



Study on Optimizing Novel Antimicrobial Peptides with Bifunctional Activity to Prevent and Treat Peri-Implant Disease
 ,
,
Abstract
:1. Introduction
2. Results
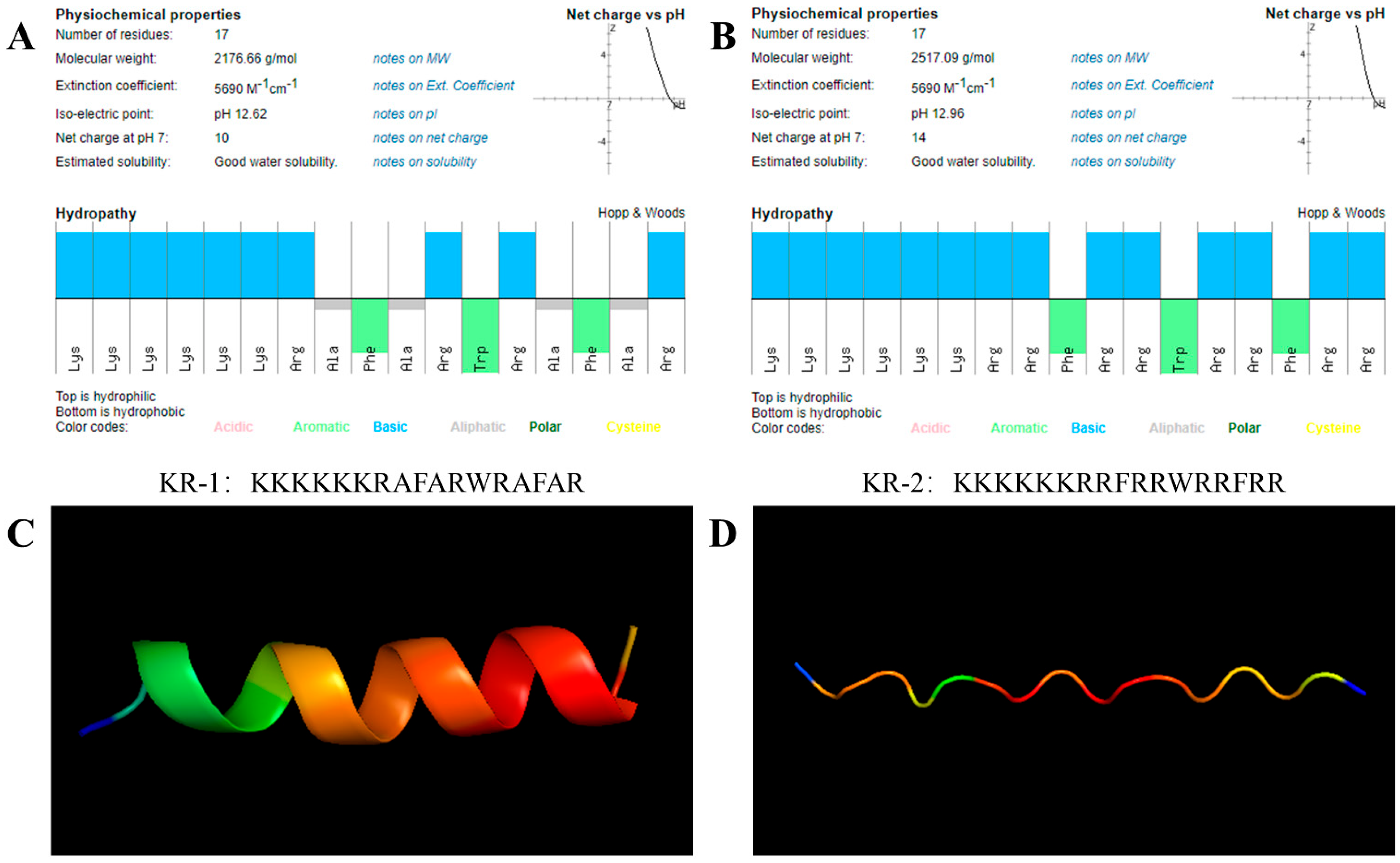
2.1. Properties of Antimicrobial Peptides
2.2. Antimicrobial Activity against S. gordonii and F. nucleatum
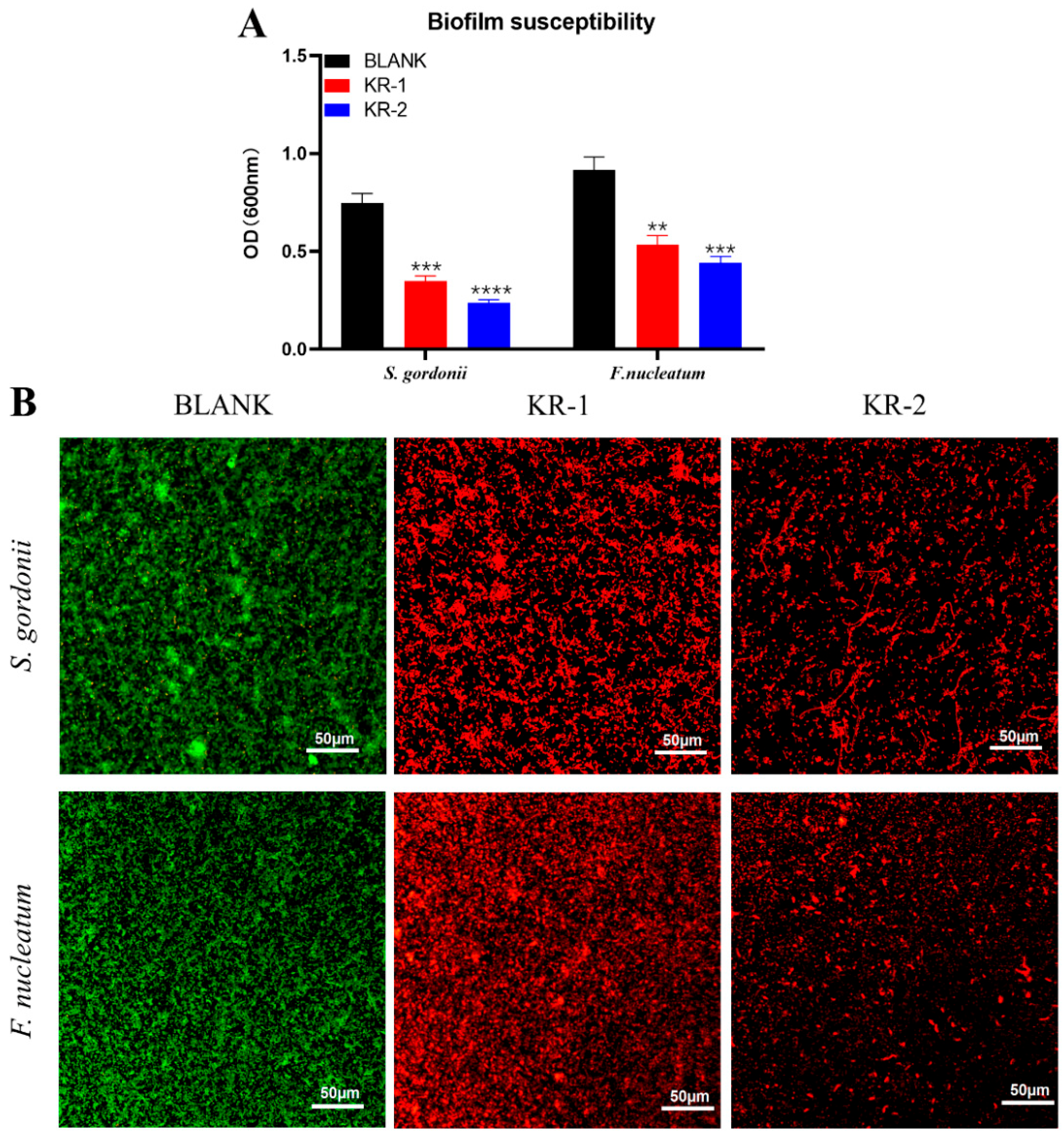
2.3. Biofilm Inhibition
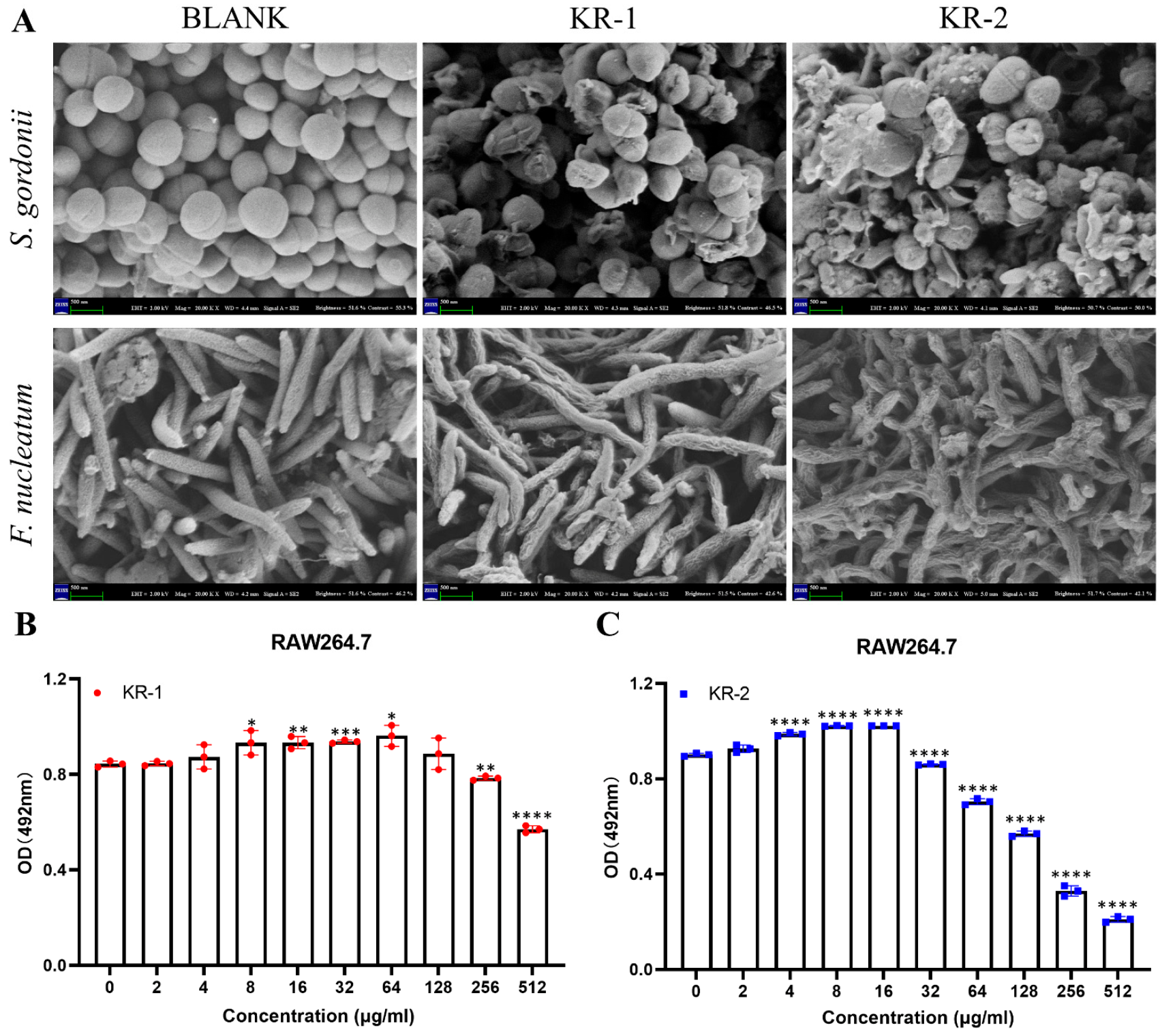
2.4. Scanning Electron Microscope (SEM)
2.5. Biocompatibility of Antimicrobial Peptides
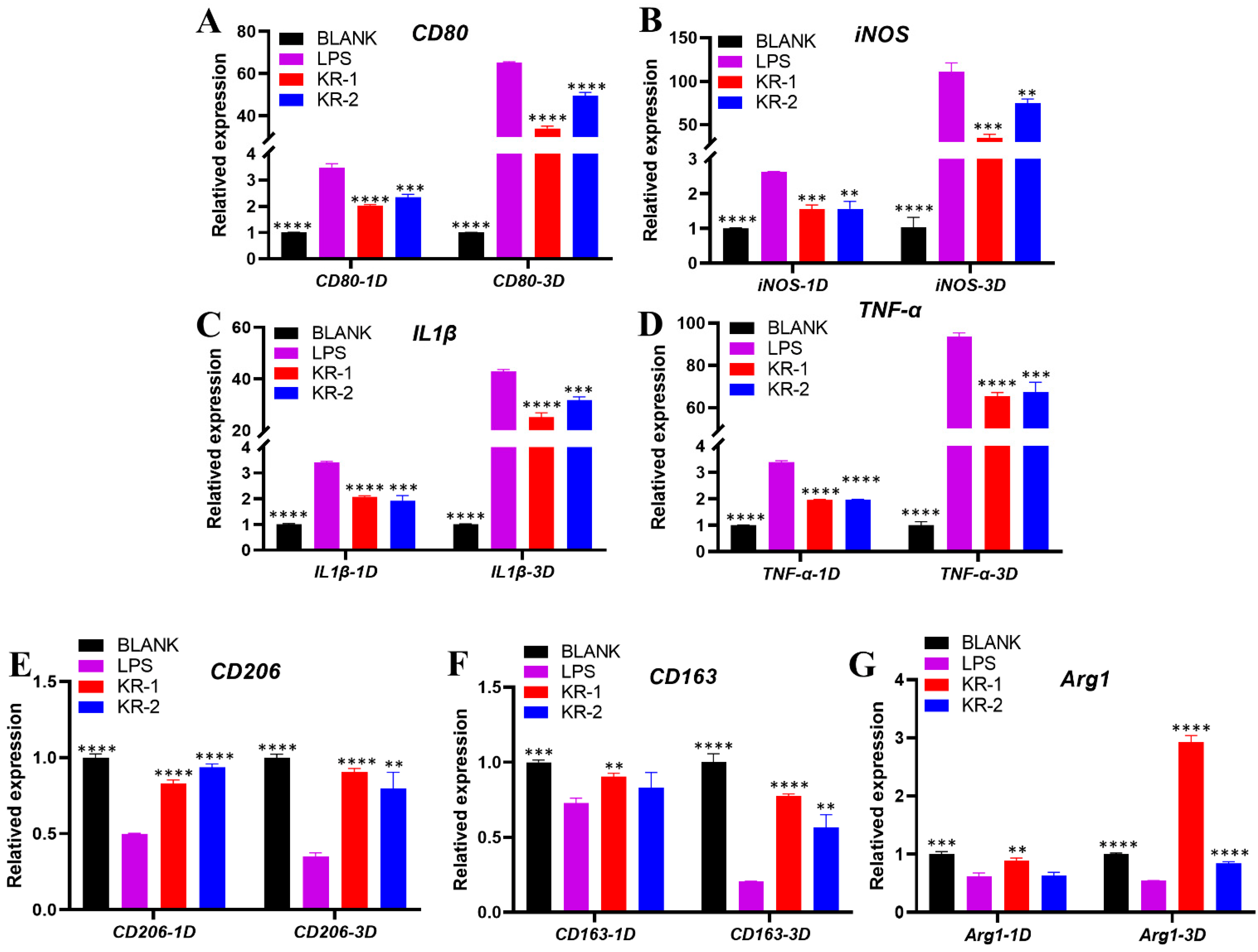
2.6. Effect of Peptides on the Expression of Pro-Inflammatory and Anti-Inflammatory Genes of RAW 264.7
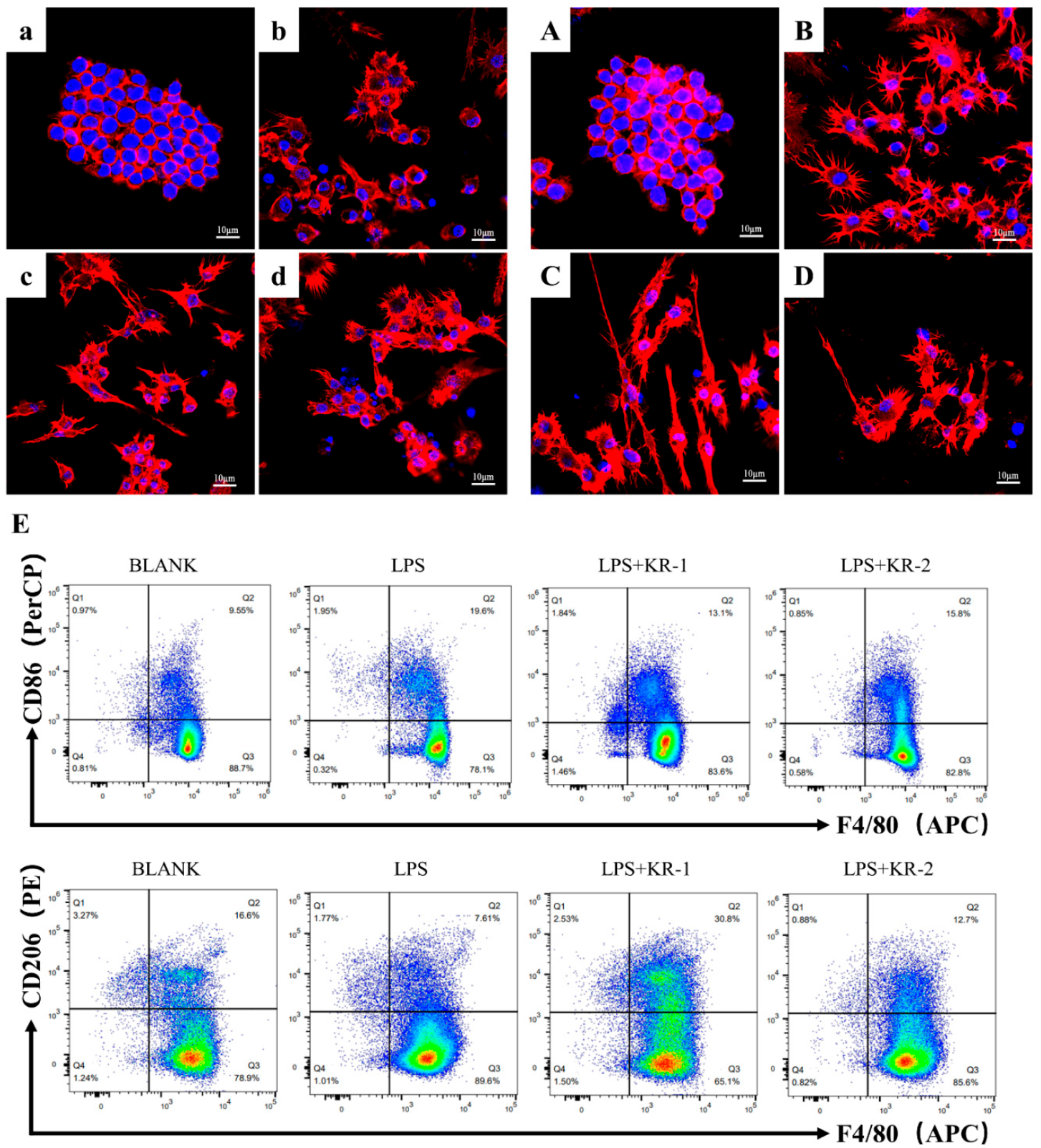
2.7. Effects of Antimicrobial Peptides on Polarized Morphology of RAW264.7
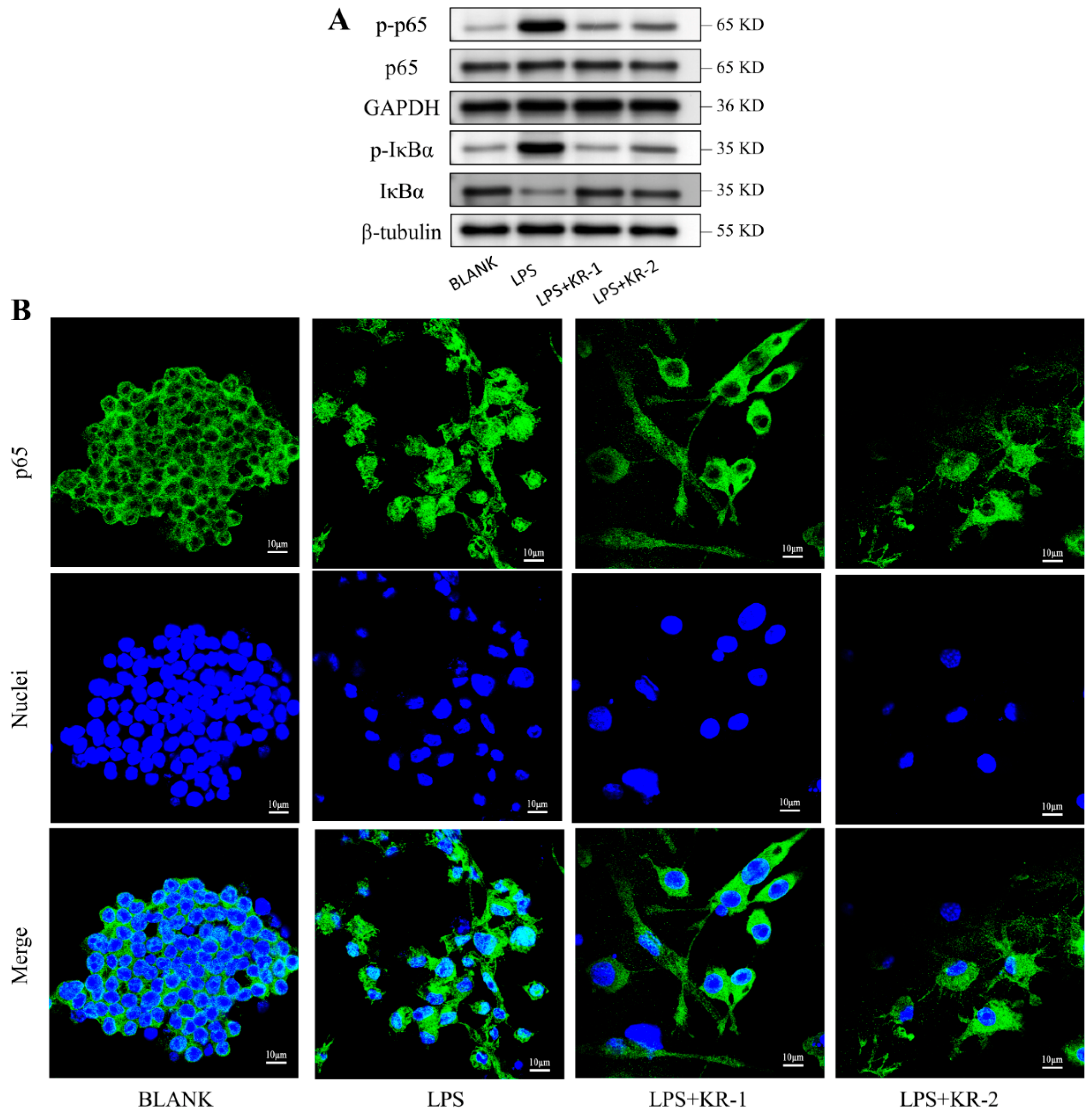
2.8. Antimicrobial Peptides Pair Regulation of NF-κB-p65 Signal Pathway during RAW 264.7 Polarization
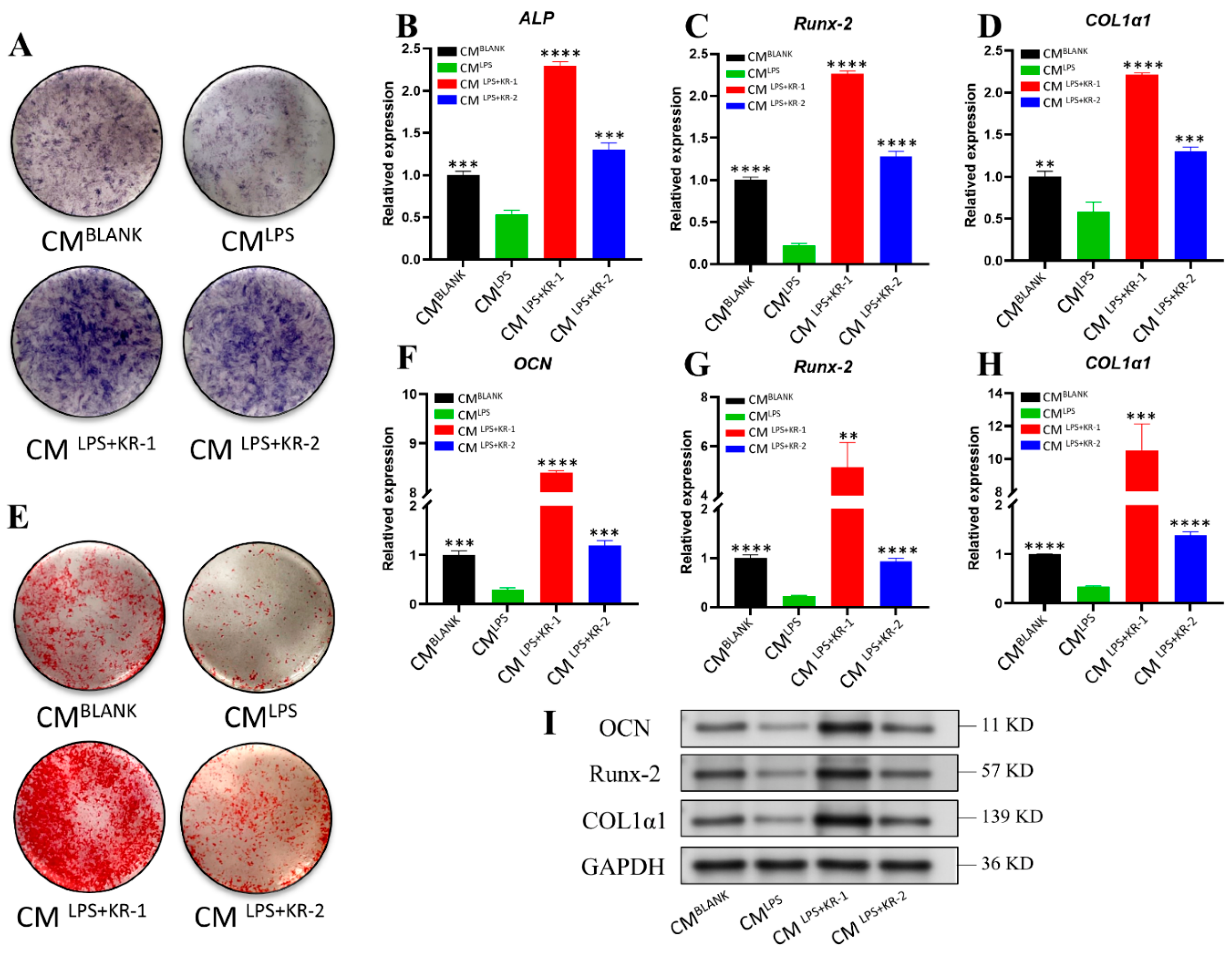
2.9. Regulation of Conditioned Medium on Osteogenic Ability of Osteoblasts MC3T3-E1
3. Discussion
4. Materials and Methods
4.1. Synthesis and Characterization of Antimicrobial Peptides
4.2. Behaviors of S. gordonii and F. nucleatum Cultured with the Different Antimicrobial Peptides
4.2.1. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
4.2.2. Biofilm Susceptibility Assay
4.2.3. Confocal Laser Scanning Microscopy (CLSM)
4.2.4. Scanning Electron Microscopy (SEM)
4.3. Behaviors of Macrophages RAW 264.7 Cultured with the Different Antimicrobial Peptides
4.3.1. Cell Culture
4.3.2. Cell Proliferation Assay
4.3.3. Gene Expression of Cell Polarization
4.3.4. Cell Morphology and p65 Immunofluorescence Staining
4.3.5. Flow Cytometry
4.4. Behavior of MC3T3-E1 Cells in Different Antimicrobial Peptides Conditioned Media (CM)
4.4.1. Preparation of Conditioned Medium
4.4.2. Cell Culture
4.4.3. Osteogenic-Related Gene Expression
