar] [CrossRef]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef]
- Zagaliotis, P.; Michalik-Provasek, J.; Gill, J.J.; Walsh, T.J. Therapeutic Bacteriophages for Gram-Negative Bacterial Infections in Animals and Humans. Pathog. Immun. 2022, 7, 1–45. [Google Scholar] [CrossRef]
- Adams, M.H. Bacteriophages; Interscience Publishers: New York, NY, USA, 1959. [Google Scholar]
- Ross, A.; Ward, S.; Hyman, P. More Is Better: Selecting for Broad Host Range Bacteriophages. Front. Microbiol. 2016, 7, 1352. [Google Scholar] [CrossRef]
- Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Dedrick, R.M.; Freeman, K.G.; Nguyen, J.A.; Bahadirli-Talbott, A.; Smith, B.E.; Wu, A.E.; Ong, A.S.; Lin, C.T.; Ruppel, L.C.; Parrish, N.M.; et al. Potent antibody-mediated neutralization limits bacteriophage treatment of a pulmonary Mycobacterium abscessus infection. Nat. Med. 2021, 27, 1357–1361. [Google Scholar] [CrossRef]
- El Haddad, L.; Harb, C.P.; Gebara, M.A.; Stibich, M.A.; Chemaly, R.F. A Systematic and Critical Review of Bacteriophage Therapy Against Multidrug-resistant ESKAPE Organisms in Humans. Clin. Infect. Dis. 2019, 69, 167–178. [Google Scholar] [CrossRef]
- Burrowes, B.H.; Molineux, I.J.; Fralick, J.A. Directed in Vitro Evolution of Therapeutic Bacteriophages: The Appelmans Protocol. Viruses 2019, 11, 241. [Google Scholar] [CrossRef] [Green Version]
- Mapes, A.C.; Trautner, B.W.; Liao, K.S.; Ramig, R.F. Development of expanded host range phage active on biofilms of multi-drug resistant Pseudomonas aeruginosa. Bacteriophage 2016, 6, e1096995. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Mathieu, J.; Li, M.; Dai, Z.; Alvarez, P.J.J. Isolation of polyvalent bacteriophages by sequential multiple-host approaches. Appl. Environ. Microbiol. 2016, 82, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Hashemolhosseini, S.; Holmes, Z.; Mutschler, B.; Henning, U. Alterations of receptor specificities of coliphages of the T2 family. J. Mol. Biol. 1994, 240, 105–110. [Google Scholar] [CrossRef]
- Hashemolhosseini, S.; Montag, D.; Krämer, L.; Henning, U. Determinants of receptor specificity of coliphages of the T4 family. A chaperone alters the host range. J. Mol. Biol. 1994, 241, 524–533. [Google Scholar] [CrossRef]
- Weitz, J. Quantitative Viral Ecology: Dynamics of Viruses and Their Microbial Hosts, 1st ed.; Princeton University Press: Princeton, NJ, USA, 2016; ISBN 978-0-691-16154-9. [Google Scholar]
- Levin, B.R.; Stewart, F.M.; Chao, L. Resource-limited growth, competition, and predation: A model and experimental studies with bacteria and bacteriophage. Am. Nat. 1977, 977, 3–24. [Google Scholar] [CrossRef]
- Bull, J.J.; Heineman, R.H.; Wilke, C.O. The phenotype-fitness map in experimental evolution of phages. PLoS ONE 2011, 6, e27796. [Google Scholar] [CrossRef] [Green Version]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Applemans, R. Le dosage du bactériophage. Compt. Rend. Soc. Biol. 1921, 85, 1098. [Google Scholar]
- Yehl, K.; Lemire, S.; Yang, A.C.; Ando, H.; Mimee, M.; Torres, M.D.T.; de la Fuente-Nunez, C.; Lu, T.K. Engineering phage host-range and suppressing bacterial resistance through phage tail fiber mutagenesis. Cell 2019, 179, 459–469.e9. [Google Scholar] [CrossRef] [PubMed]
- Latka, A.; Lemire, S.; Grimon, D.; Dams, D.; Maciejewska, B.; Lu, T.; Drulis-Kawa, Z.; Briers, Y. Engineering the modular receptor-binding proteins of Klebsiella phages switches their capsule serotype specificity. Cell 2021, 12, e00455-21. [Google Scholar] [CrossRef] [PubMed]
- Kerr, B.; Neuhauser, C.; Bohannan, B.J.M.; Dean, A.M. Local migration promotes competitive restraint in a host-pathogen “tragedy of the commons. ” Nature 2006, 442, 75–78. [Google Scholar] [CrossRef]
- Bull, J.J. Optimality models of phage life history and parallels in disease evolution. J. Theor. Biol. 2006, 241, 928–938. [Google Scholar] [CrossRef]
- Crill, W.D.; Wichman, H.A.; Bull, J.J. Evolutionary reversals during viral adaptation to alternating hosts. Genetics 2000, 154, 27–37. [Google Scholar] [CrossRef]
- Meyer, J.R.; Dobias, D.T.; Weitz, J.S.; Barrick, J.E.; Quick, R.T.; Lenski, R.E. Repeatability and contingency in the evolution of a key innovation in phage lambda. Science 2012, 335, 428–432. [Google Scholar] [CrossRef] [Green Version]
