
Antifungal Activity of a Library of Aminothioxanthones
 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
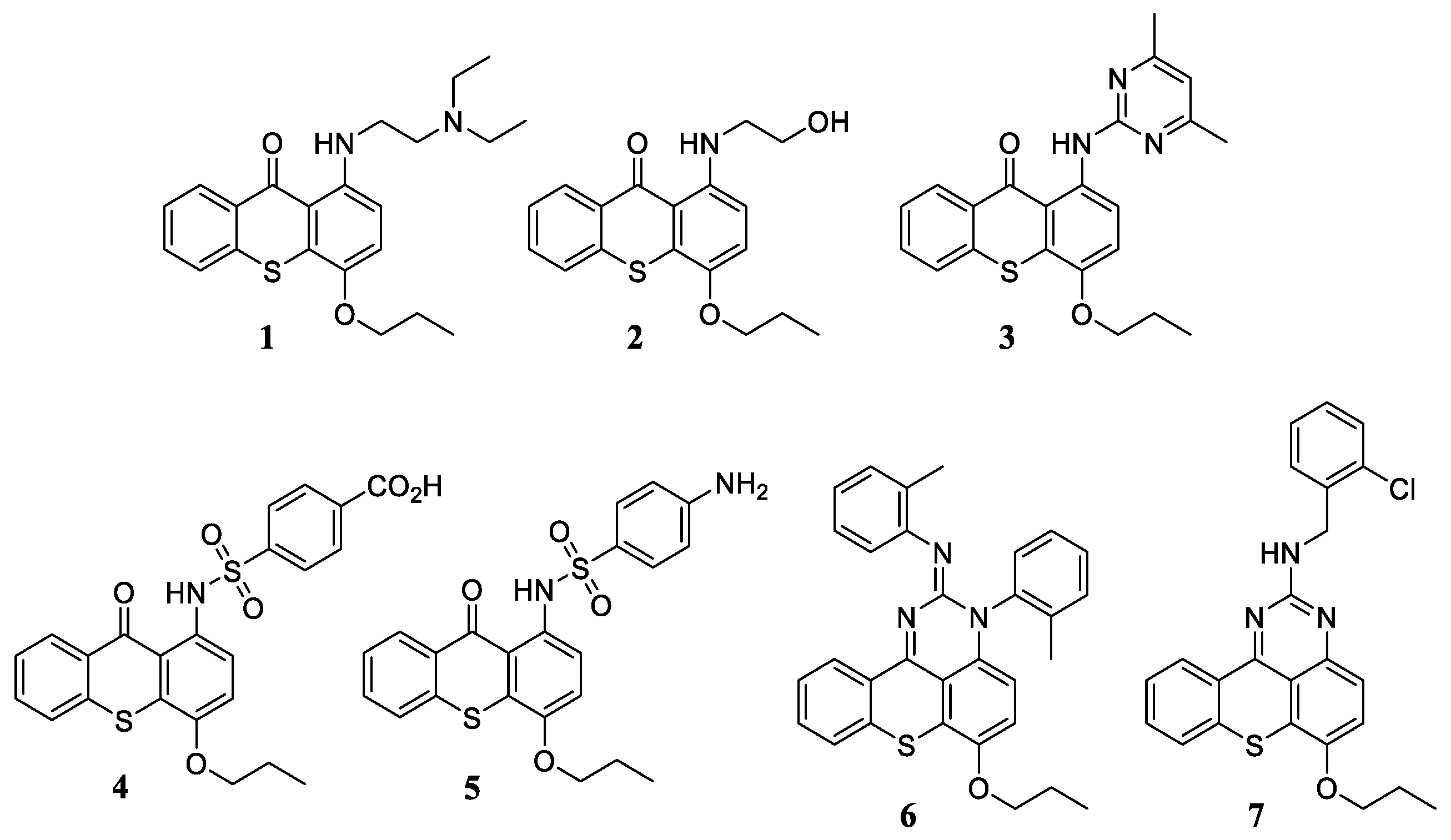
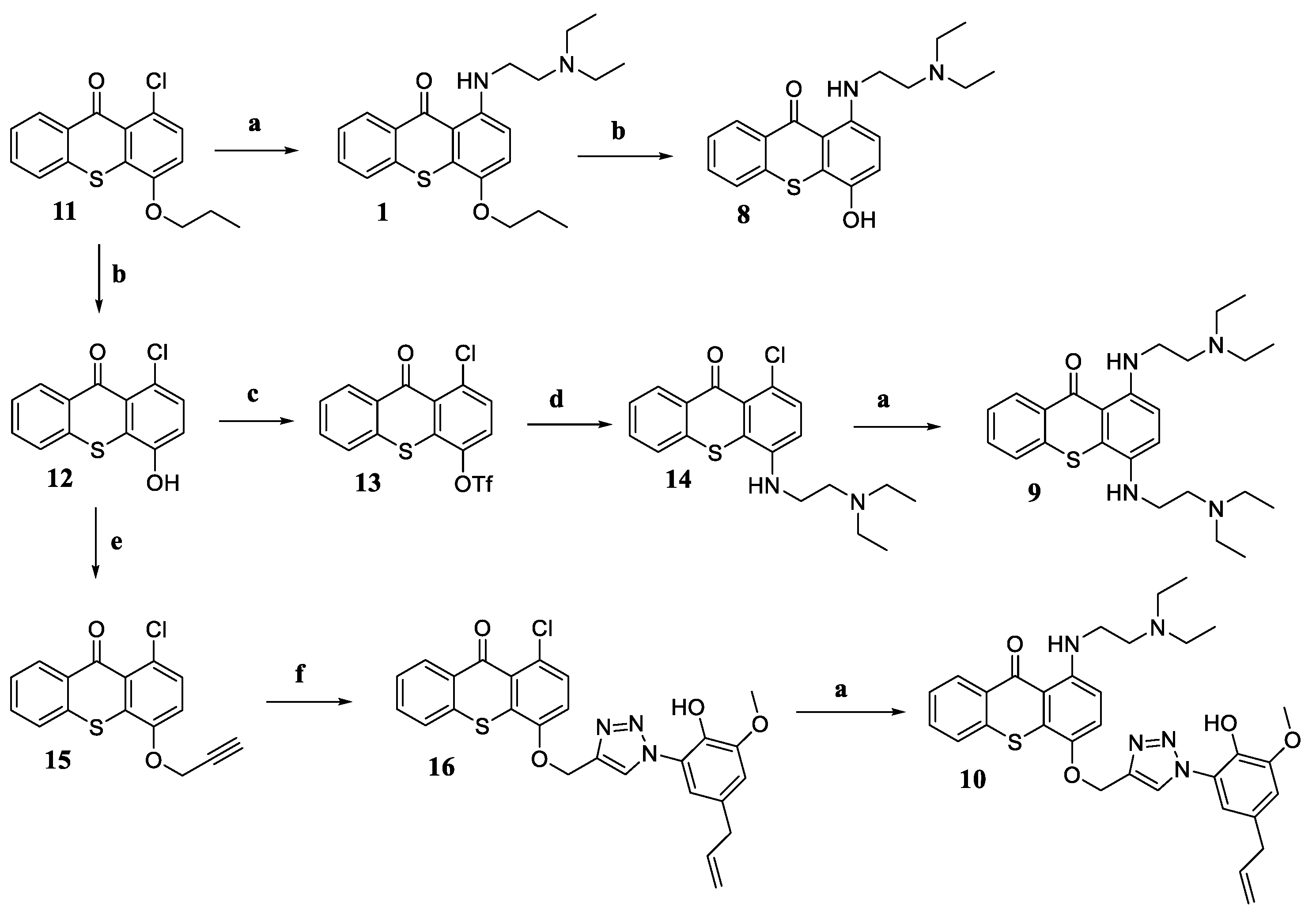
2.1. Chemistry
2.2. Antifungal Activity
2.3. Study of C. albicans Phenotypic Virulence Factors
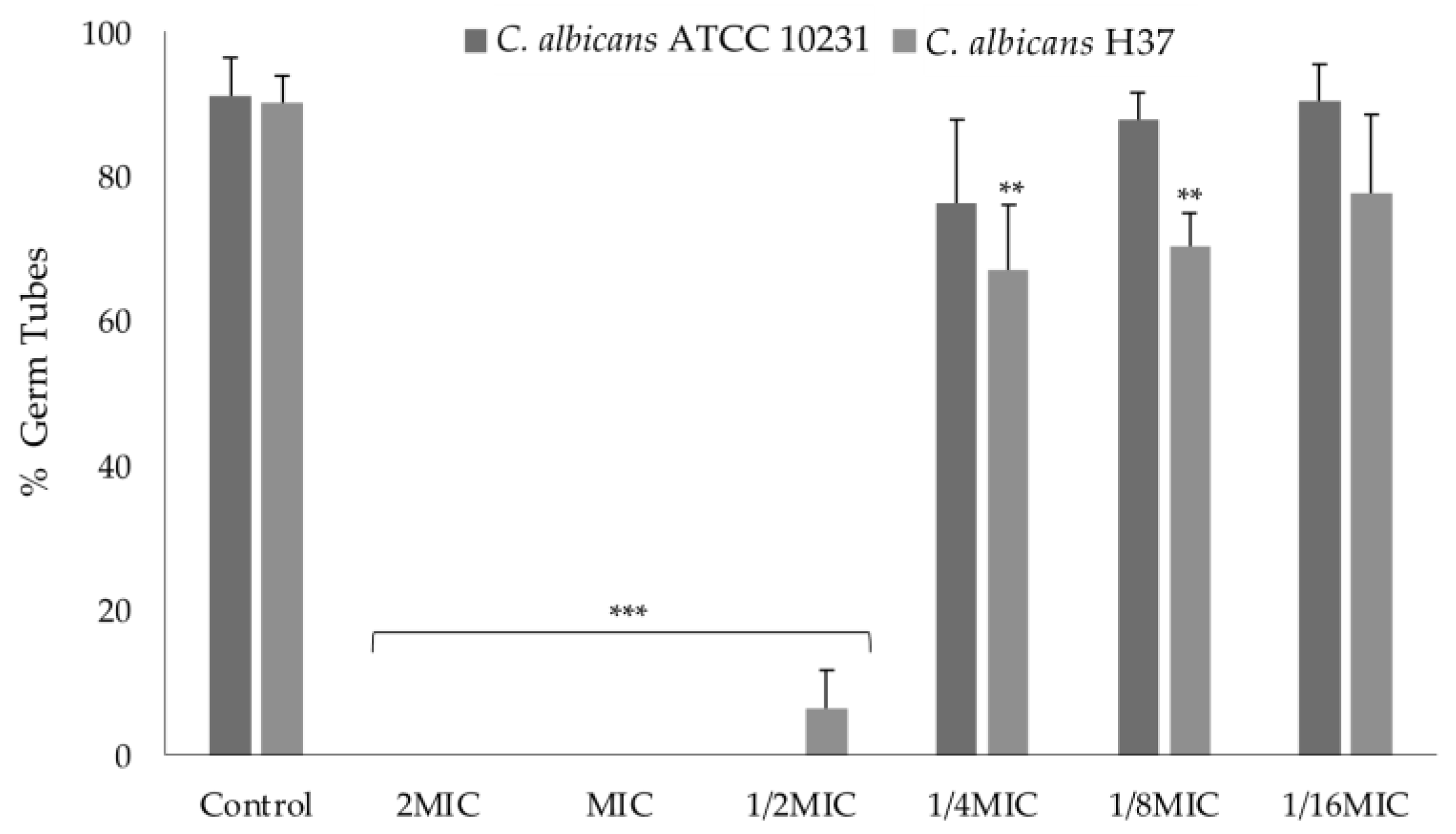
2.3.1. Effect on Germ Tube Formation
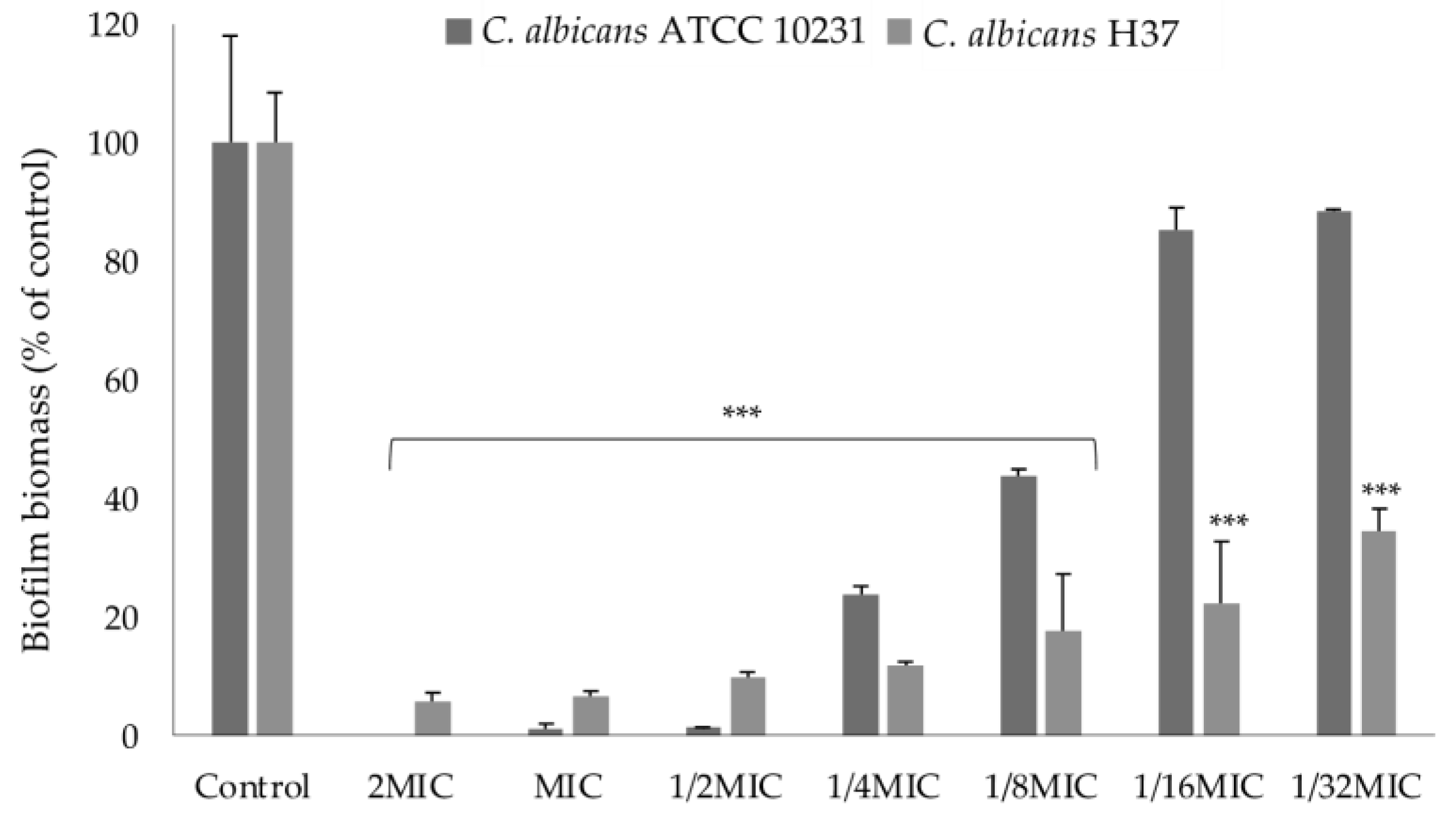
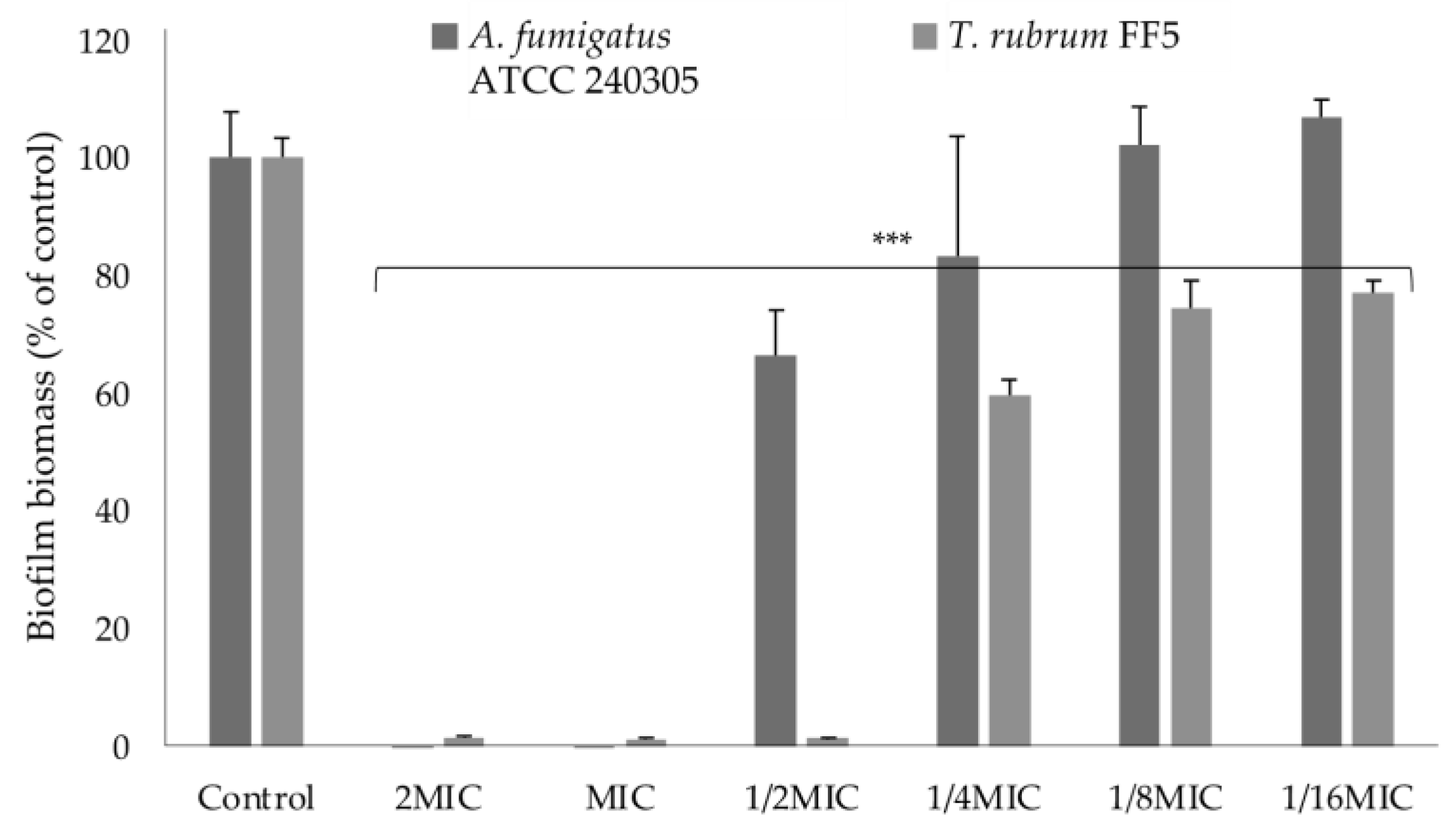
2.3.2. Effect on Biofilm Formation
2.4. Study of Mechanism of Action
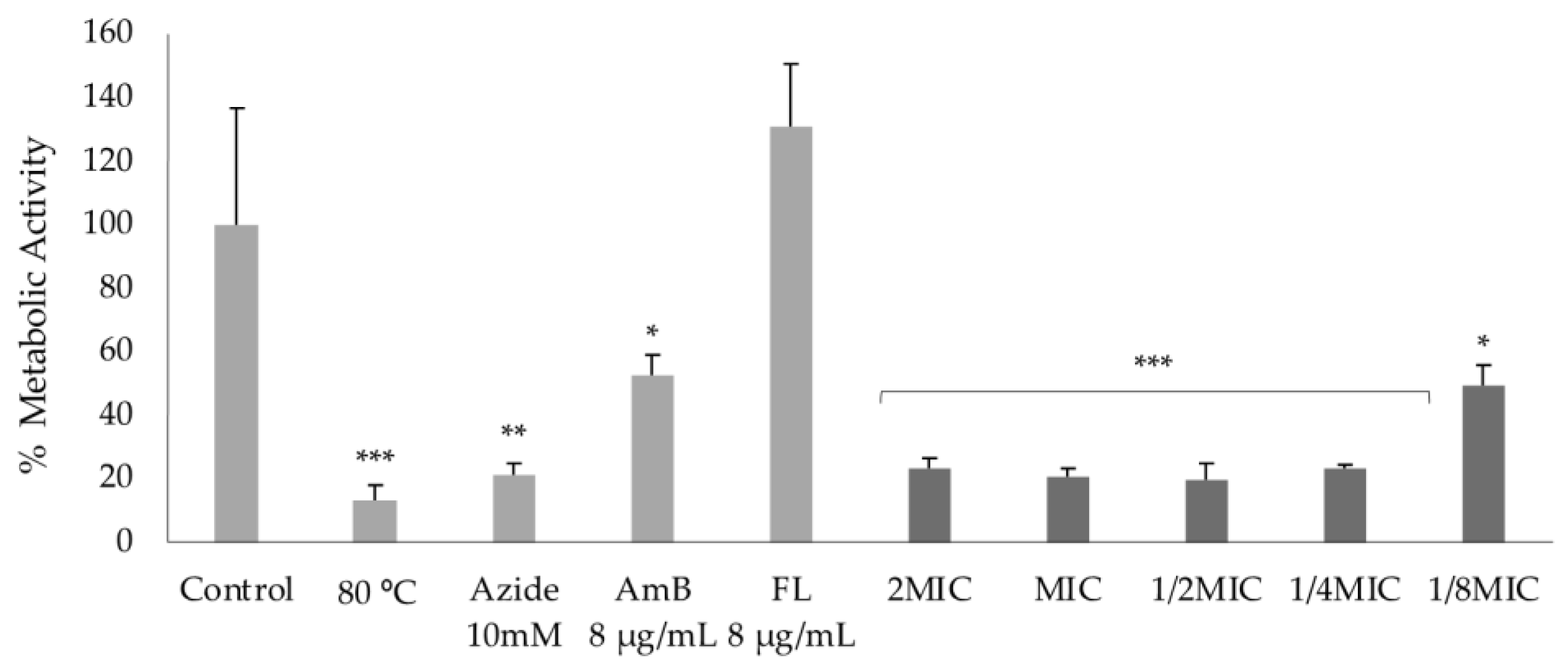
2.4.1. Effect on Cell Mitochondrial Function
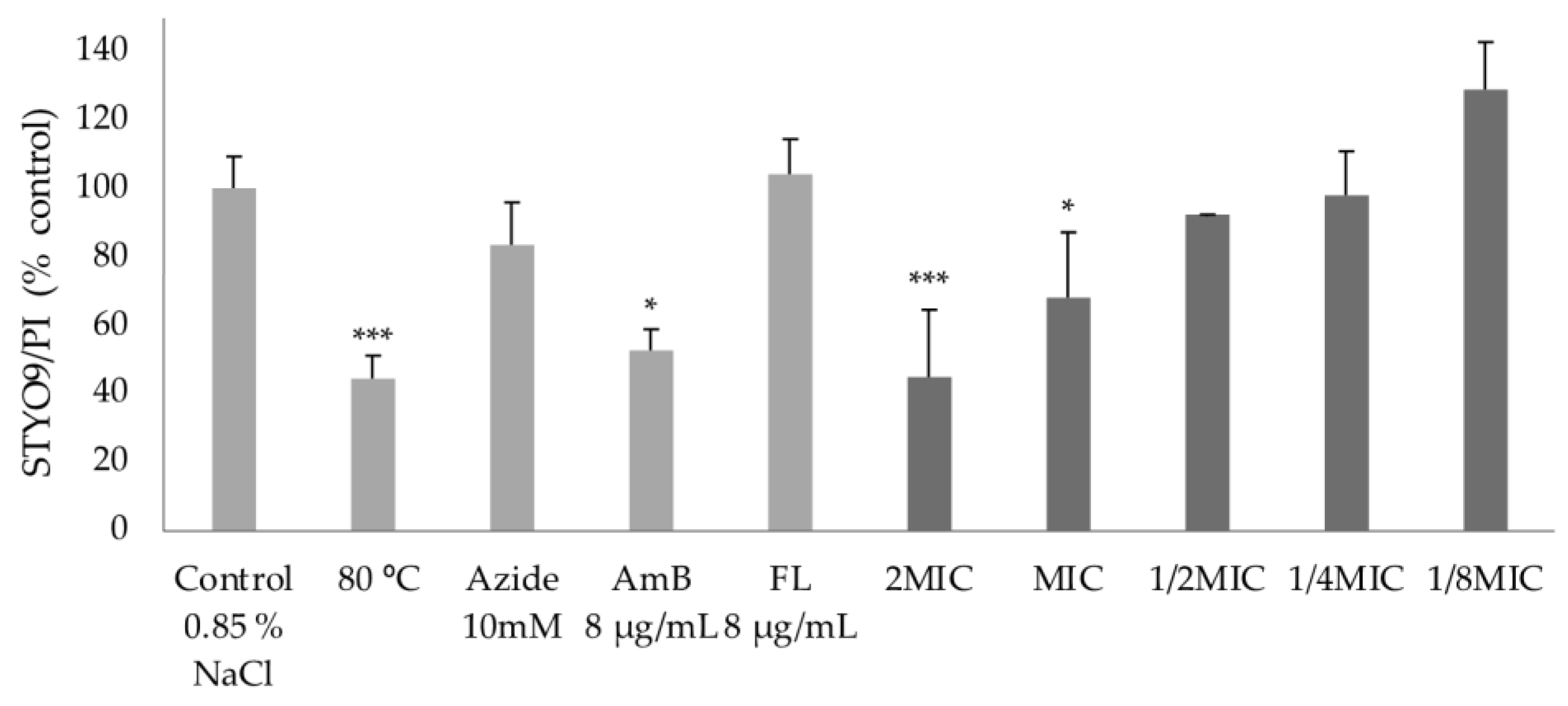
2.4.2. Effect on Membrane Integrity
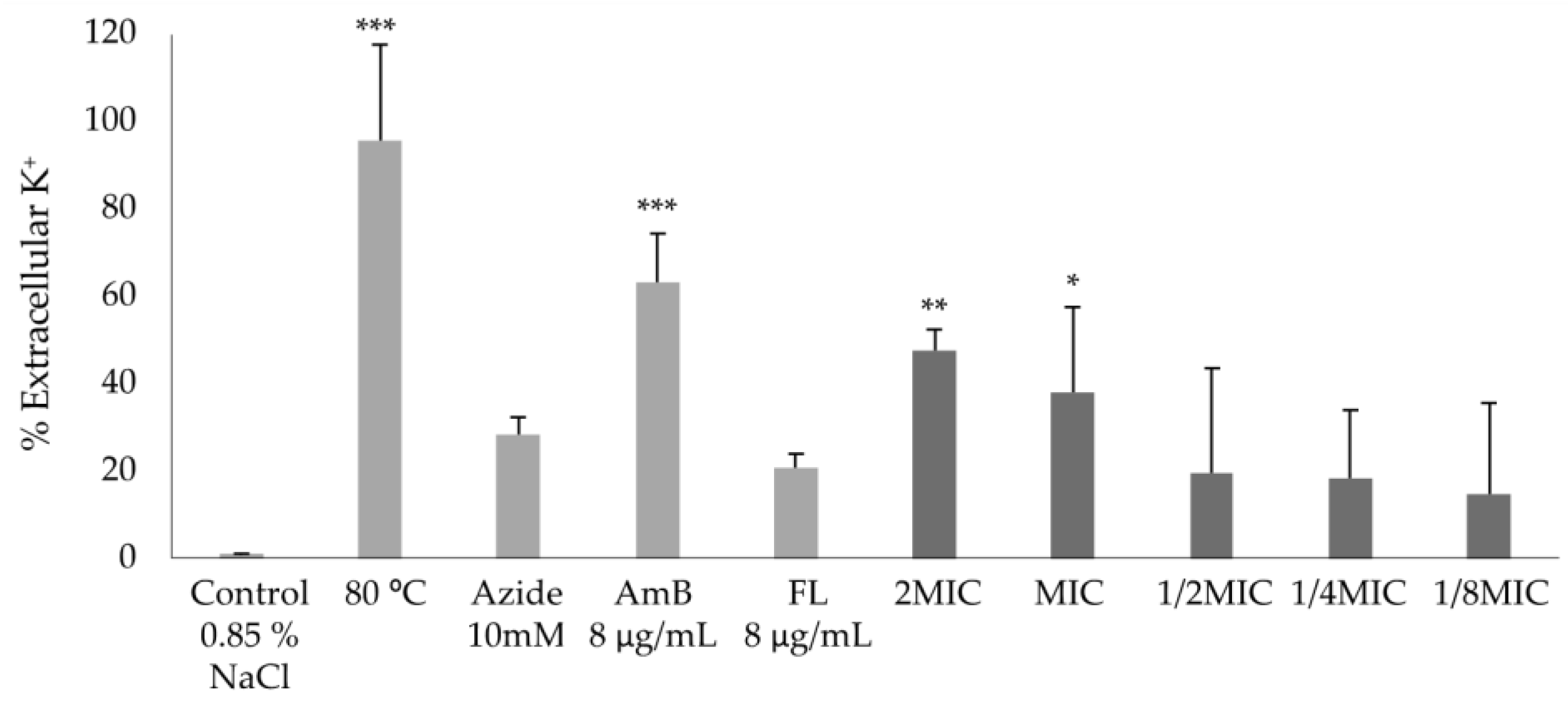
2.4.3. Effect on Potassium Efflux
2.4.4. Effect on Membrane Ergosterol
3. Materials and Methods
3.1. Chemistry
3.1.1. Material and General Methods
3.1.2. Synthesis of 1-Aminated-4-propoxy-9H-thioxanthen-9-one (1–7)
3.1.3. Synthesis of 1-{[2-(Diethylamino)ethyl]amino}-4-hydroxy-9H-thioxanthen-9-one (8)
3.1.4. Synthesis of 1,4-Bis{[2-(diethylamino)ethyl]amino}-9H-thioxanthen-9-one (9)
3.1.5. Synthesis of 4-((1-(5-Allyl-2-hydroxy-3-methoxyphenyl)-1H-1,2,3-triazol-4-yl)methoxy)-1-((2-(diethylamino)ethyl)amino)-9H-thioxanthen-9-one (10)
3.2. Microbiology
3.2.1. Compounds Preparation
3.2.2. Fungal Strains
3.2.3. Antifungal Activity
3.2.4. Germ Tube Inhibition Assay
3.2.5. Biofilm Formation Inhibition Assay
3.2.6. Yeast Metabolic Viability
3.2.7. Live/Dead Assay
3.2.8. Potassium Efflux
3.2.9. Ergosterol Binding Assay
3.2.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lockhart, S.R.; Guarner, J. Emerging and reemerging fungal infections. Semin. Diagn. Pathol. 2019, 36, 177–181. [Google Scholar] [CrossRef]
- Campoy, S.; Adrio, J.L. Antifungals. Biochem. Pharmacol. 2017, 133, 86–96. [Google Scholar] [CrossRef]
- Peres, N.T.; Maranhao, F.C.; Rossi, A.; Martinez-Rossi, N.M. Dermatophytes: Host-pathogen interaction and antifungal resistance. An. Bras. Dermatol. 2010, 85, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, N.; Pujalte, G.G.; Reese, S.T. Superficial Fungal Infections. Prim. Care 2015, 42, 501–516. [Google Scholar] [CrossRef]