4.4.4. Alkaline Phosphatase Staining Assay
4.4.5. Alizarin Red S (ARS) Staining
4.4.6. Western Blot Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bassir, S.H.; Kholy, K.E.; Chen, C.Y.; Lee, K.H.; Intini, G. Outcome of early dental implant placement versus other dental implant placement protocols: A systematic review and meta-analysis. J. Periodontol. 2019, 90, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Prestat, M.; Thierry, D. Corrosion of titanium under simulated inflammation conditions: Clinical context and in vitro investigations. Acta Biomater. 2021, 136, 72–87. [Google Scholar] [CrossRef]
- Stich, T.; Alagboso, F.; Krenek, T.; Kovarik, T.; Alt, V.; Docheva, D. Implant-bone-interface: Reviewing the impact of titanium surface modifications on osteogenic processes in vitro and in vivo. Bioeng. Transl. Med. 2022, 7, e10239. [Google Scholar] [CrossRef]
- Hanawa, T. Zirconia versus titanium in dentistry: A review. Dent. Mater. J. 2020, 39, 24–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paparella, M.L.; Domingo, M.G.; Puia, S.A.; Jacobi-Gresser, E.; Olmedo, D.G. Titanium dental implant-related pathologies: A retrospective histopathological study. Oral Dis. 2022, 28, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Gulati, K.; Arora, H.; Han, P.; Fournier, B.; Ivanovski, S. Orchestrating soft tissue integration at the transmucosal region of titanium implants. Acta Biomater. 2021, 124, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Karazisis, D.; Rasmusson, L.; Petronis, S.; Palmquist, A.; Shah, F.A.; Agheli, H.; Emanuelsson, L.; Johansson, A.; Omar, O.; Thomsen, P. The effects of controlled nanotopography, machined topography and their combination on molecular activities, bone formation and biomechanical stability during osseointegration. Acta Biomater. 2021, 136, 279–290. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.M.; Risser, G.E.; Spiller, K.L. Sequential drug delivery to modulate macrophage behavior and enhance implant integration. Adv. Drug Deliv. Rev. 2019, 149–150, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Chen, P.; Zhao, Y.; Hang, R.; Yao, X.; Tang, B.; Liu, C.; Xiao, Y.; Hang, R. A micro/nano-biomimetic coating on titanium orchestrates osteo/angio-genesis and osteoimmunomodulation for advanced osseointegration. Biomaterials 2021, 278, 121162. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Lee, K.K.; Park, K.W.; Kim, M.; Lee, C.S. Genetically Modified Ferritin Nanoparticles with Bone-Targeting Peptides for Bone Imaging. Int. J. Mol. Sci. 2021, 22, 4854. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ye, X.; Zhao, Y.; Bai, L.; He, Z.; Tong, Q.; Xie, X.; Zhu, H.; Cai, D.; Zhou, Y.; et al. Cryogenic 3D printing of porous scaffolds for in situ delivery of 2D black phosphorus nanosheets, doxorubicin hydrochloride and osteogenic peptide for treating tumor resection-induced bone defects. Biofabrication 2020, 12, 035004. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.; Beg, S.; Panda, S.K.; Rahman, M.; Alharbi, K.S.; Jain, G.K.; Ahmad, F.J. Novel therapeutic interventions in cancer treatment using protein and peptide-based targeted smart systems. Semin. Cancer Biol. 2021, 69, 249–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Huang, Y.; Lindstrom, A.R.; Lin, T.Y.; Lam, K.S.; Li, Y. Peptide-based materials for cancer immunotherapy. Theranostics 2019, 9, 7807–7825. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Bi, X.; Wu, Y.; Pan, M.; Ma, X.; Mo, L.; Wang, J.; Li, X. Cationic self-assembled peptide-based molecular hydrogels for extended ocular drug delivery. Acta Biomater. 