

Physiological Characteristics of Putative Enterobacteria Associated with Meat and Fish Available in Southern Brazilian Retail Markets: Antimicrobial Susceptibility, Toxic Metal Tolerance and Expression of Efflux Pumps

Abstract
:1. Introduction
2. Results
2.1. Bacterial Isolation and Total Viable Counts
2.2. Antimicrobial Susceptibility Patterns
2.3. Tolerance to Toxic Metals
2.4. Phenotypic Expression of Efflux Mechanisms
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Microbiological Quantification
4.2. Characterization of Enterobacteria and Antimicrobial Susceptibility Patterns
4.3. Tolerance to Toxic Metals
4.4. Phenotypic Expression of Efflux Mechanisms
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kulasooriya, G.D.B.N.; Amarasiri, M.K.U.T.; Abeykoon, A.M.H.; Kalupahana, R.S. Salmonella, Campylobacter and Escherichia coli in Raw Chicken Meat, Chicken Products and Cooked Chicken in Retail Markets in Kandy, Sri Lanka. Sri Lanka Vet. J. 2019, 66, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.; Kumar, D.; Hussain, S.; Pathak, A.; Shukla, M.; Prasanna Kumar, V.; Anisha, P.N.; Rautela, R.; Upadhyay, A.K.; Singh, S.P. Prevalence, Antimicrobial Resistance and Virulence Genes Characterization of Nontyphoidal Salmonella Isolated from Retail Chicken Meat Shops in Northern India. Food Control 2019, 102, 104–111. [Google Scholar] [CrossRef]
- Heredia, N.; García, S. Animals as Sources of Food-Borne Pathogens: A Review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef]
- Yu, H.; Elbediwi, M.; Zhou, X.; Shuai, H.; Lou, X.; Wang, H.; Li, Y.; Yue, M. Epidemiological and Genomic Characterization of Campylobacter Jejuni Isolates from a Foodborne Outbreak at Hangzhou, China. Int. J. Mol. Sci. 2020, 21, 3001. [Google Scholar] [CrossRef]
- CDC Burden of Foodborne Illnesses in the United States|Estimates of Foodborne Illness|CDC. Available online: https://www.cdc.gov/foodborneburden/burden/index.html (accessed on 20 September 2022).
- Scharff, R.L. The Economic Burden of Foodborne Illness in the United States. In Food Safety Economics; Springer International Publishing: Cham, Switzerland, 2018; pp. 123–142. [Google Scholar]
- Hindermann, D.; Gopinath, G.; Chase, H.; Negrete, F.; Althaus, D.; Zurfluh, K.; Tall, B.D.; Stephan, R.; Nüesch-Inderbinen, M. Salmonella enterica Serovar Infantis from Food and Human Infections, Switzerland, 2010–2015: Poultry-Related Multidrug Resistant Clones and an Emerging ESBL Producing Clonal Lineage. Front. Microbiol. 2017, 8, 1322. [Google Scholar] [CrossRef] [Green Version]