- Paff, M.L.; Stolte, S.P.; Bull, J.J. Lethal mutagenesis failure may augment viral adaptation. Mol. Biol. Evol. 2014, 31, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Botstein, D. A theory of modular evolution for bacteriophages. Ann. New York Acad. Sci. 1980, 354, 484–490. [Google Scholar] [CrossRef]
- Stemmer, W.P. Rapid evolution of a protein in vitro by DNA shuffling. Nature 1994, 370, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ning, H.; Lin, H.; She, J.; Wang, L.; Jing, Y.; Wang, J. Expansion of the Plaquing Host Range and Improvement of the Absorption Rate of a T5-like Salmonella Phage by Altering the Long Tail Fibers. Appl. Environ. Microbiol. 2022, 88, e0089522. [Google Scholar] [CrossRef] [PubMed]
- Korona, R.; Levin, B.R. Phage-mediated selection and the evolution and maintenance of restriction-modification. Evolution 1993, 47, 556–575. [Google Scholar] [CrossRef]
- Bull, J.J.; Badgett, M.R.; Springman, R.; Molineux, I.J. Genome properties and the limits of adaptation in bacteriophages. Evolution 2004, 58, 692–701. [Google Scholar] [CrossRef]
- Nguyen, A.H.; Molineux, I.J.; Springman, R.; Bull, J.J. Multiple genetic pathways to similar fitness limits during viral adaptation to a new host. Evolution 2012, 66, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Patwa, Z.; Wahl, L.M. Fixation probability for lytic viruses: The attachment-lysis model. Genetics 2008, 180, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Turner, P.E.; Chao, L. Prisoner’s dilemma in an RNA virus. Nature 1999, 398, 441–443. [Google Scholar] [CrossRef]
- Karki, B.; Bull, J.J.; Krone, S.M. Modeling the therapeutic potential of defective interfering particles in the presence of immunity. Virus Evol. 2022, 8, veac047. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Notation | Meaning | Values Used |
|---|---|---|
| density of phage i growing on host j (function of time) | > 0 | |
| density of host j (function of time) | > 0 | |
| density of host j infected with phage i (function of time) | > 0 | |
| lysis rate of | 1.0 | |
| number of phage progeny released at lysis from an infection | 5–25 | |
| adsorption rate constant of phage i onto cells of strain j | 10−9 | |
| rj | Growth rate of bacterial strain j | 0.1, 0.3 |
| Host A | Host B | ||
|---|---|---|---|
| Phage | ΦA | NA|A | 1 |
| ΦB | 1 | NB|B | |
| ΦAB | NAB|A | NAB|B |
| Phage | Descendants per Cycle |
|---|---|
| ΦA | NA|A +1 |
| ΦB | 1 + NB|B |
| ΦAB | NAB|A + NAB|B |
| Phage | Descendants per Cycle |
|---|---|
| ΦA | NA|A · 1 |
| ΦB | 1 · NB|B |
| ΦAB | NAB|A · NAB|B |
| Protocol | Dilution | Bacterial r | Outcome |
|---|---|---|---|
| Parallel | fixed count | 0.3 | Phages with the best growth on a host are retained; by and large, generalists have no specific advantage over specialists. |
| “ | fixed count | 0.1 | With the low bacterial growth rate, the fixed count dilution allows all phages growing on the poorer host to be lost; only one phage is maintained in the pool. |
| “ | fixed volume | 0.1 | Despite the low bacterial growth rate, the change in dilution protocol now retains phages for both hosts; the generalist has a modest advantage over specialists |
| “ | fixed count by host | 0.1 | The change in dilution protocol ensures that phages are retained for both hosts; the generalist has a slight advantage over specialists, but the method broadly rewards phage growth on a per-host basis |
| Sequential | fixed count | 0.3 | Generalists displace specialists except when the specialist is extremely superior |
| Mixed | fixed count | 0.3 | Generalists are strongly favored over specialists; the outcome is intermediate between Parallel and Sequential presentation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bull, J.J.; Wichman, H.A.; Krone, S.M. Modeling the Directed Evolution of Broad Host Range Phages. Antibiotics 2022, 11, 1709. https://doi.org/10.3390/antibiotics11121709
Bull JJ, Wichman HA, Krone SM. Modeling the Directed Evolution of Broad Host Range Phages. Antibiotics. 2022; 11(12):1709. https://doi.org/10.3390/antibiotics11121709
Chicago/Turabian StyleBull, James J., Holly A. Wichman, and Stephen M. Krone. 2022. "Modeling the Directed Evolution of Broad Host Range Phages" Antibiotics 11, no. 12: 1709. https://doi.org/10.3390/antibiotics11121709