- De Hoog, S.; Tang, C.; Kong, X.; Kandemir, H.; Jiang, Y. Introduction to Dermatophytes. In Dermatophytes and Dermatophytoses; Bouchara, J.-P., Nenoff, P., Gupta, A.K., Chaturvedi, V., Eds.; Springer: Cham, Switzerland, 2021; pp. 3–12. [Google Scholar] [CrossRef]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, E383–E392. [Google Scholar] [CrossRef]
- Ngo, H.X.; Garneau-Tsodikova, S.; Green, K.D. A complex game of hide and seek: The search for new antifungals. MedChemComm 2016, 7, 1285–1306. [Google Scholar] [CrossRef] [Green Version]
- Fuentefria, A.M.; Pippi, B.; Dalla Lana, D.F.; Donato, K.K.; de Andrade, S.F. Antifungals discovery: An insight into new strategies to combat antifungal resistance. Lett. Appl. Microbiol. 2018, 66, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.M.M.; Palmeira, A.; Fernandes, C.; Resende, D.; Sousa, E.; Cidade, H.; Tiritan, M.E.; Correia-da-Silva, M.; Cravo, S. From Natural Products to New Synthetic Small Molecules: A Journey through the World of Xanthones. Molecules 2021, 26, 431. [Google Scholar] [CrossRef]
- Paiva, A.M.; Pinto, M.M.; Sousa, E. A century of thioxanthones: Through synthesis and biological applications. Curr. Med. Chem. 2013, 20, 2438–2457. [Google Scholar] [CrossRef]
- Durães, F.; Palmeira, A.; Cruz, B.; Freitas-Silva, J.; Szemerédi, N.; Gales, L.; da Costa, P.M.; Remião, F.; Silva, R.; Pinto, M.; et al. Antimicrobial Activity of a Library of Thioxanthones and Their Potential as Efflux Pump Inhibitors. Pharmaceuticals 2021, 14, 572. [Google Scholar] [CrossRef]
- Bessa, L.J.; Palmeira, A.; Gomes, A.S.; Vasconcelos, V.; Sousa, E.; Pinto, M.; Martins da Costa, P. Synergistic Effects Between Thioxanthones and Oxacillin Against Methicillin-Resistant Staphylococcus aureus. Microb. Drug Resist. 2015, 21, 404–415. [Google Scholar] [CrossRef]
