


Novel Alligator Cathelicidin As-CATH8 Demonstrates Anti-Infective Activity against Clinically Relevant and Crocodylian Bacterial Pathogens
 , , , , ,
, , , , ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Bioinformatic Screen of Crocodylian Genomes Identified 18 Novel crocCATHs
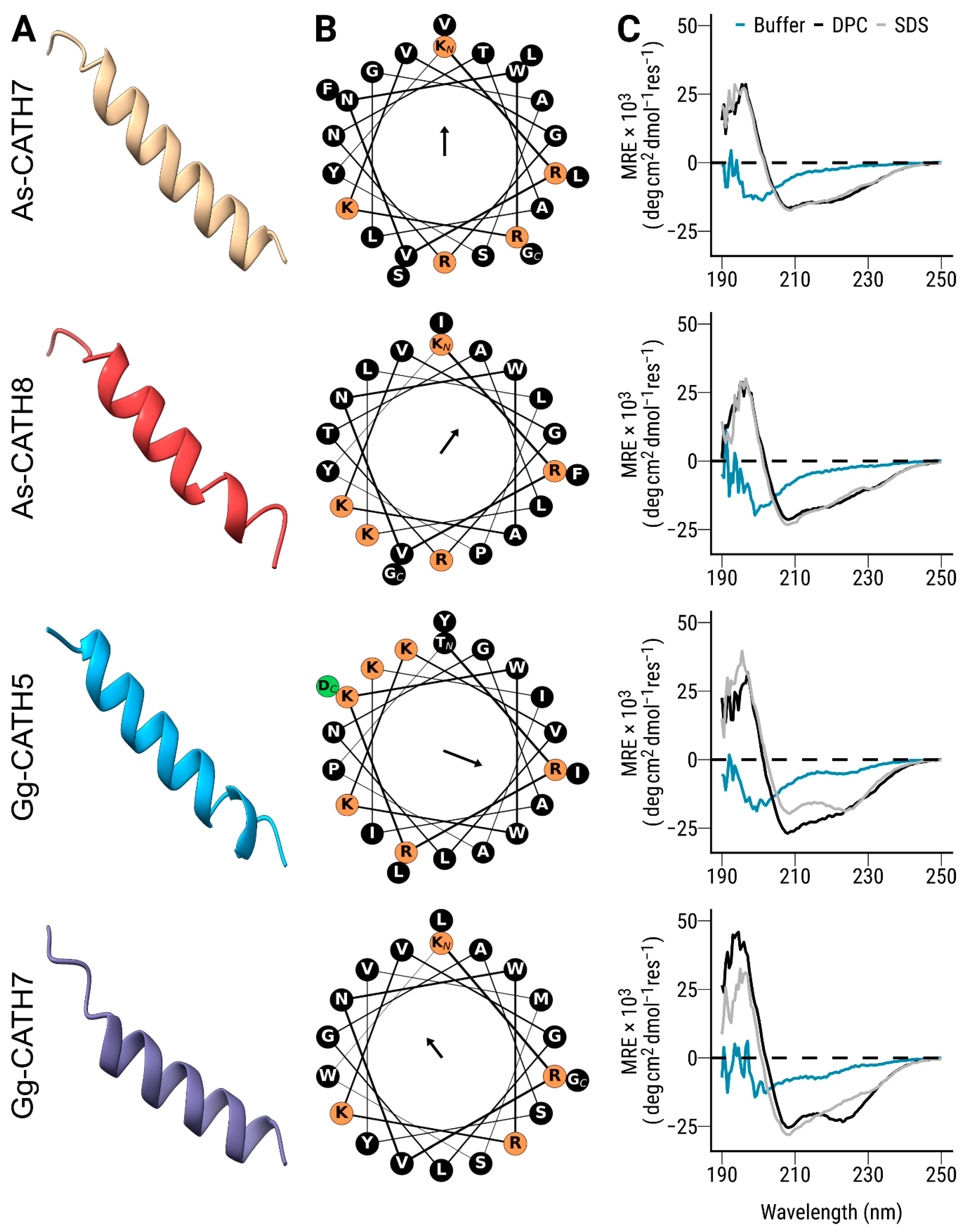
2.2. Mature crocCATHs Displayed Characteristic Properties of α-Helical Cathelicidins
2.3. As-CATH8 and Gg-CATH5 Exhibited Broad In Vitro Activities against Planktonic and Biofilm Bacteria
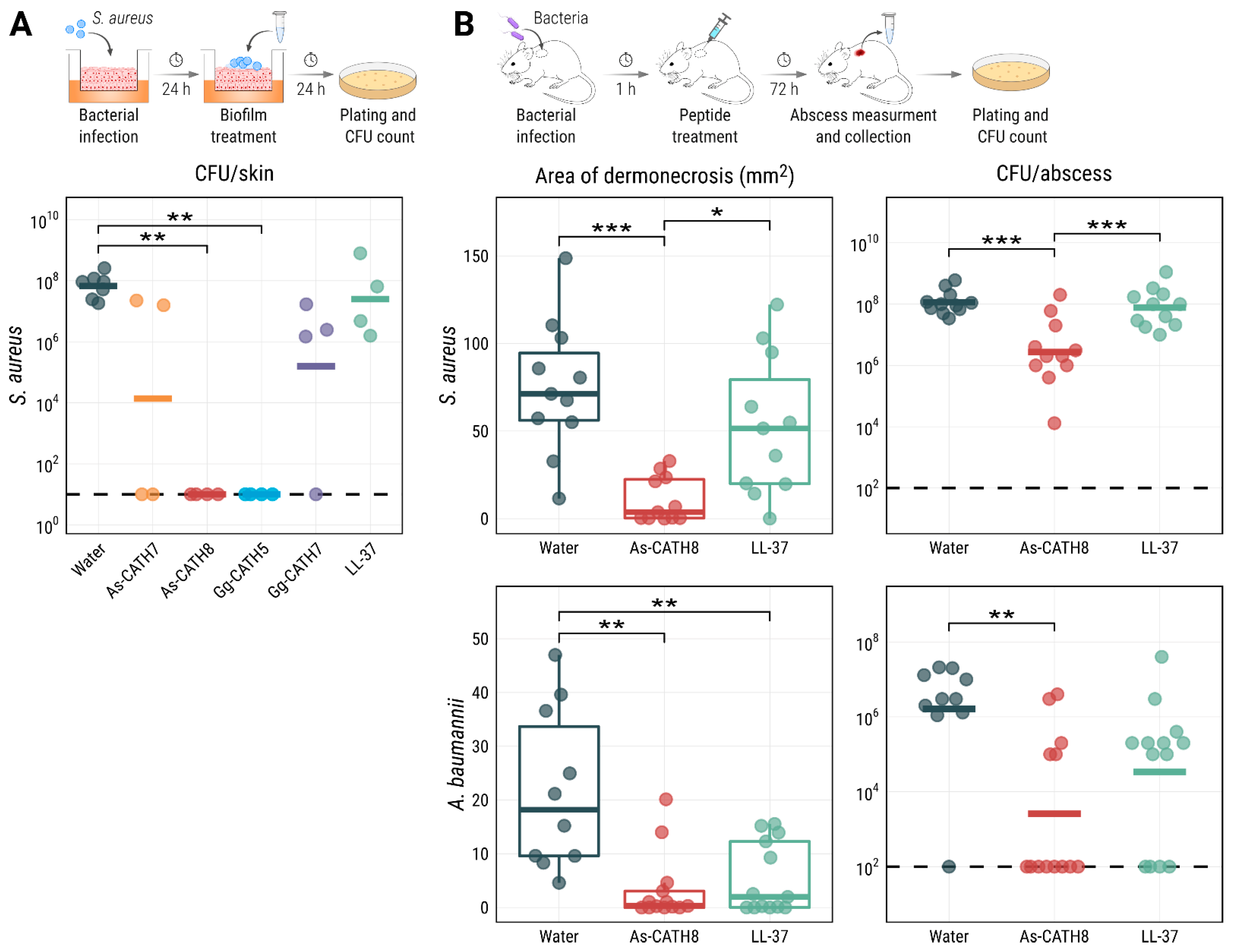
2.4. As-CATH8 and Gg-CATH5 Completely Eradicated S. aureus Biofilms in a Human Organoid Skin Model
2.5. As-CATH8 Showed a Strong Anti-Infective Effect in a Murine Abscess Model
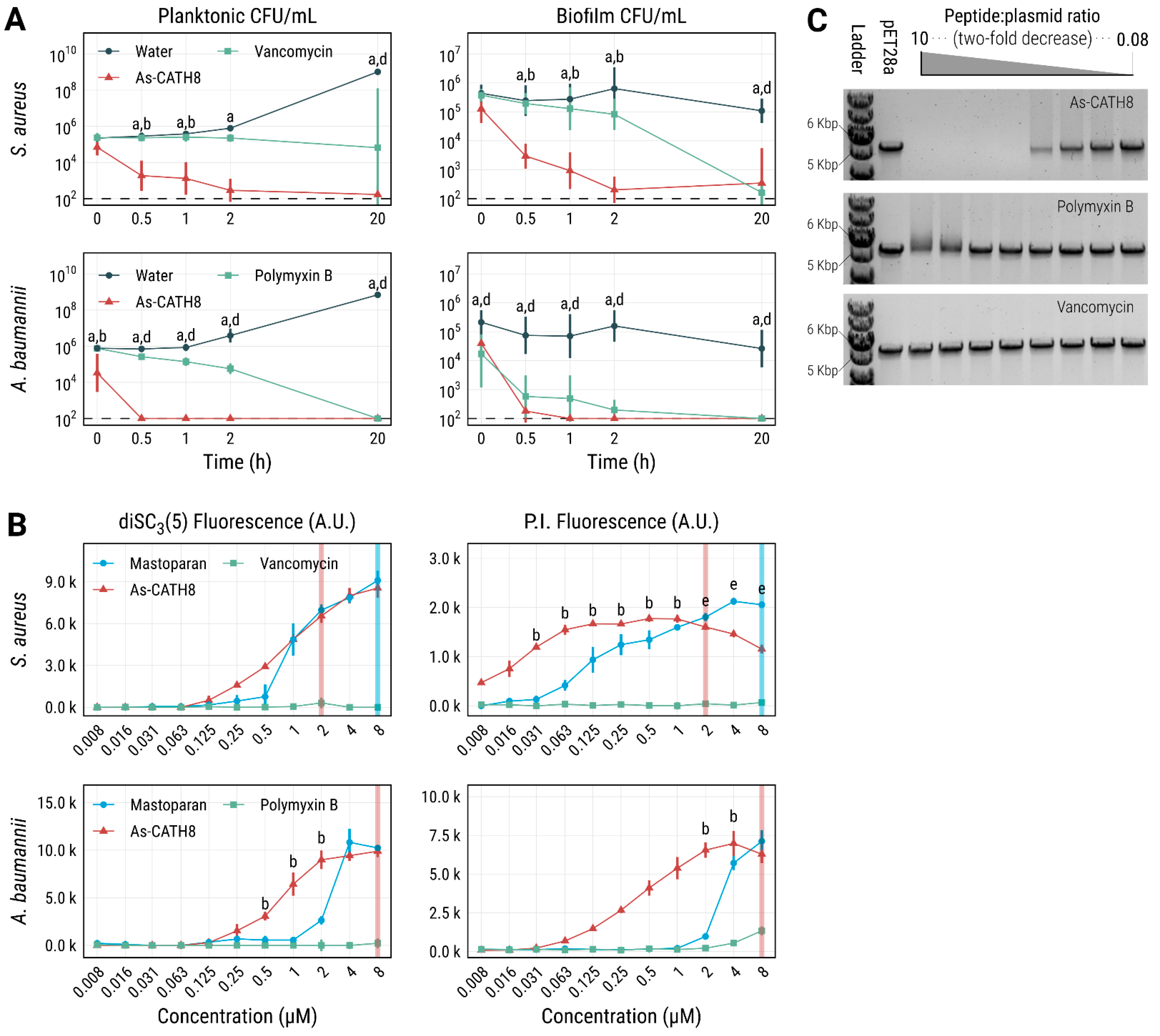
2.6. As-CATH8 Was More Bactericidal and Killed Faster Than Antibiotics
2.7. Potential Role of Interaction with Bacterial Membranes and DNA in the Antibacterial Activity of As-CATH8
3. Discussion
4. Material and Methods
4.1. Identification of Cathelicidin Sequences Using HMMs
4.2. Phylogenetic Analysis of the Cathelicidin Sequences
4.3. Prediction of Physicochemical and Structural Characteristics of Mature crocCATHs
4.4. Peptides, Reagents and Culture Media
4.5. Determination of the Secondary Structure of Synthetic Cathelicidins
4.6. Bacterial Strains and Culture Conditions
4.7. Antimicrobial Activity and Biofilm Inhibition Assays
4.8. Cell Lines and Peripheral Blood Mononuclear Cells (PBMCs)
4.9. Lactate Dehydrogenase (LDH) Release Assays
4.10. Skin Model Experiments
4.11. Bacterial Abscess Formation and Peptide Treatment
4.12. Planktonic and Biofilm-Killing Assays
4.13. Membrane Depolarization and Permeabilization Assays
4.14. Agarose Gel Electrophoretic Mobility Shift Assay
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antimicrobial Resistance Collaborators. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Mahoney, A.R.; Safaee, M.M.; Wuest, W.M.; Furst, A.L. The Silent Pandemic: Emergent Antibiotic Resistances Following the Global Response to SARS-CoV-2. iScience 2021, 24, 102304. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Dostert, M.; Trimble, M.J.; Hancock, R.E.W. Antibiofilm Peptides: Overcoming Biofilm-Related Treatment Failure. RSC Adv. 2021, 11, 2718–2728. [Google Scholar] [CrossRef]
- Cámara, M.; Green, W.; MacPhee, C.E.; Rakowska, P.D.; Raval, R.; Richardson, M.C.; Slater-Jefferies, J.; Steventon, K.; Webb, J.S. Economic Significance of Biofilms: A Multidisciplinary and Cross-Sectoral Challenge. NPJ Biofilms Microbiomes 2022, 8, 42. [Google Scholar] [CrossRef]
- Alford, M.A.; Baquir, B.; Santana, F.L.; Haney, E.F.; Hancock, R.E.W. Cathelicidin Host Defense Peptides and Inflammatory Signaling: Striking a Balance. Front. Microbiol. 2020, 11, 1902. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Alford, M.A.; Haney, E.F. Antibiofilm Activity of Host Defence Peptides: Complexity Provides Opportunities. Nat. Rev. Microbiol. 2021, 19, 786–797. [Google Scholar] [CrossRef]