2021, 131, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.Y.; Steinbach-Rankins, J.M.; Demuth, D.R. Functional assessment of peptide-modified PLGA nanoparticles against oral biofilms in a murine model of periodontitis. J. Control. Release 2019, 297, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Hojabri, N.; Kaisarly, D.; Kunzelmann, K.H. Adhesion and whitening effects of P11-4 self-assembling peptide and HAP suspension on bovine enamel. Clin. Oral Investig. 2021, 25, 3237–3247. [Google Scholar] [CrossRef] [PubMed]
- Buda De Cesare, G.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. Antimicrobial Peptides: A New Frontier in Antifungal Therapy. mBio 2020, 11, e02123-20. [Google Scholar] [CrossRef] [PubMed]
- Schutz, D.; Ruiz-Blanco, Y.B.; Munch, J.; Kirchhoff, F.; Sanchez-Garcia, E.; Muller, J.A. Peptide and peptide-based inhibitors of SARS-CoV-2 entry. Adv. Drug Deliv. Rev. 2020, 167, 47–65. [Google Scholar] [CrossRef]
- Rizvi, S.F.A.; Zhang, H. Emerging trends of receptor-mediated tumor targeting peptides: A review with perspective from molecular imaging modalities. Eur. J. Med. Chem. 2021, 221, 113538. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Li, X.; Du, H.; Jiang, C.; Xu, S.; Cao, Y. Immunomodulatory significance of natural peptides in mammalians: Promising agents for medical application. Immunobiology 2020, 225, 151936. [Google Scholar] [CrossRef]
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.Y. Antimicrobial peptides: New hope in the war against multidrug resistance. Zool. Res. 2019, 40, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.A.; Cole, G.B.; Ravamehr-Lake, D.; Nguyen, H.Q.; Khan, F.; Sharpe, S.; Deber, C.M. Positive Charge Patterning and Hydrophobicity of Membrane-Active Antimicrobial Peptides as Determinants of Activity, Toxicity, and Pharmacokinetic Stability. J. Med. Chem. 2019, 62, 6276–6286. [Google Scholar] [CrossRef]