- Resende, D.I.S.P.; Pereira-Terra, P.; Inácio, Â.S.; Costa, P.M.d.; Pinto, E.; Sousa, E.; Pinto, M.M.M. Lichen Xanthones as Models for New Antifungal Agents. Molecules 2018, 23, 2617. [Google Scholar] [CrossRef] [Green Version]
- Pinto, E.; Afonso, C.; Duarte, S.; Vale-Silva, L.; Costa, E.; Sousa, E.; Pinto, M. Antifungal activity of xanthones: Evaluation of their effect on ergosterol biosynthesis by high-performance liquid chromatography. Chem. Biol. Drug Des. 2011, 77, 212–222. [Google Scholar] [CrossRef]
- Palmeira, A.; Vasconcelos, M.H.; Paiva, A.; Fernandes, M.X.; Pinto, M.; Sousa, E. Dual inhibitors of P-glycoprotein and tumor cell growth: (Re)discovering thioxanthones. Biochem. Pharmacol. 2012, 83, 57–68. [Google Scholar] [CrossRef]
- Archer, S.; Miller, K.J.; Rej, R.; Periana, C.; Fricker, L. Ring-hydroxylated analogues of lucanthone as antitumor agents. J. Med. Chem. 1982, 25, 220–227. [Google Scholar] [CrossRef]
- Wu, L.; Burgess, K. Synthesis and spectroscopic properties of rosamines with cyclic amine substituents. J. Org. Chem. 2008, 73, 8711–8718. [Google Scholar] [CrossRef]
- Marchese, A.; Barbieri, R.; Coppo, E.; Orhan, I.E.; Daglia, M.; Nabavi, S.F.; Izadi, M.; Abdollahi, M.; Nabavi, S.M.; Ajami, M. Antimicrobial activity of eugenol and essential oils containing eugenol: A mechanistic viewpoint. Crit. Rev. Microbiol. 2017, 43, 668–689. [Google Scholar] [CrossRef]
- Kazeminejad, Z.; Marzi, M.; Shiroudi, A.; Kouhpayeh, S.A.; Farjam, M.; Zarenezhad, E. Novel 1, 2, 4-Triazoles as Antifungal Agents. Biomed. Res. Int. 2022, 2022, 4584846. [Google Scholar] [CrossRef]
- Sydnes, M.O.; Doi, I.; Ohishi, A.; Kuse, M.; Isobe, M. Determination of solvent-trapped products obtained by photolysis of aryl azides in 2,2,2-trifluoroethanol. Chem. Asian J. 2008, 3, 102–112. [Google Scholar] [CrossRef]