- Riley, L.W. Extraintestinal Foodborne Pathogens. Annu. Rev. Food Sci. Technol. 2020, 11, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-Based Phylogeny and Taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales Ord. Nov. Divided into the Families Enterobacteriaceae, Erwiniaceae Fam. Nov., Pectobacteriaceae Fam. Nov., Yersiniaceae Fam. Nov., Hafniaceae Fam. Nov., Morganellaceae Fam. Nov., and Budviciaceae Fam. Nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar]
- Jackson, P.; Meah, A. Re-Assessing Vulnerability to Foodborne Illness: Pathways and Practices. Crit. Public Health 2018, 28, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Randall, L.P.; Lodge, M.P.; Elviss, N.C.; Lemma, F.L.; Hopkins, K.L.; Teale, C.J.; Woodford, N. Evaluation of Meat, Fruit and Vegetables from Retail Stores in Five United Kingdom Regions as Sources of Extended-Spectrum Beta-Lactamase (ESBL)-Producing and Carbapenem-Resistant Escherichia coli. Int. J. Food Microbiol. 2017, 241, 283–290. [Google Scholar] [CrossRef]
- Finley, R.L.; Collignon, P.; Larsson, D.G.J.; Mcewen, S.A.; Li, X.Z.; Gaze, W.H.; Reid-Smith, R.; Timinouni, M.; Graham, D.W.; Topp, E. The Scourge of Antibiotic Resistance: The Important Role of the Environment. Clin. Infect Diseases 2013, 57, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Wales, A.D.; Davies, R.H. Co-Selection of Resistance to Antibiotics, Biocides and Heavy Metals, and Its Relevance to Foodborne Pathogens. Antibiotics 2015, 4, 567–604. [Google Scholar] [CrossRef]
- Stecher, B.; Denzler, R.; Maier, L.; Bernet, F.; Sanders, M.J.; Pickard, D.J.; Barthel, M.; Westendorf, A.M.; Krogfelt, K.A.; Walker, A.W.; et al. Gut Inflammation Can Boost Horizontal Gene Transfer between Pathogenic and Commensal Enterobacteriaceae. Proc. Natl. Acad. Sci. USA 2012, 109, 1269–1274. [Google Scholar] [CrossRef] [Green Version]
- Schjørring, S.; Krogfelt, K.A. Assessment of Bacterial Antibiotic Resistance Transfer in the Gut. Int. J. Microbiol. 2011, 2011, 312956. [Google Scholar] [CrossRef] [Green Version]
- Nadimpalli, M.; Delarocque-Astagneau, E.; Love, D.C.; Price, L.B.; Huynh, B.T.; Collard, J.M.; Lay, K.S.; Borand, L.; Ndir, A.; Walsh, T.R.; et al. Combating Global Antibiotic Resistance: Emerging One Health Concerns in Lower-and Middle-Income Countries. Clin. Infect. Diseases 2018, 66, 963–969. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microb. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Brazil Ministry of Agriculture, Livestock and Supply. Métodos Oficiais para a Análise de Produtos de Origem Animal; Ministry of Agriculture, Livestock and Supply: Brasília, Brazil, 2022; ISBN 978-85-7991-155-2.