- Beaudoin, T.; Stone, T.A.; Glibowicka, M.; Adams, C.; Yau, Y.; Ahmadi, S.; Bear, C.E.; Grasemann, H.; Waters, V.; Deber, C.M. Activity of a novel antimicrobial peptide against Pseudomonas aeruginosa biofilms. Sci. Rep. 2018, 8, 14728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laselva, O.; Stone, T.A.; Bear, C.E.; Deber, C.M. Anti-Infectives Restore ORKAMBI((R)) Rescue of F508del-CFTR Function in Human Bronchial Epithelial Cells Infected with Clinical Strains of P. aeruginosa. Biomolecules 2020, 10, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feger, G.; Angelov, B.; Angelova, A. Prediction of Amphiphilic Cell-Penetrating Peptide Building Blocks from Protein-Derived Amino Acid Sequences for Engineering of Drug Delivery Nanoassemblies. J. Phys. Chem. B 2020, 124, 4069–4078. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guan, M.; Ren, R.; Gao, C.; Cheng, H.; Li, Y.; Gao, B.; Wei, Y.; Fu, J.; Sun, J.; et al. Improved Immunoregulation of Ultra-Low-Dose Silver Nanoparticle-Loaded TiO2 Nanotubes via M2 Macrophage Polarization by Regulating GLUT1 and Autophagy. Int. J. Nanomed. 2020, 15, 2011–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; You, Y.; Ma, A.; Song, Y.; Jiao, J.; Song, L.; Shi, E.; Zhong, X.; Li, Y.; Li, C. Zn-Incorporated TiO2 Nanotube Surface Improves Osteogenesis Ability through Influencing Immunomodulatory Function of Macrophages. Int. J. Nanomed. 2020, 15, 2095–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, F.N.; Jensen, S.S.; Dahl, M. Implant treatment after traumatic tooth loss: A systematic review. Dent. Traumatol. 2022, 38, 105–116. [Google Scholar] [CrossRef]
- Naseri, R.; Yaghini, J.; Feizi, A. Levels of smoking and dental implants failure: A systematic review and meta-analysis. J. Clin. Periodontol. 2020, 47, 518–528. [Google Scholar] [CrossRef]