- Azevedo-Barbosa, H.; Ferreira-Silva, G.; Silva, C.F.; de Souza, T.B.; Dias, D.F.; de Paula, A.C.C.; Ionta, M.; Carvalho, D.T. Phenylpropanoid-based sulfonamide promotes cyclin D1 and cyclin E down-regulation and induces cell cycle arrest at G1/S transition in estrogen positive MCF-7 cell line. Toxicol. In Vitro 2019, 59, 150–160. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, Approved Standard, 2nd ed.; Document M38-A2; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, Approved Standard, 3rd ed.; Document M27-A3; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, J.; Nakayama, D.G.; Sousa, E.; Pinto, E. Marine-Derived Compounds and Prospects for Their Antifungal Application. Molecules 2020, 25, 5856. [Google Scholar] [CrossRef] [PubMed]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of Antifungal Drug Resistance. Cold Spring Harb. Perspect. Med. 2014, 5, a019752. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.H.; Ma, Y.Y.; Ding, Y.; Chen, X.Q.; Gao, G.X. An insight into new strategies to combat antifungal drug resistance. Drug Des. Dev. Ther. 2018, 12, 3807–3816. [Google Scholar] [CrossRef] [Green Version]
- Pierce, C.G.; Chaturvedi, A.K.; Lazzell, A.L.; Powell, A.T.; Saville, S.P.; McHardy, S.F.; Lopez-Ribot, J.L. A Novel Small Molecule Inhibitor of Candida albicans Biofilm Formation, Filamentation and Virulence with Low Potential for the Development of Resistance. NPJ Biofilms Microbiomes 2015, 1, 15012. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, J.V.; Mitchell, A.P.; Andes, D.R. Fungal biofilms, drug resistance, and recurrent infection. Cold Spring Harb. Perspect. Med. 2014, 4, a019729. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Singh, S. Biofilm formation by Aspergillus fumigatus. Med. Mycol. 2014, 52, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Pereira, F.O. A review of recent research on antifungal agents against dermatophyte biofilms. Med. Mycol. 2021, 59, 313–326. [Google Scholar] [CrossRef]