- Drayton, M.; Deisinger, J.P.; Ludwig, K.C.; Raheem, N.; Müller, A.; Schneider, T.; Straus, S.K. Host Defense Peptides: Dual Antimicrobial and Immunomodulatory Action. Int. J. Mol. Sci. 2021, 22, 11172. [Google Scholar] [CrossRef]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial Host Defence Peptides: Functions and Clinical Potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Webb, G.J.W.; Messel, H. Abnormalities and Injuries in the Estuarine Crocodile Crocodylus Porosus. Wildl. Res. 1977, 4, 311–319. [Google Scholar] [CrossRef]
- Merchant, M.E.; Mills, K.; Leger, N.; Jerkins, E.; Vliet, K.A.; McDaniel, N. Comparisons of Innate Immune Activity of All Known Living Crocodylian Species. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 143, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Charruau, P.; Pérez-Flores, J.; Pérez-Juárez, J.G.; Cedeño-Vázquez, J.R.; Rosas-Carmona, R. Oral and Cloacal Microflora of Wild Crocodiles Crocodylus Acutus and C. Moreletii in the Mexican Caribbean. Dis. Aquat. Org. 2012, 98, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.S.A.; Mota, R.A.; Pinheiro Júnior, J.W.; Almeida, M.C.S.; Silva, D.R.; Ferreira, D.R.A.; Azevedo, J.C.N. Aerobic Bacterial Microflora of Broad-Snouted Caiman (Caiman latirostris) Oral Cavity and Cloaca, Originating from Parque Zoológico Arruda Câmara, Paraíba, Brazil. Braz. J. Microbiol. 2009, 40, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Lovely, C.J.; Leslie, A.J. Normal Intestinal Flora of Wild Nile Crocodiles (Crocodylus niloticus) in the Okavango Delta, Botswana. J. S. Afr. Vet. Assoc. 2008, 79, 67–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flandry, F.; Lisecki, E.J.; Domingue, G.J.; Nichols, R.L.; Greer, D.L.; Haddad, R.J. Initial Antibiotic Therapy for Alligator Bites: Characterization of the Oral Flora of Alligator Mississippiensis. South. Med. J. 1989, 82, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cai, S.; Qiao, X.; Wu, M.; Guo, Z.; Wang, R.; Kuang, Y.-Q.; Yu, H.; Wang, Y. As-Cath1-6, Novel Cathelicidins with Potent Antimicrobial and Immunomodulatory Properties from Alligator Sinensis, Play Pivotal Roles in Host Antimicrobial Immune Responses. Biochem. J. 2017, 474, 2861–2885. [Google Scholar] [CrossRef] [PubMed]
- Barksdale, S.M.; Hrifko, E.J.; van Hoek, M.L. Cathelicidin Antimicrobial Peptide from Alligator Mississippiensis Has Antibacterial Activity Against Multi-Drug Resistant Acinetobacter Baumanii and Klebsiella Pneumoniae. Dev. Comp. Immunol. 2017, 70, 135–144. [Google Scholar] [CrossRef]