- Li, B.; Cai, Q.; Wang, Z.; Qiao, S.; Ou, Y.; Ma, R.; Luo, C.; Meng, W. D-arginine Enhances the Effect of Alpha-Amylase on Disassembling Actinomyces viscosus Biofilm. Front. Bioeng. Biotechnol. 2022, 10, 864012. [Google Scholar] [CrossRef] [PubMed]
- Hwang, G. In it together: Candida-bacterial oral biofilms and therapeutic strategies. Environ. Microbiol. Rep. 2022, 14, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Eger, M.; Liron, T.; Hiram-Bab, S.; Awida, Z.; Giladi, E.; Dangoor, D.; Fridkin, M.; Kohavi, D.; Gozes, I.; Gabet, Y. Therapeutic Potential of Vasoactive Intestinal Peptide and its Derivative Stearyl-Norleucine-VIP in Inflammation-Induced Osteolysis. Front. Pharmacol. 2021, 12, 638128. [Google Scholar] [CrossRef] [PubMed]
- Kartikasari, N.; Yamada, M.; Watanabe, J.; Tiskratok, W.; He, X.; Kamano, Y.; Egusa, H. Titanium surface with nanospikes tunes macrophage polarization to produce inhibitory factors for osteoclastogenesis through nanotopographic cues. Acta Biomater. 2022, 137, 316–330. [Google Scholar] [CrossRef]
- Ma, Q.L.; Fang, L.; Jiang, N.; Zhang, L.; Wang, Y.; Zhang, Y.M.; Chen, L.H. Bone mesenchymal stem cell secretion of sRANKL/OPG/M-CSF in response to macrophage-mediated inflammatory response influences osteogenesis on nanostructured Ti surfaces. Biomaterials 2018, 154, 234–247. [Google Scholar] [CrossRef]
- Qoreishi, M.; Panahi, M.; Dorodi, O.; Ghanbari, N.; Jousheghan, S.S. Involvement of NF-κB/NLRP3 axis in the progression of aseptic loosening of total joint arthroplasties: A review of molecular mechanisms. Naunyn Schmiedebergs Arch. Pharmacol. 2022, 395, 757–767. [Google Scholar] [CrossRef]
- Moshaverinia, A.; Chen, C.; Xu, X.; Ansari, S.; Zadeh, H.H.; Schricker, S.R.; Paine, M.L.; Moradian-Oldak, J.; Khademhosseini, A.; Snead, M.L.; et al. Regulation of the Stem Cell-Host Immune System Interplay Using Hydrogel Coencapsulation System with an Anti-Inflammatory Drug. Adv. Funct. Mater. 