- Mowat, E.; Butcher, J.; Lang, S.; Williams, C.; Ramage, G. Development of a simple model for studying the effects of antifungal agents on multicellular communities of Aspergillus fumigatus. J. Med. Microbiol. 2007, 56, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Hou, Y.; Chen, X.; Gao, Y.; Li, H.; Sun, S. Combination of fluconazole with non-antifungal agents: A promising approach to cope with resistant Candida albicans infections and insight into new antifungal agent discovery. Int. J. Antimicrob. Agents 2014, 43, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Pinto, E.; Andrade, P.B.; Valentao, P. Antifungal activity of phlorotannins against dermatophytes and yeasts: Approaches to the mechanism of action and influence on Candida albicans virulence factor. PLoS ONE 2013, 8, e72203. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zeng, H.; Tian, J.; Ban, X.; Ma, B.; Wang, Y. Antifungal mechanism of essential oil from Anethum graveolens seeds against Candida albicans. J. Med. Microbiol. 2013, 62, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Pina-Vaz, C.; Rodrigues, A.G.; Sansonetty, F.; Martinez-De-Oliveira, J.; Fonseca, A.F.; Mårdh, P.A. Antifungal activity of local anesthetics against Candida species. Infect. Dis. Obstet. Gynecol. 2000, 8, 124–137. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Santos, G.C.; Vasconcelos, C.C.; Lopes, A.J.O.; de Sousa Cartágenes, M.D.S.; Filho, A.; do Nascimento, F.R.F.; Ramos, R.M.; Pires, E.; de Andrade, M.S.; Rocha, F.M.G.; et al. Candida Infections and Therapeutic Strategies: Mechanisms of Action for Traditional and Alternative Agents. Front. Microbiol. 2018, 9, 1351. [Google Scholar] [CrossRef]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef] [Green Version]
- Miron, D.; Battisti, F.; Silva, F.K.; Lana, A.D.; Pippi, B.; Casanova, B.; Gnoatto, S.; Fuentefria, A.; Mayorga, P.; Schapoval, E.E.S. Antifungal activity and mechanism of action of monoterpenes against dermatophytes and yeasts. Rev. Bras. Farmacogn. 2014, 24, 660–667. [Google Scholar] [CrossRef]
- Leite, M.C.; Bezerra, A.P.; de Sousa, J.P.; Guerra, F.Q.; Lima Ede, O. Evaluation of Antifungal Activity and Mechanism of Action of Citral against Candida albicans. Evid. Based Complement. Alternat. Med. 2014, 2014, 378280. [Google Scholar] [CrossRef] [Green Version]