- Guo, Z.; Qiao, X.; Cheng, R.; Shi, N.; Wang, A.; Feng, T.; Chen, Y.; Zhang, F.; Yu, H.; Wang, Y. As-Cath4 and 5, Two Vertebrate-Derived Natural Host Defense Peptides, Enhance the Immuno-Resistance Efficiency Against Bacterial Infections in Chinese Mitten Crab, Eriocheir Sinensis. Fish Shellfish Immunol. 2017, 71, 202–209. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, J.; Ming, Z.; Tong, H.; Wu, J.; Chen, Q.; Wang, Y.; Luo, F.; Wang, Y.; Feng, T. As-Cathelicidin4 Enhances the Immune Response and Resistance Against Aeromonas Hydrophila in Caridean Shrimp. J. Fish Dis. 2022, 45, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Hidden Markov Models. Curr. Opin. Struct. Biol. 1996, 6, 361–365. [Google Scholar] [CrossRef]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Liu, Y.; Luo, F.; Yi, Q.; Wang, Y.; Deng, L.; Dai, J.; Feng, T. Antiviral Activity of Cathelicidin 5, a Peptide from Alligator Sinensis, Against WSSV in Caridean Shrimp Exopalaemon Modestus. Fish Shellfish Immunol. 2019, 93, 82–89. [Google Scholar] [CrossRef]
- Xhindoli, D.; Pacor, S.; Benincasa, M.; Scocchi, M.; Gennaro, R.; Tossi, A. The Human Cathelicidin LL-37 Pore-Forming Antibacterial Peptide and Host-Cell Modulator. Biochim. Biophys. Acta 2016, 1858, 546–566. [Google Scholar] [CrossRef]
- Phoenix, D.A.; Harris, F. The Hydrophobic Moment and Its Use in the Classification of Amphiphilic Structures (Review). Mol. Membr. Biol. 2002, 19, 1–10. [Google Scholar] [CrossRef]
- Greenfield, N.J. Using Circular Dichroism Spectra to Estimate Protein Secondary Structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Wei, L.; Gao, J.; Zhang, S.; Wu, S.; Xie, Z.; Ling, G.; Kuang, Y.-Q.; Yang, Y.; Yu, H.; Wang, Y. Identification and Characterization of the First Cathelicidin from Sea Snakes with Potent Antimicrobial and Anti-Inflammatory Activity and Special Mechanism. J. Biol. Chem. 2015, 290, 16633–16652. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Park, K.; Kang, S.-W.; Shin, S.Y.; Hahm, K.-S.; Kim, Y. Solution Structure of a Cathelicidin-Derived Antimicrobial Peptide, CRAMP as Determined by NMR Spectroscopy. J. Pept. Res. 2002, 60, 1–9. [Google Scholar] [CrossRef]
- Gao, W.; Xing, L.; Qu, P.; Tan, T.; Yang, N.; Li, D.; Chen, H.; Feng, X. Identification of a Novel Cathelicidin Antimicrobial Peptide from Ducks and Determination of Its Functional Activity and Antibacterial Mechanism. Sci. Rep. 2015, 5, 17260. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Sidorowicz, A.; Mikosinski, J.; Krzyżanowski, M.; Orleanski, J.; Twardowska-Saucha, K.; Nykaza, A.; Dyaczynski, M.; Belz-Lagoda, B.; Dziwiszek, G.; et al. Evaluation of LL-37 in Healing of Hard-to-Heal Venous Leg Ulcers: A Multicentric Prospective Randomized Placebo-Controlled Clinical Trial. Wound Repair Regen. 2021, 29, 938–950. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.C.; Haney, E.F.; Akhoundsadegh, N.; Pletzer, D.; Trimble, M.J.; Adriaans, A.E.; Nibbering, P.H.; Hancock, R.E.W. Human Organoid Biofilm Model for Assessing Antibiofilm Activity of Novel Agents. NPJ Biofilms Microbiomes 2021, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Pletzer, D.; Mansour, S.C.; Wuerth, K.; Rahanjam, N.; Hancock, R.E.W. New Mouse Model for Chronic Infections by Gram-negative Bacteria Enabling the Study of Anti-Infective Efficacy and Host-Microbe Interactions. mBio 2017, 8, e00140-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Dhillon, P.; Yan, H.; Farmer, S.; Hancock, R.E. Interactions of Bacterial Cationic Peptide Antibiotics with Outer and Cytoplasmic Membranes of Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3317–3321. [Google Scholar] [CrossRef] [Green Version]
- Sims, P.J.; Waggoner, A.S.; Wang, C.H.; Hoffman, J.F. Studies on the Mechanism by Which Cyanine Dyes Measure Membrane Potential in Red Blood Cells and Phosphatidylcholine Vesicles. Biochemistry 1974, 13, 3315–3330. [Google Scholar] [CrossRef]
- Benfield, A.H.; Henriques, S.T. Mode-of-Action of Antimicrobial Peptides: Membrane Disruption Vs. Intracellular Mechanisms. Front. Med. Technol. 2020, 2, 610997. [Google Scholar] [CrossRef]
- dos Santos Cabrera, M.P.; Alvares, D.S.; Leite, N.B.; de Souza, B.M.; Palma, M.S.; Riske, K.A.; Neto, J.R. New Insight into the Mechanism of Action of Wasp Mastoparan Peptides: Lytic Activity and Clustering Observed with Giant Vesicles. Langmuir 2011, 27, 10805–10813. [Google Scholar] [CrossRef]
- Kristensen, K.; Ehrlich, N.; Henriksen, J.R.; Andresen, T.L. Single-Vesicle Detection and Analysis of Peptide-Induced Membrane Permeabilization. Langmuir 2015, 31, 2472–2483. [Google Scholar] [CrossRef]
- Mühlberg, E.; Umstätter, F.; Kleist, C.; Domhan, C.; Mier, W.; Uhl, P. Renaissance of Vancomycin: Approaches for Breaking Antibiotic Resistance in Multidrug-Resistant Bacteria. Can. J. Microbiol. 2020, 66, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Trimble, M.J.; Mlynárčik, P.; Kolář, M.; Hancock, R.E.W. Polymyxin: Alternative Mechanisms of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025288. [Google Scholar] [CrossRef]
- Haney, E.F.; Petersen, A.P.; Lau, C.K.; Jing, W.; Storey, D.G.; Vogel, H.J. Mechanism of Action of Puroindoline Derived Tryptophan-Rich Antimicrobial Peptides. Biochim. Biophys. Acta 2013, 1828, 1802–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santana, F.L.; Estrada, K.; Ortiz, E.; Corzo, G. Reptilian β-Defensins: Expanding the Repertoire of Known Crocodylian Peptides. Peptides 2021, 136, 170473. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Prickett, M.D.; Gutowska, W.; Kuo, R.; Belov, K.; Burt, D.W. Evolution of the Avian β-Defensin and Cathelicidin Genes. BMC Evol. Biol. 2015, 15, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelehan, C.J.; Barry-Reidy, A.; Meade, K.G.; Eckersall, P.D.; Chapwanya, A.; Narciandi, F.; Lloyd, A.T.; O’Farrelly, C. Characterisation and Expression Profile of the Bovine Cathelicidin Gene Repertoire in Mammary Tissue. BMC Genom. 2014, 15, 128. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Maier, E.; Benz, R.; Hancock, R.E. Mechanism of Interaction of Different Classes of Cationic Antimicrobial Peptides with Planar Bilayers and with the Cytoplasmic Membrane of Escherichia coli. Biochemistry 1999, 38, 7235–7242. [Google Scholar] [CrossRef]