- Podpečan, B.; Pengov, A.; Vadnjal, S. The Source of Contamination of Ground Meat for Production of Meat Products with Bacteria Staphylococcus aureus. Slov. Vet. Res. 2007, 44, 25–30. [Google Scholar]
- Atlabachew, T.; Mamo, J. Microbiological Quality of Meat and Swabs from Contact Surface in Butcher Shops in Debre Berhan, Ethiopia. J. Food Qual. 2021, 2021, 7520882. [Google Scholar] [CrossRef]
- Kuncara, M.C.; Yuliati, F.N.; Prahesti, K.I. The Total Plate Count, Staphylococcus aureus, and PH Value of Raw Chicken Meat Sold at the Traditional Markets in Maros Regency. IOP Conf. Ser. Earth Environ. Sci. 2021, 788, 012157. [Google Scholar] [CrossRef]
- Procop, G.W.; Church, D.L.; Hall, G.S.; Janda, W.M.; Koneman, E.W.; Schreckenberger, P.C.; Woods, G.L. Koneman’ s Color Atlas and Textbook of Diagnostic Microbiology, 7th ed.; Wolters Kluwer: Alphen aan den Rijn, The Netherlands, 2017; pp. 375–515. [Google Scholar]
- Leister, C.; Hügler, M. Genome Analysis of Enterobacter asburiae and Lelliottia spp. Proliferating in Oligotrophic Drinking Water Reservoirs and Lakes. Appl. Environ. Microbiol. 2022, 88, 14. [Google Scholar] [CrossRef]
- Reitter, C.; Neuhaus, K.; Hügler, M. Draft Genome Sequences of Enterobacter spp., Lelliottia spp., and Serratia spp., Coliform Bacteria from Drinking Water Reservoirs and Lakes. Microbiol. Resour. Announc. 2021, 10, e0062221. [Google Scholar] [CrossRef]
- Yang, X.; Huang, J.; Zhang, Y.; Liu, S.; Chen, L.; Xiao, C.; Zeng, H.; Wei, X.; Gu, Q.; Li, Y.; et al. Prevalence, Abundance, Serovars and Antimicrobial Resistance of Salmonella Isolated from Retail Raw Poultry Meat in China. Sci. Tot. Environ. 2020, 713, 136385. [Google Scholar] [CrossRef]
- Dsani, E.; Afari, E.A.; Danso-Appiah, A.; Kenu, E.; Kaburi, B.B.; Egyir, B. Antimicrobial Resistance and Molecular Detection of Extended Spectrum β-Lactamase Producing Escherichia coli Isolates from Raw Meat in Greater Accra Region, Ghana. BMC Microbiol. 2020, 20, 253. [Google Scholar] [CrossRef]
- Onohuean, H.; Igere, B.E. Occurrence, Antibiotic Susceptibility and Genes Encoding Antibacterial Resistance of Salmonella spp. and Escherichia coli From Milk and Meat Sold in Markets of Bushenyi District, Uganda. Microbiol. Insights 2022, 15, 117863612210889. [Google Scholar] [CrossRef]
- Yulistiani, R.; Praseptiangga, D.; Supyani; Sudibya; Raharjo, D.; Shirakawa, T. Prevalence of Antibiotic-Resistance Enterobacteriaceae Strains Isolated from Chicken Meat at Traditional Markets in Surabaya, Indonesia. IOP Conf. Ser. Mater. Sci. Eng. 2017, 193, 012007. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Jiménez, D.; García-Meniño, I.; Fernández, J.; García, V.; Mora, A. Chicken and Turkey Meat: Consumer Exposure to Multidrug-Resistant Enterobacteriaceae Including Mcr-Carriers, Uropathogenic E. Coli and High-Risk Lineages Such as ST131. Int. J. Food Microbiol. 2020, 331, 108750. [Google Scholar] [CrossRef]
- Uzeh, R.E.; Adewumi, F.; Odumosu, B.T. Antibiotic Resistance and Plasmid Analysis of Enterobacteriaceae Isolated from Retail Meat in Lagos Nigeria. One Health Outlook 2021, 3, 10. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Document M100 ED32:2018; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Cherak, Z.; Loucif, L.; Moussi, A.; Rolain, J.M. Epidemiology of Mobile Colistin Resistance (Mcr) Genes in Aquatic Environments. J. Glob. Antimicrob. Resist. 2021, 27, 51–62. [Google Scholar] [CrossRef]
- Schauss, T.; Glaeser, S.P.; Gütschow, A.; Dott, W.; Kämpfer, P. Improved Detection of Extended Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli in Input and Output Samples of German Biogas Plants by a Selective Pre-Enrichment Procedure. PLoS ONE 2015, 10, e0119791. [Google Scholar] [CrossRef]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-Spectrum β-Lactamase-Producing and AmpC-Producing Escherichia coli from Livestock and Companion Animals, and Their Putative Impact on Public Health: A Global Perspective. Clin. Microb. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Skočková, A.; Koláčková, I.; Bogdanovičová, K.; Karpíšková, R. Characteristic and Antimicrobial Resistance in Escherichia coli from Retail Meats Purchased in the Czech Republic. Food Control 2015, 47, 401–406. [Google Scholar] [CrossRef]
- Castro, H.; Douillard, F.P.; Korkeala, H.; Lindström, M. Mobile Elements Harboring Heavy Metal and Bacitracin Resistance Genes Are Common among Listeria monocytogenes Strains Persisting on Dairy Farms. mSphere 2021, 6, 4. [Google Scholar] [CrossRef]
- Ejaz, H.; Junaid, K.; Yasmeen, H.; Naseer, A.; Alam, H.; Younas, S.; Qamar, M.U.; Abdalla, A.E.; Abosalif, K.O.A.; Ahmad, N.; et al. Multiple Antimicrobial Resistance and Heavy Metal Tolerance of Biofilm-Producing Bacteria Isolated from Dairy and Non-Dairy Food Products. Foods 2022, 11, 2728. [Google Scholar] [CrossRef]
- Mustafa, G.R.; Zhao, K.; He, X.; Chen, S.; Liu, S.; Mustafa, A.; He, L.; Yang, Y.; Yu, X.; Penttinen, P.; et al. Heavy Metal Resistance in Salmonella typhimurium and Its Association with Disinfectant and Antibiotic Resistance. Front. Microbiol. 2021, 12, 702725. [Google Scholar] [CrossRef]
- da Silva, V.L.; Caçador, N.C.; da Silva, C.; dos, S.F.; Fontes, C.O.; Garcia, G.D.; Nicoli, J.R.; Diniz, C.G. Occurrence of Multidrug-Resistant and Toxic-Metal Tolerant Enterococci in Fresh Feces from Urban Pigeons in Brazil. Microbes Environ. 2012, 27, 179. [Google Scholar] [CrossRef] [Green Version]
- Resende, J.A.; Silva, V.L.; Fontes, C.O.; Souza-Filho, J.A.; de Oliveira, T.L.R.; Coelho, C.M.; César, D.E.; Diniz, C.G. Multidrug-Resistance and Toxic Metal Tolerance of Medically Important Bacteria Isolated from an Aquaculture System. Microbes Environ. 2012, 27, 449–455. [Google Scholar] [CrossRef] [Green Version]
- al Salah, D.M.M.; Laffite, A.; Sivalingam, P.; Poté, J. Occurrence of Toxic Metals and Their Selective Pressure for Antibiotic-Resistant Clinically Relevant Bacteria and Antibiotic-Resistant Genes in River Receiving Systems under Tropical Conditions. Environ. Sci. Pol. Res. 2022, 29, 20530–20541. [Google Scholar] [CrossRef]
- Yazdankhah, S.; Skjerve, E.; Wasteson, Y. Antimicrobial resistance due to the content of potentially toxic metals in soil and fertilizing products. Microb. Ecol. Health Dis. 2018, 29, 1548248. [Google Scholar] [CrossRef]
- Lindemann, M.D.; Lu, N. Use of Chromium as an Animal Feed Supplement. The Nutritional Biochemistry of Chromium (III), 2nd ed.; Elsevier: Amsterdan, The Netherlands, 2019; pp. 79–125. [Google Scholar]
- Singh, A.; Kumar, M.; Kumar, V.; Roy, D.; Kushwaha, R.; Vaswani, S.; Kumar, A. Effects of Nickel Supplementation on Antioxidant Status, Immune Characteristics, and Energy and Lipid Metabolism in Growing Cattle. Biol. Trace Elem. Res. 2019, 190, 65–75. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, X.; Niu, Z.; Lu, D.; Sun, X.; Zhao, S.; Hou, L.; Liu, M.; Yang, Y. Antibiotic Resistance Genes (ARGs) and Their Associated Environmental Factors in the Yangtze Estuary, China: From Inlet to Outlet. Mar. Pollut. Bull 2020, 158, 111360. [Google Scholar] [CrossRef]
- Khesin, R.B.; Karasyova, E.V. Mercury-Resistant Plasmids in Bacteria from a Mercury and Antimony Deposit Area. Mol. Gen. Genet. 1984, 197, 280–285. [Google Scholar] [CrossRef]
- Yang, S.; Deng, W.; Liu, S.; Yu, X.; Mustafa, G.R.; Chen, S.; He, L.; Ao, X.; Yang, Y.; Zhou, K.; et al. Presence of Heavy Metal Resistance Genes in Escherichia coli and Salmonella Isolates and Analysis of Resistance Gene Structure in E. coli E308. J. Glob. Antimicrob. Resist. 2020, 21, 420–426. [Google Scholar] [CrossRef]
- Nies, D.H. Microbial Heavy-Metal Resistance. Appli. Microb. Biotech. 1999, 51, 730–750. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Tartor, Y.H.; Gharieb, R.M.A.; Erfan, A.M.; Khalifa, E.; Said, M.A.; Ammar, A.M.; Samir, M. Extensive Drug-Resistant Salmonella enterica Isolated from Poultry and Humans: Prevalence and Molecular Determinants Behind the Co-Resistance to Ciprofloxacin and Tigecycline. Front. Microbiol. 2021, 12, 738784. [Google Scholar] [CrossRef]
- Willers, C.; Wentzel, J.F.; du Plessis, L.H.; Gouws, C.; Hamman, J.H. Efflux as a Mechanism of Antimicrobial Drug Resistance in Clinical Relevant Microorganisms: The Role of Efflux Inhibitors. Expert. Opin. Ther. Targets 2017, 21, 23–36. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Martins, M.; Viveiros, M.; Couto, I.; Costa, S.S.; Pacheco, T.; Fanning, S.; Pagès, J.; Aamaral, L. Identification of Efflux Pump-Mediated Multidrug-Resistant Bacteria by the Ethidium Bromide-Agar Cartwheel Method. In Vivo (Brooklyn) 2011, 25, 171–178. [Google Scholar]


{kind=link}
| Source | BA 1 | EMBA 2 | MSA 3 | |
|---|---|---|---|---|
| Meat | Beef | 5.69 | 4.94 | 5.00 |
| Pork | 5.82 | 4.95 | 5.28 | |
| Chicken | 6.04 | 4.85 | 4.97 | |
| Fish | 6.14 | 5.16 | 3.77 * | |
| Total (mean count; sd 4) | 5.92; ±0.17 | 4.97; ±0.11 | 4.75; ±0.58 | |
| Antimicrobial Drugs | Bacterial Source | Total (n = 436) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Beef (n = 91) | Pork (n = 112) | Chicken (n = 91) | Fish (n = 136) | |||||||
| R | I | R | I | R | I | R | I | R | I | |
| AMP | 31.9 | 6.5 | 29.5 | 5.4 | 45.0 | 11 | 20.6 | 5.9 | 30.5 | 6.0 |
| ATM | 0.0 | 0.0 | 0.0 | 0.0 | 3.3 | 0.0 | 2.21 | 0.0 | 1.4 | 0.0 |
| AMC | 22.0 | 3.2 | 17.9 | 5.4 | 26.3 | 6.5 | 10.3 | 2.2 | 18.1 | 3.0 |
| AMI | 1.1 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.2 | 0.0 |
| GEN | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| CIP | 1.1 | 0.0 | 3.6 | 1.8 | 2.2 | 0.0 | 0.0 | 0.0 | 1.6 | 0.5 |
| LEV | 1.1 | 0.0 | 1.8 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 |
| CFZ | 46.2 | 0.0 | 33.5 | 0.0 | 46.1 | 0.0 | 34.5 | 0.0 | 39.1 | 0.0 |
| CFO | 23.1 | 4.4 | 17.0 | 2.7 | 23.0 | 3.2 | 11.0 | 2.9 | 17.7 | 3.3 |
| CAZ | 0.0 | 0.0 | 0.0 | 0.0 | 3.3 | 1.0 | 1.5 | 0.7 | 1.2 | 0.2 |
| CRO | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 0.5 | 0.0 |
| CPM | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 0.5 | 0.0 |
| CLO | 2.2 | 0.0 | 5.4 | 2.7 | 4.4 | 0.0 | 0.7 | 0.0 | 3.3 | 0.5 |
| MPM | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| SUT | 2.2 | 0.0 | 5.4 | 0.0 | 2.2 | 0.0 | 2.2 | 0.0 | 3.0 | 0.0 |
| TET | 6.6 | 0.0 | 8.9 | 1.8 | 5.5 | 2.2 | 4.4 | 2.2 | 6.3 | 1.6 |
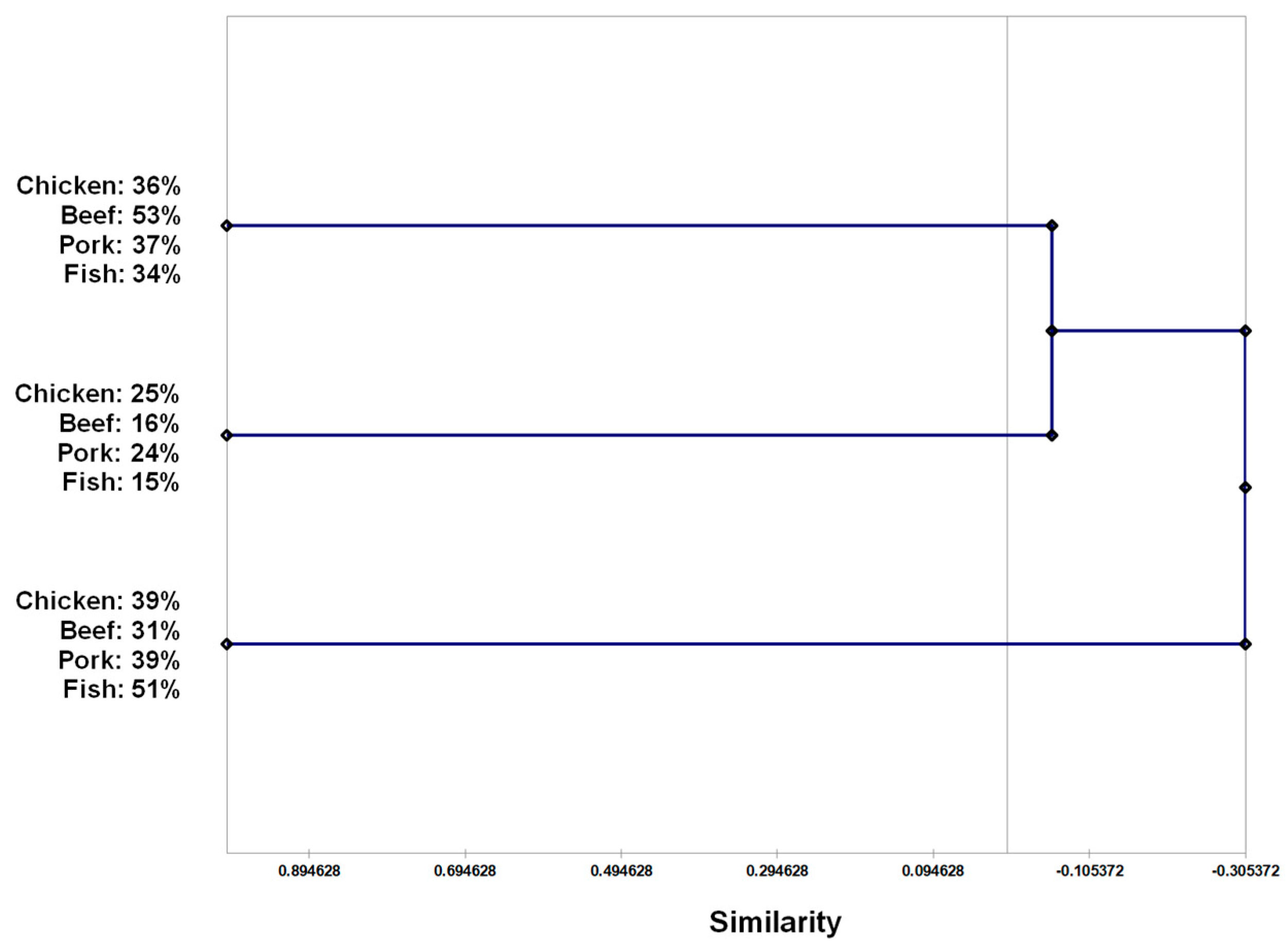
| No. of Resistance to Drugs a | % of Resistant Isolates According to Bacteria Source | Total (%) | |||
|---|---|---|---|---|---|
| Beef | Pork | Chicken | Fish | ||
| 1 | 41.8 | 33.0 | 26.4 | 30.1 | 32.3 |
| 2 | 9.9 | 9.8 | 11.0 | 6.6 | 8.1 |
| 3 | 5.5 | 10.7 | 8.8 | 4.4 | 7.9 |
| 4 | 13.2 | 9.8 | 20.9 | 5.1 | 11.4 |
| 5 | 1.1 | 2.7 | 3.3 | 2.2 | 2.3 |
| 6 | 1.1 | 1.8 | 1.1 | nd c | 0.9 |
| % of MDR b | 20.9 | 25.0 | 34.3 | 11.7 | 22.7 |
| Metal | MDR | RES | SUS | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MIC50 | MIC90 | Range | MIC50 | MIC90 | Range | MIC50 | MIC90 | Range | |
| Ni | 1024 | >1024 | 1024–>1024 | >1024 | >1024 | 512–>1024 | 1024 | >1024 | 256–>1024 |
| Zn | >1024 | >1024 | 512–>1024 | >1024 | >1024 | 128–>1024 | 1024 | >1024 | 256–>1024 |
| Hg | 32 | 256 | 2–256 | 16 | 128 | 2–256 | 32 | 64 | 8–256 |
| Cd | 512 | 1024 | 128–1024 | 256 | 512 | 128–512 | 256 | 512 | 32–1024 |
| Cr | >1024 | >1024 | >1024 | >1024 | >1024 | >1024 | >1024 | >1024 | >1024 |
| Cu | >1024 | >1024 | >1024 | >1024 | >1024 | >1024 | 1024 | 1024 | 256–1024 |
| EtBr Efflux Measure a | MDR | RES | SUS |
|---|---|---|---|
| MFC50 | 2.0 | 1.5 | 1.5 |
| MFC90 | 2.5 | 2.0 | 2.0 |
| MFC Range | 1.0–2.5 | 1.0–2.5 | 0.5–2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, R.d.C.B.; Andrade, J.; Dias, V.C.; Sequeto, J.C.; Santos, N.F.; da Silva, V.L.; Diniz, C.G. Physiological Characteristics of Putative Enterobacteria Associated with Meat and Fish Available in Southern Brazilian Retail Markets: Antimicrobial Susceptibility, Toxic Metal Tolerance and Expression of Efflux Pumps. Antibiotics 2022, 11, 1677. https://doi.org/10.3390/antibiotics11121677
Silva RdCB, Andrade J, Dias VC, Sequeto JC, Santos NF, da Silva VL, Diniz CG. Physiological Characteristics of Putative Enterobacteria Associated with Meat and Fish Available in Southern Brazilian Retail Markets: Antimicrobial Susceptibility, Toxic Metal Tolerance and Expression of Efflux Pumps. Antibiotics. 2022; 11(12):1677. https://doi.org/10.3390/antibiotics11121677
Chicago/Turabian StyleSilva, Renata da Costa Barros, Jéssica Andrade, Vanessa Cordeiro Dias, Jéssica Carla Sequeto, Nayara Felga Santos, Vânia Lúcia da Silva, and Cláudio Galuppo Diniz. 2022. "Physiological Characteristics of Putative Enterobacteria Associated with Meat and Fish Available in Southern Brazilian Retail Markets: Antimicrobial Susceptibility, Toxic Metal Tolerance and Expression of Efflux Pumps" Antibiotics 11, no. 12: 1677. https://doi.org/10.3390/antibiotics11121677