- Guerra, F.Q.; de Araujo, R.S.; de Sousa, J.P.; Pereira Fde, O.; Mendonca-Junior, F.J.; Barbosa-Filho, J.M.; de Oliveira Lima, E. Evaluation of Antifungal Activity and Mode of Action of New Coumarin Derivative, 7-Hydroxy-6-nitro-2H-1-benzopyran-2-one, against Aspergillus spp. Evid. Based Complement. Alternat. Med. 2015, 2015, 925096. [Google Scholar] [CrossRef]
- Cerqueira, F.; Maia, M.; Gabriel, C.; Medeiros, R.; Cravo, S.; Ribeiro, A.I.; Dantas, D.; Dias, A.M.; Saraiva, L.; Raimundo, L.; et al. Mechanism of Antifungal Activity by 5-Aminoimidazole-4-Carbohydrazonamide Derivatives against Candida albicans and Candida krusei. Antibiotics 2021, 10, 183. [Google Scholar] [CrossRef]
- Bae, S.H.; Park, J.H.; Choi, H.G.; Kim, H.; Kim, S.H. Imidazole Antifungal Drugs Inhibit the Cell Proliferation and Invasion of Human Breast Cancer Cells. Biomol. Ther. 2018, 26, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Li, J.; Zhang, T.; Zou, L.; Chen, Y.; Wang, K.; Lei, Y.; Yuan, K.; Li, Y.; Lan, J.; et al. Itraconazole suppresses the growth of glioblastoma through induction of autophagy: Involvement of abnormal cholesterol trafficking. Autophagy 2014, 10, 1241–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, J.; Lima, R.T.; Sousa, D.; Gomes, A.S.; Palmeira, A.; Seca, H.; Choosang, K.; Pakkong, P.; Bousbaa, H.; Pinto, M.M.; et al. Screening a Small Library of Xanthones for Antitumor Activity and Identification of a Hit Compound which Induces Apoptosis. Molecules 2016, 21, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, R.T.; Sousa, D.; Paiva, A.M.; Palmeira, A.; Barbosa, J.; Pedro, M.; Pinto, M.M.; Sousa, E.; Vasconcelos, M.H. Modulation of Autophagy by a Thioxanthone Decreases the Viability of Melanoma Cells. Molecules 2016, 21, 1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, R.T.; Sousa, D.; Gomes, A.S.; Mendes, N.; Matthiesen, R.; Pedro, M.; Marques, F.; Pinto, M.M.; Sousa, E.; Vasconcelos, M.H. The Antitumor Activity of a Lead Thioxanthone is Associated with Alterations in Cholesterol Localization. Molecules 2018, 23, 3301. [Google Scholar] [CrossRef] [Green Version]