2015, 25, 2296–2307. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Wang, H.; Chen, H.; Ge, G.; Wang, M.; Gao, A.; Tong, L.; Xu, Y.; Yang, H.; Pan, G.; et al. Biomimetic osteogenic peptide with mussel adhesion and osteoimmunomodulatory functions to ameliorate interfacial osseointegration under chronic inflammation. Biomaterials 2020, 255, 120197. [Google Scholar] [CrossRef]
- Nobles, K.P.; Janorkar, A.V.; Williamson, R.S. Surface modifications to enhance osseointegration-Resulting material properties and biological responses. J. Biomed. Mater. Res. B Appl. Biomater. 2021, 109, 1909–1923. [Google Scholar] [CrossRef]
- Trier, N.; Hansen, P.; Houen, G. Peptides, Antibodies, Peptide Antibodies and More. Int. J. Mol. Sci. 2019, 20, 6289. [Google Scholar] [CrossRef]
- Zeth, K.; Sancho-Vaello, E. Structural Plasticity of LL-37 Indicates Elaborate Functional Adaptation Mechanisms to Bacterial Target Structures. Int. J. Mol. Sci. 2021, 22, 5200. [Google Scholar] [CrossRef] [PubMed]
- Francis, M.; Gopinathan, G.; Foyle, D.; Fallah, P.; Gonzalez, M.; Luan, X.; Diekwisch, T.G.H. Histone Methylation: Achilles Heel and Powerful Mediator of Periodontal Homeostasis. J. Dent. Res. 2020, 99, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Mokashi-Punekar, S.; Walsh, T.R.; Rosi, N.L. Tuning the Structure and Chiroptical Properties of Gold Nanoparticle Single Helices via Peptide Sequence Variation. J. Am. Chem. Soc. 2019, 141, 15710–15716. [Google Scholar] [CrossRef] [PubMed]
- Kabelka, I.; Vacha, R. Advances in Molecular Understanding of α-Helical Membrane-Active Peptides. Acc. Chem. Res. 2021, 54, 2196–2204. [Google Scholar] [CrossRef] [PubMed]
- Isik, M.; Tan, J.P.; Ono, R.J.; Sanchez-Sanchez, A.; Mecerreyes, D.; Yang, Y.Y.; Hedrick, J.L.; Sardon, H. Tuning the Selectivity of Biodegradable Antimicrobial Cationic Polycarbonates by Exchanging the Counter-Anion. Macromol. Biosci. 2016, 16, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Kabelka, I.; Vacha, R. Optimal Hydrophobicity and Reorientation of Amphiphilic Peptides Translocating through Membrane. Biophys. J. 2018, 115, 1045–1054. [Google Scholar] [CrossRef] [Green Version]
- Goel, P.N.; Egol, A.J.; Moharrer, Y.; Brandfield-Harvey, B.; Ahn, J.; Ashley, J.W. Notch signaling inhibition protects against LPS mediated osteolysis. Biochem. Biophys. Res. Commun. 2019, 515, 538–543. [Google Scholar] [CrossRef]
- Zeng, X.Z.; Zhang, Y.Y.; Yang, Q.; Wang, S.; Zou, B.H.; Tan, Y.H.; Zou, M.; Liu, S.W.; Li, X.J. Artesunate attenuates LPS-induced osteoclastogenesis by suppressing TLR4/TRAF6 and PLCgamma1-Ca2+-NFATc1 signaling pathway. Acta Pharmacol. Sin. 2020, 41, 229–236. [Google Scholar] [CrossRef]
- Wang, T.; Bai, J.; Lu, M.; Huang, C.; Geng, D.; Chen, G.; Wang, L.; Qi, J.; Cui, W.; Deng, L. Engineering immunomodulatory and osteoinductive implant surfaces via mussel adhesion-mediated ion coordination and molecular clicking. Nat. Commun. 2022, 13, 160. [Google Scholar] [CrossRef]
- Qiu, W.; Hu, J.; Magnuson, J.T.; Greer, J.; Yang, M.; Chen, Q.; Fang, M.; Zheng, C.; Schlenk, D. Evidence linking exposure of fish primary macrophages to antibiotics activates the NF-κB pathway. Environ. Int. 2020, 138, 105624. [Google Scholar] [CrossRef]
- Dai, F.; Du, Y.T.; Zheng, Y.L.; Zhou, B. Inhibiting NF-κB-Mediated Inflammation by Catechol-Type Diphenylbutadiene via an Intracellular Copper- and Iron-Dependent Pro-Oxidative Role. J. Agric. Food Chem. 2020, 68, 10029–10035. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Guan, P.; Xiao, C.; Wen, H.; Wang, Q.; Liu, C.; Luo, Y.; Ma, L.; Tan, G.; Yu, P.; et al. Exosome-functionalized polyetheretherketone-based implant with immunomodulatory property for enhancing osseointegration. Bioact. