
Effect of Glycerol on Fosfomycin Activity against Escherichia coli
 , ,
, ,
Abstract
:1. Introduction
2. Results
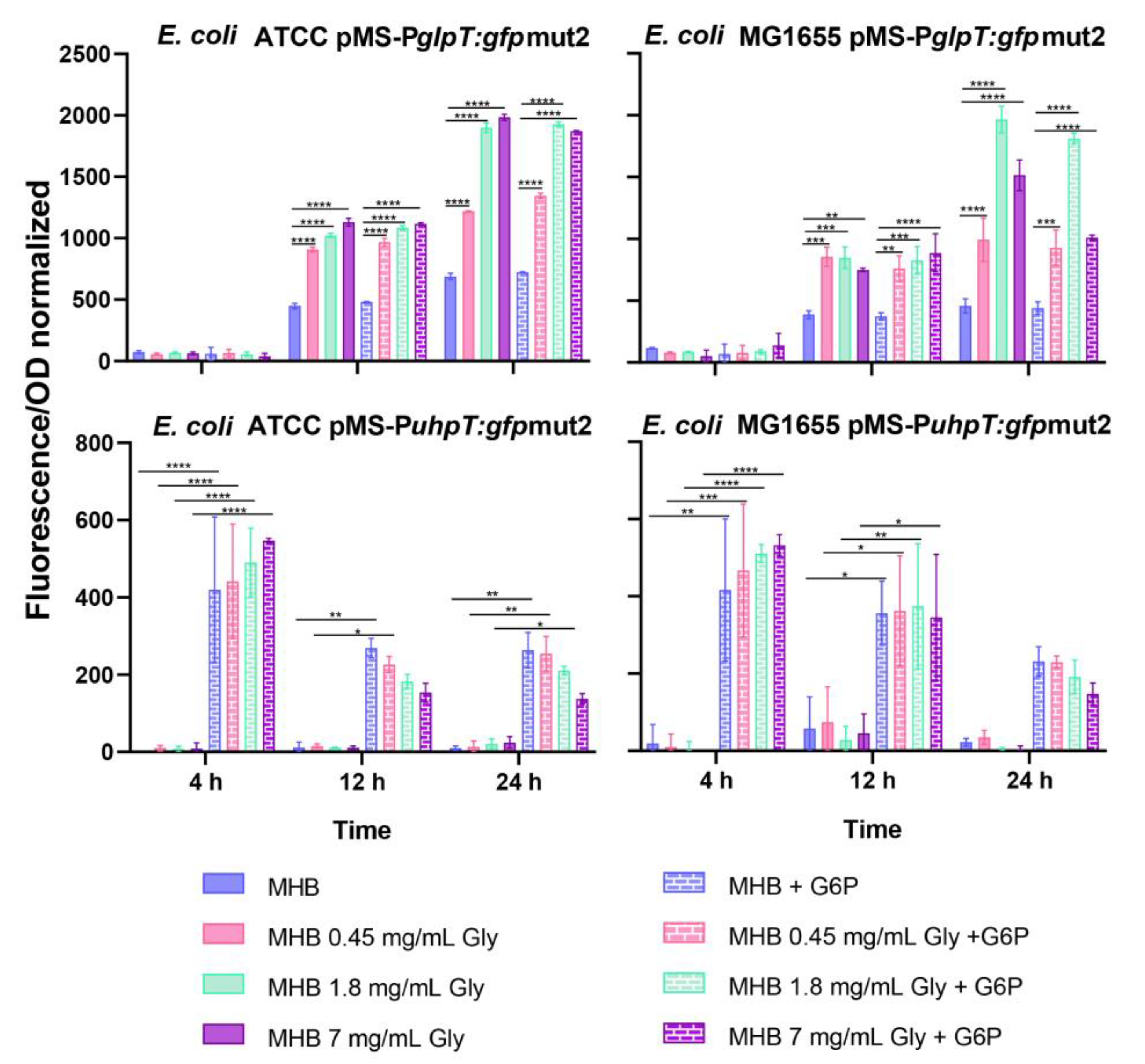
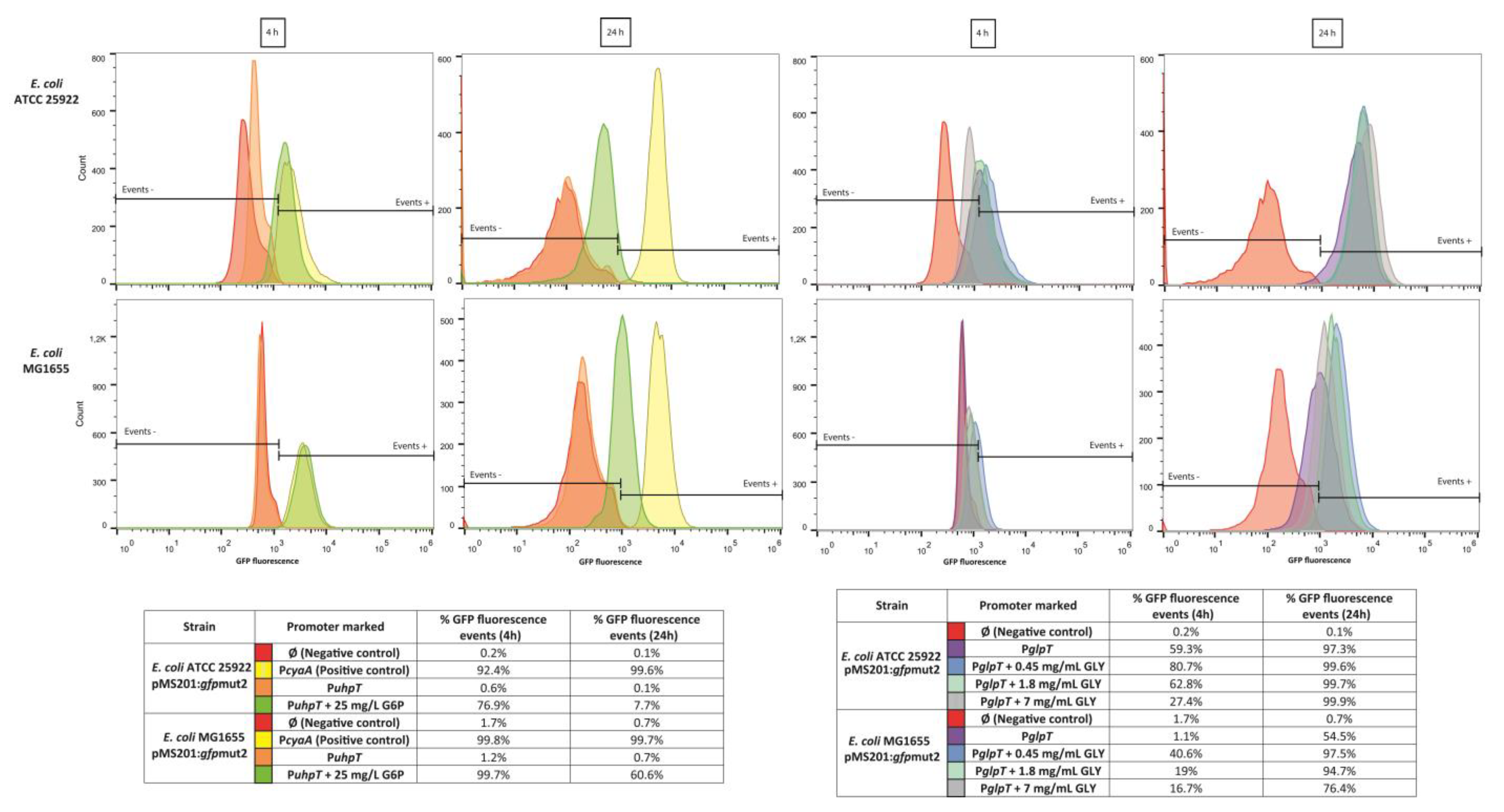
2.1. Fosfomycin Transporters Promoters Activity
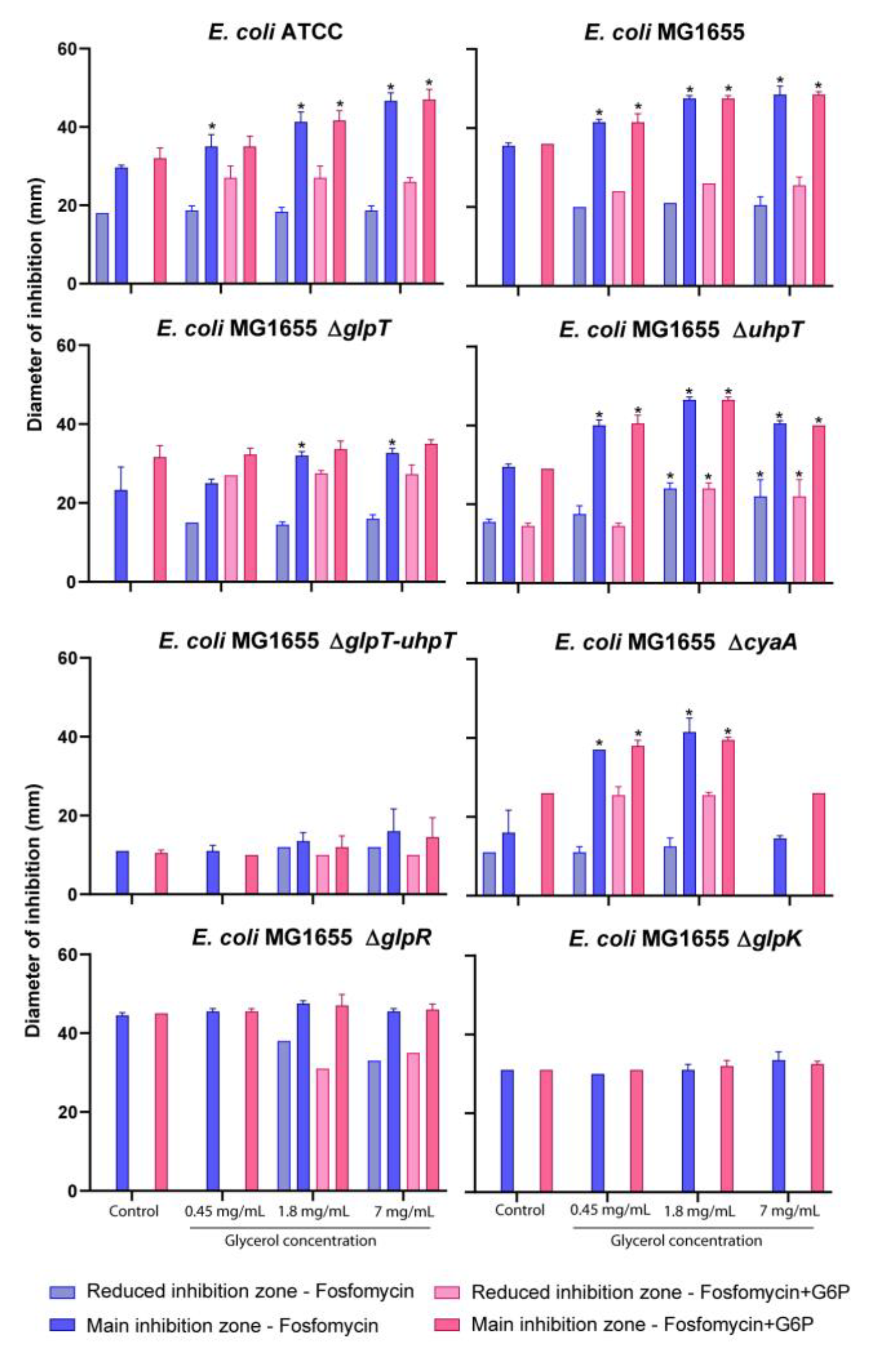
2.2. Susceptibility Testing
2.3. Fosfomycin and Glycerol Interaction Assay
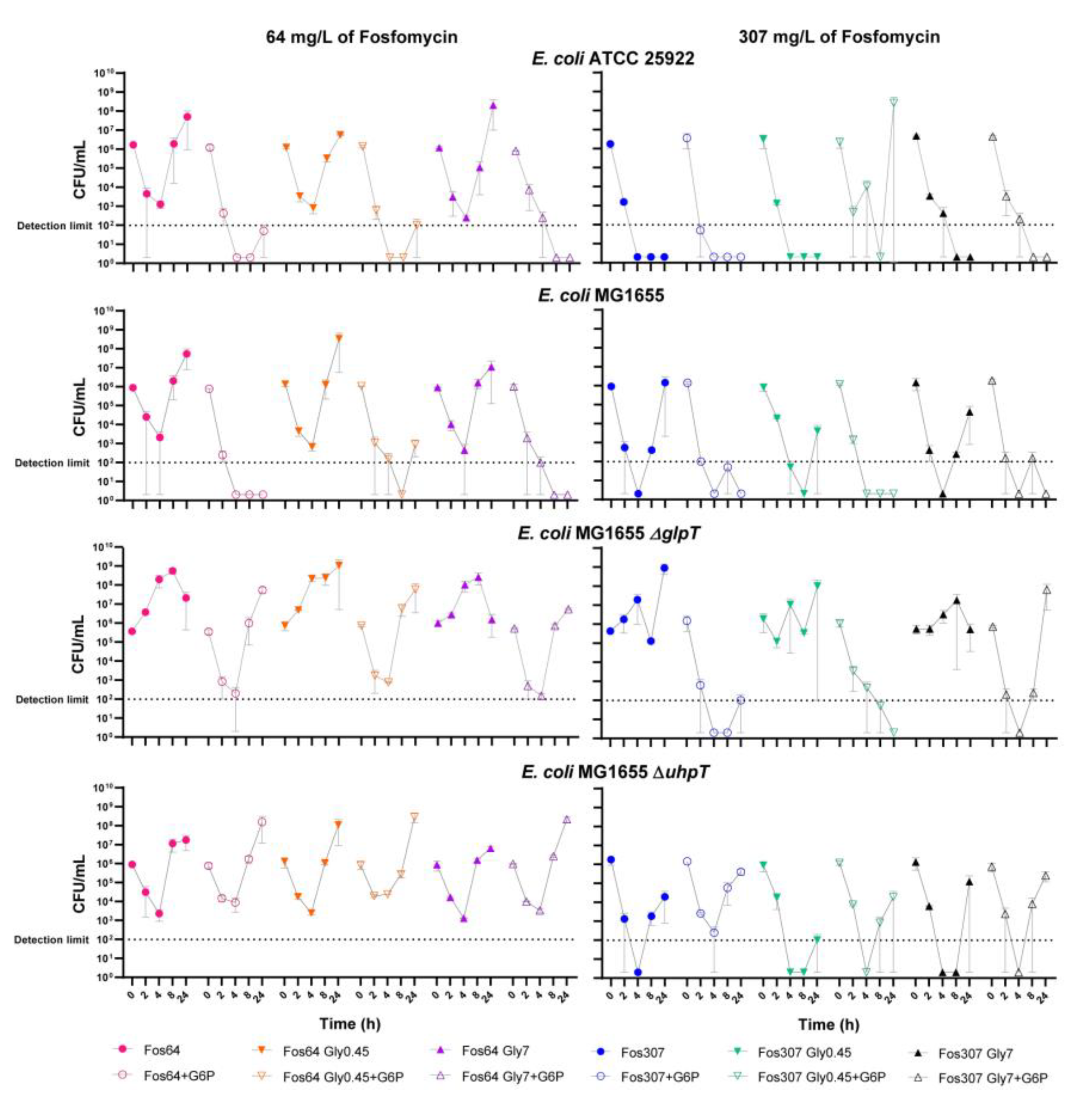
2.4. Fosfomycin and Glycerol Time-Kill Assays
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Activation Kinetic of Fosfomycin Transporters Promoters
4.3. Population Analysis of Fosfomycin Transporters Promoters Induction
4.4. Susceptibility Testing
4.5. Fosfomycin and Glycerol Interaction Assay
4.6. Fosfomycin and Glycerol Time-Kill Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballestero-Téllez, M.; Docobo-Pérez, F.; Portillo-Calderón, I.; Rodríguez-Martínez, J.M.; Racero, L.; Ramos-Guelfo, M.S.; Blázquez, J.; Rodríguez-Baño, J.; Pascual, A. Molecular insights into fosfomycin resistance in Escherichia coli. J. Antimicrob. Chemother. 2017, 72, 1303–1309. [Google Scholar] [CrossRef] [Green Version]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog Glob Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Docobo-Pérez, F.; Drusano, G.L.; Johnson, A.; Goodwin, J.; Whalley, S.; Ramos-Martín, V.; Ballestero-Tellez, M.; Rodriguez-Martinez, J.M.; Conejo, M.C.; van Guilder, M.; et al. Pharmacodynamics of fosfomycin: Insights into clinical use for antimicrobial resistance. Antimicrob. Agents Chemother. 2015, 59, 5602–5610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Maraki, S.; Karageorgopoulos, D.E.; Kastoris, A.C.; Mavromanolakis, E.; Samonis, G. Antimicrobial susceptibility of multidrug-resistant (MDR) and extensively drug-resistant (XDR) Enterobacteriaceae isolates to fosfomycin. Int. J. Antimicrob. Agents 2010, 35, 240–243. [Google Scholar] [CrossRef] [Green Version]
- Marger, M.D.; Saier, M.H.J. A major superfamily of transmembrane facilitators that catalyse uniport, symport and antiport. Trends Biochem. Sci. 1993, 18, 13–20. [Google Scholar] [CrossRef]
- Franchini, A.G.; Ihssen, J.; Egli, T. Effect of Global Regulators RpoS and Cyclic-AMP/CRP on the Catabolome and Transcriptome of Escherichia coli K12 during Carbon- and Energy-Limited Growth. PLoS ONE 2015, 10, e0133793. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, Y.; Furukawa, S.; Ogihara, H.; Yamasaki, M. Fosmidomycin resistance in adenylate cyclase deficient (cya) mutants of Escherichia coli. Biosci. Biotechnol. Biochem. 2003, 67, 2030–2033. [Google Scholar] [CrossRef] [Green Version]
- Kadner, R.J.; Shattuck-Eidens, D.M. Genetic control of the hexose phosphate transport system of Escherichia coli: Mapping of deletion and insertion mutations in the uhp region. J. Bacteriol. 1983, 155, 1052–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Gerhardt, S.G.; Larson, T.J. Action at a distance for glp repressor control of glpTQ transcription in Escherichia coli K-12. Mol. Microbiol. 1997, 24, 511–521. [Google Scholar] [CrossRef]
- Santoro, A.; Cappello, A.R.; Madeo, M.; Martello, E.; Iacopetta, D.; Dolce, V. Interaction of fosfomycin with the Glycerol 3-phosphate Transporter of Escherichia coli. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2011, 1810, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.S.; Nahata, M.C.; Hilty, M.D. Glycerol: A review of its pharmacology, pharmacokinetics, adverse reactions, and clinical use. Pharmacotherapy 1981, 1, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.C. Glycerol dissimilation and its regulation in bacteria. Annu. Rev. Microbiol. 1976, 30, 535–578. [Google Scholar] [CrossRef]
- Nikel, P.I.; Romero-Campero, F.J.; Zeidman, J.A.; Goñi-Moreno, Á.; de Lorenzo, V. The glycerol-dependent metabolic persistence of Pseudomonas putida KT2440 reflects the regulatory logic of the GlpR repressor. MBio 2015, 6, e00340-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, J.P.; Hayashi, S.; Lin, E.C. The Control of Dissimilation of Glycerol and L-α-Glycerophosphate in Escherichia coli. J. Biol. Chem. 1964, 239, 3106–3108. [Google Scholar] [CrossRef]
- Martínez-Gómez, K.; Flores, N.; Castañeda, H.M.; Martínez-Batallar, G.; Hernández-Chávez, G.; Ramírez, O.T.; Gosset, G.; Encarnación, S.; Bolivar, F. New insights into Escherichia coli metabolism: Carbon scavenging, acetate metabolism and carbon recycling responses during growth on glycerol. Microb. Cell Factories 2012, 11, 46. [Google Scholar] [CrossRef] [Green Version]
- Nikel, P.I.; Kim, J.; de Lorenzo, V. Metabolic and regulatory rearrangements underlying glycerol metabolism in Pseudomonas putidaKT2440. Environ. Microbiol. 2014, 16, 239–254. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-García, A.; Rodríguez-Rojas, A.; Guelfo, J.R.; Blázquez, J. The Glycerol-3-Phosphate Permease GlpT Is the Only Fosfomycin Transporter in Pseudomonas aeruginosa. J. Bacteriol. 2009, 191, 6968–6974. [Google Scholar] [CrossRef] [Green Version]
- Edgar, J.R.; Bell, R.M. Biosynthesis in Escherichia coli of sn-glycerol-3-phosphate, a precursor of phospholipid. Further kinetic characterization of wild type and feedback-resistant forms of the biosynthetic sn-glycerol-3-phosphate dehydrogenase. J. Biol. Chem. 1980, 255, 3492–3497. [Google Scholar] [CrossRef]
- Castañeda-García, A.; Blázquez, J.; Rodríguez-Rojas, A. Molecular mechanisms and clinical impact of acquired and intrinsic fosfomycin resistance. Antibiotics 2013, 2, 217–236. [Google Scholar] [CrossRef]
- Ballestero-Téllez, M.; Docobo-Pérez, F.; Rodríguez-Martínez, J.; Conejo, M.; Ramos-Guelfo, M.; Blázquez, J.; Rodríguez-Baño, J.; Pascual, A. Role of inoculum and mutant frequency on fosfomycin MIC discrepancies by agar dilution and broth microdilution methods in Enterobacteriaceae. Clin. Microbiol. Infect. 2017, 23, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Portillo-Calderón, I.; Ortiz-Padilla, M.; Rodríguez-Martínez, J.M.; De Gregorio-Iaria, B.; Blázquez, J.; Rodríguez-Baño, J.; Pascual, A.; Docobo-Pérez, F. Contribution of hypermutation to fosfomycin heteroresistance in Escherichia coli. J. Antimicrob. Chemother. 2020, 75, 2066–2075. [Google Scholar] [CrossRef]
- Portillo-Calderón, I.; Ortiz-Padilla, M.; de Gregorio-Iaria, B.; Merino-Bohorquez, V.; Blázquez, J.; Rodríguez-Baño, J.; Rodríguez-Martínez, J.M.; Pascual, A.; Docobo-Pérez, F. Activity of fosfomycin and amikacin against fosfomycin-heteroresistant Escherichia coli strains in a hollow-fiber infection model. Antimicrob. Agents Chemother. 2021, 65, e02213-20. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomason, L.C.; Costantino, N.; Court, D.L. E. coli genome manipulation by P1 transduction. Curr. Protoc. Mol. Biol. 2007, 79, 1.17.1–1.17.8. [Google Scholar] [CrossRef]
- Zaslaver, A.; Bren, A.; Ronen, M.; Itzkovitz, S.; Kikoin, I.; Shavit, S.; Liebermeister, W.; Surette, M.G.; Alon, U. A comprehensive library of fluorescent transcriptional reporters for Escherichia coli. Nat. Methods 2006, 3, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.; Powner, D.; Grenvik, A. Neurologic intensive care. In Anesthesia and Neurosurgery; Cottrell, J., Turndorf, H., Eds.; Mosby: Maryland Heights, MI, USA, 1980; pp. 322–360. [Google Scholar]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 21 May 2022).
- Yadav, B.; Wennerberg, K.; Aittokallio, T.; Tang, J. Searching for Drug Synergy in Complex Dose–Response Landscapes Using an Interaction Potency Model. Comput. Struct. Biotechnol. J. 2015, 13, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfausler, B. Concentrations of fosfomycin in the cerebrospinal fluid of neurointensive care patients with ventriculostomy-associated ventriculitis. J. Antimicrob. Chemother. 2004, 53, 848–852. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fosfomycin MIC (mg/L) | |||||
|---|---|---|---|---|---|
| No Glycerol | Glycerol 7 mg/L | ||||
| No G6P | G6P 25 mg/L | No G6P | G6P 25 mg/L | ||
| E. coliATCC25922 | >64 | 2 | 64 | 2 | |
| E. coliMG1655 | Wild-type | 64 | 2 | 32 | 2 |
| ΔglpT | >64 | 2 | >64 | 2 | |
| ΔuhpT | >64 | >64 | >64 | >64 | |
| ΔcyaA | >64 | 4 | >64 | 2 | |
| ΔglpR | 4 | 1 | 2 | 1 | |
| ΔglpK | >64 | 4 | 32 | 4 | |
| ΔglpT-uhpT | >64 | >64 | >64 | >64 | |
| ZIP SYNERGY SCORE | ATCC25922 | MG1655 | ||||||
|---|---|---|---|---|---|---|---|---|
| Wild-Type | ΔglpT | ΔuhpT | ΔglpT-uhpT | ΔcyaA | ΔglpR | ΔglpK | ||
| Fosfomycin + Glycerol | 16.4 ± 1.2 | 16.5 ± 1.9 | 2.5 ± 4.8 | 11.3 ± 2.9 | −12.8 ± 5.4 | −0.7 ± 6.4 | 2.7 ± 1.6 | 1.7 ± 1.9 |
| Fosfomycin + Glycerol + G6P | 6.8 ± 1.9 | 5.8 ± 2.6 | 6.9 ± 4.3 | 12.4 ± 3.6 | −21 ± 3.8 | 1.8 ± 6.6 | 6.1 ± 1.6 | 2.7 ± 3.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz-Padilla, M.; Portillo-Calderón, I.; Velázquez-Escudero, A.; Rodríguez-Baño, J.; Pascual, Á.; Rodríguez-Martínez, J.M.; Docobo-Pérez, F. Effect of Glycerol on Fosfomycin Activity against Escherichia coli. Antibiotics 2022, 11, 1612. https://doi.org/10.3390/antibiotics11111612
Ortiz-Padilla M, Portillo-Calderón I, Velázquez-Escudero A, Rodríguez-Baño J, Pascual Á, Rodríguez-Martínez JM, Docobo-Pérez F. Effect of Glycerol on Fosfomycin Activity against Escherichia coli. Antibiotics. 2022; 11(11):1612. https://doi.org/10.3390/antibiotics11111612
Chicago/Turabian StyleOrtiz-Padilla, Miriam, Inés Portillo-Calderón, Ana Velázquez-Escudero, Jesús Rodríguez-Baño, Álvaro Pascual, José Manuel Rodríguez-Martínez, and Fernando Docobo-Pérez. 2022. "Effect of Glycerol on Fosfomycin Activity against Escherichia coli" Antibiotics 11, no. 11: 1612. https://doi.org/10.3390/antibiotics11111612