- Mansi, I.A.; Al-Sha’er, M.A.; Mhaidat, N.M.; Taha, M.O.; Shahin, R. Investigation of Binding Characteristics of Phosphoinositide-dependent Kinase-1 (PDK1) Co-crystallized Ligands Through Virtual Pharmacophore Modeling Leading to Novel Anti-PDK1 Hits. Med. Chem. 2020, 16, 860–880. [Google Scholar] [CrossRef]
- Pinto, E.; Queiroz, M.J.; Vale-Silva, L.A.; Oliveira, J.F.; Begouin, A.; Begouin, J.M.; Kirsch, G. Antifungal activity of synthetic di(hetero)arylamines based on the benzo[b]thiophene moiety. Bioorg. Med. Chem. 2008, 16, 8172–8177. [Google Scholar] [CrossRef]
- Costa-Orlandi, C.B.; Sardi, J.C.; Santos, C.T.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J. In vitro characterization of Trichophyton rubrum and T. mentagrophytes biofilms. Biofouling 2014, 30, 719–727. [Google Scholar] [CrossRef]
- Gucwa, K.; Milewski, S.; Dymerski, T.; Szweda, P. Investigation of the Antifungal Activity and Mode of Action of Thymus vulgaris, Citrus limonum, Pelargonium graveolens, Cinnamomum cassia, Ocimum basilicum, and Eugenia caryophyllus Essential Oils. Molecules 2018, 23, 1116. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Compounds | FL2 | ||
|---|---|---|---|---|
| 1 | 8 | 9 | ||
| Yeasts | ||||
| C. albicans ATCC 10231 | 64 (>128) | 256 (>256) | 128 (≥128) | 2 (>128) |
| C. albicans H37 | 32 (32) | 64 (64) | 128 (128) | ≥128 (>128) |
| C. albicans FF172 | 128 (>128) | >128 (>128) | >128 (>128) | 0.25 (128) |
| C. albicans FF176 | 64 (>128) | 128 (>128) | 128 (128) | 32 (>128) |
| C. albicans DSY294 | 64 (>128) | >128 (>128) | >128 (>128) | 0.125 (128) |
| C. albicans DSY296 | 128 (>128) | >128 (>128) | >128 (>128) | 64 (>128) |
| C. glabrata 10R | 32 (64) | 32 (32) | 64 (64) | 64 (>128) |
| C. glabrata DSY562 | 32 (64) | 32 (32) | 32 (32) | 4 (>128) |
| C. glabrata DSY565 | 32 (64) | 64 (128) | 64 (64) | 128 (>128) |
| C. krusei ATCC 6258 | 32 (32) | 32 (32) | 64 (128) | 32 (>128) |
| C. neoformans CECT 1078 | 16 (16) | 16 (16) | 32 (32) | 8 (>32) |
| Filamentous Fungi | ||||
| A. fumigatus ATCC 240305 | 32 (>128) | 32 (>128) | 64 (≥128) | ≥128 (>128) |
| A. fumigatus C111 | 32 (>128) | 32 (>128) | 64 (64) | ≥128 (>128) |
| A. niger ATCC16404 | 32 (>128) | 32 (>128) | 128 (>128) | ≥128 (>128) |
| A. flavus F44 | >128 (>128) | >128 (>128) | >128 (>128) | 128 (>128) |
| F. solani FF125 | 64 (>128) | 128 (>128) | >128 (>128) | ≥128 (>128) |
| F. oxysporum FF115 | 64 (128) | 64 (64) | 128 (128) | 64 (>128) |
| Scedosporium spp. | 8 (8) | 16 (16) | 32 (32) | 4 (16) |
| Lichtheimia spp. | 16 (16) | 32 (64) | 64 (64) | 64 (>128) |
| Mucor spp. | 16 (16) | 16 (16) | 32 (32) | >128 (>128) |
| T. rubrum FF5 | 16 (16) | 8 (8) | 32 (32) | 16 (64) |
| T. mentagrophytes FF7 | 16 (16) | 16 (32) | 32 (64) | 8 (32) |
| N. gypsea FF3 | 16 (32) | 16 (16) | 32 (64) | 32 (≥128) |
| M. canis FF1 | 32 (64) | 16 (32) | 32 (64) | 32 (≥128) |
| Strains | Compound 1 | AmB 1 | FL 1 | |||
|---|---|---|---|---|---|---|
| Absence | 400 | Absence | 400 | Absence | 400 | |
| C. albicans ATCC 10231 | 64 | 64 | 2 | 16 | 2 | 2 |
| A. fumigatus ATCC 240305 | 32 | 32 | 0.5 | 64 | 128–256 | 128 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso, J.; Freitas-Silva, J.; Durães, F.; Carvalho, D.T.; Gales, L.; Pinto, M.; Sousa, E.; Pinto, E. Antifungal Activity of a Library of Aminothioxanthones. Antibiotics 2022, 11, 1488. https://doi.org/10.3390/antibiotics11111488
Cardoso J, Freitas-Silva J, Durães F, Carvalho DT, Gales L, Pinto M, Sousa E, Pinto E. Antifungal Activity of a Library of Aminothioxanthones. Antibiotics. 2022; 11(11):1488. https://doi.org/10.3390/antibiotics11111488
Chicago/Turabian StyleCardoso, Joana, Joana Freitas-Silva, Fernando Durães, Diogo Teixeira Carvalho, Luís Gales, Madalena Pinto, Emília Sousa, and Eugénia Pinto. 2022. "Antifungal Activity of a Library of Aminothioxanthones" Antibiotics 11, no. 11: 1488. https://doi.org/10.3390/antibiotics11111488