- Hsu, C.-H.; Chen, C.; Jou, M.-L.; Lee, A.Y.-L.; Lin, Y.-C.; Yu, Y.-P.; Huang, W.-T.; Wu, S.-H. Structural and DNA-binding Studies on the Bovine Antimicrobial Peptide, Indolicidin: Evidence for Multiple Conformations Involved in Binding to Membranes and DNA. Nucleic Acids Res. 2005, 33, 4053–4064. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Shan, B.; Qi, J.; Ma, Y. Systemic Responses of Multidrug-Resistant Pseudomonas Aeruginosa and Acinetobacter Baumannii Following Exposure to the Antimicrobial Peptide Cathelicidin-BF Imply Multiple Intracellular Targets. Front. Cell. Infect. Microbiol. 2017, 7, 466. [Google Scholar] [CrossRef]
- Limoli, D.H.; Rockel, A.B.; Host, K.M.; Jha, A.; Kopp, B.T.; Hollis, T.; Wozniak, D.J. Cationic Antimicrobial Peptides Promote Microbial Mutagenesis and Pathoadaptation in Chronic Infections. PLoS Pathog. 2014, 10, e1004083. [Google Scholar] [CrossRef] [Green Version]
- Scheenstra, M.R.; van den Belt, M.; Tjeerdsma-van Bokhoven, J.L.M.; Schneider, V.A.F.; Ordonez, S.R.; van Dijk, A.; Veldhuizen, E.J.A.; Haagsman, H.P. Cathelicidins PMAP-36, LL-37 and CATH-2 Are Similar Peptides with Different Modes of Action. Sci. Rep. 2019, 9, 4780. [Google Scholar] [CrossRef] [Green Version]
- Velkov, T.; Thompson, P.E.; Nation, R.L.; Li, J. Structure-Activity Relationships of Polymyxin Antibiotics. J. Med. Chem. 2010, 53, 1898–1916. [Google Scholar] [CrossRef]
- Jia, Z.; O’Mara, M.L.; Zuegg, J.; Cooper, M.A.; Mark, A.E. The Effect of Environment on the Recognition and Binding of Vancomycin to Native and Resistant Forms of Lipid II. Biophys. J. 2011, 101, 2684–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas, I.; Ibarra, M.A.; Santana, F.L.; Villegas, E.; Hancock, R.E.W.; Corzo, G. In Vitro and in Vivo Antibiotic Capacity of Two Host Defense Peptides. Antimicrob. Agents Chemother. 2020, 64, e00145-20. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Cai, S.; Gao, J.; Qiao, X.; Yang, H.; Wang, Y.; Yu, H. Roles of Polymorphic Cathelicidins in Innate Immunity of Soft-Shell Turtle, Pelodiscus Sinensis. Dev. Comp. Immunol. 2019, 92, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Qiao, X.; Feng, L.; Shi, N.; Wang, H.; Yang, H.; Guo, Z.; Wang, M.; Chen, Y.; Wang, Y.; et al. Python Cathelicidin CATHPb1 Protects Against Multidrug-Resistant Staphylococcal Infections by Antimicrobial-Immunomodulatory Duality. J. Med. Chem. 2018, 61, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.-H.; Pan, S.-K.; Hu, L.; Zhu, Y.; Xu, P.-W.; Xia, J.-Q.; Chen, H.; He, G.-Y.; He, J.; Ni, X.-W.; et al. Genome Analysis and Signature Discovery for Diving and Sensory Properties of the Endangered Chinese Alligator. Cell Res. 2013, 23, 1091–1105. [Google Scholar] [CrossRef] [Green Version]
- St John, J.A.; Braun, E.L.; Isberg, S.R.; Miles, L.G.; Chong, A.Y.; Gongora, J.; Dalzell, P.; Moran, C.; Bed’hom, B.; Abzhanov, A.; et al. Sequencing Three Crocodilian Genomes to Illuminate the Evolution of Archosaurs and Amniotes. Genome Biol. 2012, 13, 415. [Google Scholar] [CrossRef]
- Köster, J.; Rahmann, S. Snakemake—A Scalable Bioinformatics Workflow Engine. Bioinformatics 2012, 28, 2520–2522. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [Green Version]
- Slater, G.S.C.; Birney, E. Automated Generation of Heuristics for Biological Sequence Comparison. BMC Bioinform. 2005, 6, 31. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Gan, T.-X.; Liu, X.-D.; Jin, Y.; Lee, W.-H.; Shen, J.-H.; Zhang, Y. Identification and Characterization of Novel Reptile Cathelicidins from Elapid Snakes. Peptides 2008, 29, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Cai, Y.; Bommineni, Y.R.; Fernando, S.C.; Prakash, O.; Gilliland, S.E.; Zhang, G. Identification and Functional Characterization of Three Chicken Cathelicidins with Potent Antimicrobial Activity. J. Biol. Chem. 2006, 281, 2858–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.J.; Chen, J.; Huang, Z.A.; Shi, Y.H.; Lv, J.N. Identification and Characterization of a Novel Cathelicidin from Ayu, Plecoglossus Altivelis. Fish Shellfish Immunol. 2011, 31, 52–57. [Google Scholar] [CrossRef]
- Hao, X.; Yang, H.; Wei, L.; Yang, S.; Zhu, W.; Ma, D.; Yu, H.; Lai, R. Amphibian Cathelicidin Fills the Evolutionary Gap of Cathelicidin in Vertebrate. Amino Acids 2012, 43, 677–685. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of Dna and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-G.; Lam, T.T.-Y.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R Package for Phylogenetic Tree Input and Output with Richly Annotated and Associated Data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef]
- Yu, G. Using Ggtree to Visualize Data on Tree-Like Structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- RStudio Team. Rstudio: Integrated Development Environment for R; RStudio, PBC.: Boston, MA, USA, 2022. [Google Scholar]
- Osorio, D.; Rondon-Villarreal, P.; Torres, R. Peptides: A Package for Data Mining of Antimicrobial Peptides. R J. 2015, 7, 4–14. [Google Scholar] [CrossRef]
- Bjellqvist, B.; Hughes, G.J.; Pasquali, C.; Paquet, N.; Ravier, F.; Sanchez, J.C.; Frutiger, S.; Hochstrasser, D. The Focusing Positions of Polypeptides in Immobilized pH Gradients Can Be Predicted from Their Amino Acid Sequences. Electrophoresis 1993, 14, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A Simple Method for Displaying the Hydropathic Character of a Protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.M. (Ed.) The Proteomics Protocols Handbook, 3rd ed.; Humana Press: Totowa, NJ, USA, 2005; ISBN 978-1-58829-343-5. [Google Scholar]
- Müller, A.T.; Gabernet, G.; Hiss, J.A.; Schneider, G. modlAMP: Python for Antimicrobial Peptides. Bioinformatics 2017, 33, 2753–2755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Ovchinnikov, S.; Steinegger, M. ColabFold—Making Protein Folding Accessible to All. bioRxv 2021. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure Visualization for Researchers, Educators, and Developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Haney, E.F.; Wu, B.C.; Lee, K.; Hilchie, A.L.; Hancock, R.E.W. Aggregation and Its Influence on the Immunomodulatory Activity of Synthetic Innate Defense Regulator Peptides. Cell Chem. Biol. 2017, 24, 969–980.e4. [Google Scholar] [CrossRef] [Green Version]
- Hilchie, A.L.; Sharon, A.J.; Haney, E.F.; Hoskin, D.W.; Bally, M.B.; Franco, O.L.; Corcoran, J.A.; Hancock, R.E.W. Mastoparan Is a Membranolytic Anti-Cancer Peptide That Works Synergistically with Gemcitabine in a Mouse Model of Mammary Carcinoma. Biochim. Biophys. Acta 2016, 1858, 3195–3204. [Google Scholar] [CrossRef]
- Raheem, N.; Kumar, P.; Lee, E.; Cheng, J.T.J.; Hancock, R.E.W.; Straus, S.K. Insights into the Mechanism of Action of Two Analogues of Aurein 2.2. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183262. [Google Scholar] [CrossRef]
- Chase-Topping, M.; Gally, D.; Low, C.; Matthews, L.; Woolhouse, M. Super-Shedding and the Link Between Human Infection and Livestock Carriage of Escherichia coli O157. Nat. Rev. Microbiol. 2008, 6, 904–912. [Google Scholar] [CrossRef]
- Mansour, S.C.; Pletzer, D.; de la Fuente-Núñez, C.; Kim, P.; Cheung, G.Y.C.; Joo, H.-S.; Otto, M.; Hancock, R.E.W. Bacterial Abscess Formation Is Controlled by the Stringent Stress Response and Can Be Targeted Therapeutically. EBioMedicine 2016, 12, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pletzer, D.; Mansour, S.C.; Hancock, R.E.W. Synergy Between Conventional Antibiotics and Anti-Biofilm Peptides in a Murine, Sub-Cutaneous Abscess Model Caused by Recalcitrant ESKAPE Pathogens. PLoS Pathog. 2018, 14, e1007084. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Trimble, M.J.; Hancock, R.E.W. Microtiter Plate Assays to Assess Antibiofilm Activity Against Bacteria. Nat. Protoc. 2021, 16, 2615–2632. [Google Scholar] [CrossRef]
- Santana, F.L.; Arenas, I.; Haney, E.F.; Estrada, K.; Hancock, R.E.W.; Corzo, G. Identification of a Crocodylian -Defensin Variant from Alligator Mississippiensis with Antimicrobial and Antibiofilm Activity. Peptides 2021, 141, 170549. [Google Scholar] [CrossRef]
- Boix-Lemonche, G.; Lekka, M.; Skerlavaj, B. A Rapid Fluorescence-Based Microplate Assay to Investigate the Interaction of Membrane Active Antimicrobial Peptides with Whole Gram-Positive Bacteria. Antibiotics 2020, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A. Practical Statistics in R for Comparing Groups: Numerical Variables; Practical statistics in R series; Independently Published, 2019; ISBN 978-1-71233-088-3. [Google Scholar]
- Tankrathok, A.; Punpad, A.; Kongchaiyapoom, M.; Sosiangdi, S.; Jangpromma, N.; Daduang, S.; Klaynongsruang, S. Identification of the First Crocodylus Siamensis Cathelicidin Gene and Rn15 Peptide Derived from Cathelin Domain Exhibiting Antibacterial Activity. Biotechnol. Appl. Biochem. 2019, 66, 142–152. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Length | MW | Charge | HI | HM |
|---|---|---|---|---|---|---|
| As-CATH7 | KRVNWRKVGRNTALGASYVLSFLG | 24 | 2693 | 4.76 | −0.15 | 0.25 |
| As-CATH8 | KRVNWAKVGRTALKLLPYIFG | 21 | 2431 | 4.76 | 0.06 | 0.29 |
| Gg-CATH5 | TRRKWWKKVLNGAIKIAPYILD | 22 | 2670 | 4.76 | −0.39 | 0.40 |
| Gg-CATH7 | KRVNWRKVGLGASYVMSWLG | 20 | 2308 | 3.76 | −0.11 | 0.23 |
| LL-37 | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | 37 | 4493 | 5.76 | −0.72 | 0.56 |
| Bacteria | MIC in μM | |||||
|---|---|---|---|---|---|---|
| As-CATH7 | As-CATH8 | Gg-CATH5 | Gg-CATH7 | Polymyxin B | Vancomycin | |
| E. cloacae | 0.5 | 0.5 | 0.5 | 2 | 0.5 | n.d. |
| S. aureus | 4 | 0.5 | 1 | 16 | n.d. | 0.5 |
| K. pneumoniae | 1 | 0.5 | 0.5 | 4 | 0.5 | n.d. |
| A. baumannii | 0.25 | 0.25 | 0.5 | 1 | 0.5 | n.d. |
| P. aeruginosa | 4 | 1 | 1 | 8 | 0.5 | n.d. |
| E. faecium | >64 | >64 | >64 | 64 | n.d. | >64 |
| E. coli | 2 | 1 | 4 | 4 | 0.5 | n.d. |
| S. Typhimurium | 2 | 1 | 0.5 | 4 | 1 | n.d. |
| P. vulgaris | >64 | 4 | 8 | >64 | >64 | n.d. |
| Bacteria | MBIC95 in μM | |||
|---|---|---|---|---|
| As-CATH7 | As-CATH8 | Gg-CATH5 | Gg-CATH7 | |
| E. cloacae | 32 | 4 | 4 | 16 |
| S. aureus | 4 | 1 | 1 | 4 |
| A. baumannii | 1 | 0.5 | 0.5 | 1 |
| P. aeruginosa | >64 | 64 | 32 | >64 |
| E. coli | 32 | 1 | 32 | 32 |
| S. Typhimurium | 1 | 1 | 1 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, F.L.; Estrada, K.; Alford, M.A.; Wu, B.C.; Dostert, M.; Pedraz, L.; Akhoundsadegh, N.; Kalsi, P.; Haney, E.F.; Straus, S.K.; et al. Novel Alligator Cathelicidin As-CATH8 Demonstrates Anti-Infective Activity against Clinically Relevant and Crocodylian Bacterial Pathogens. Antibiotics 2022, 11, 1603. https://doi.org/10.3390/antibiotics11111603
Santana FL, Estrada K, Alford MA, Wu BC, Dostert M, Pedraz L, Akhoundsadegh N, Kalsi P, Haney EF, Straus SK, et al. Novel Alligator Cathelicidin As-CATH8 Demonstrates Anti-Infective Activity against Clinically Relevant and Crocodylian Bacterial Pathogens. Antibiotics. 2022; 11(11):1603. https://doi.org/10.3390/antibiotics11111603
Chicago/Turabian StyleSantana, Felix L., Karel Estrada, Morgan A. Alford, Bing C. Wu, Melanie Dostert, Lucas Pedraz, Noushin Akhoundsadegh, Pavneet Kalsi, Evan F. Haney, Suzana K. Straus, and et al. 2022. "Novel Alligator Cathelicidin As-CATH8 Demonstrates Anti-Infective Activity against Clinically Relevant and Crocodylian Bacterial Pathogens" Antibiotics 11, no. 11: 1603. https://doi.org/10.3390/antibiotics11111603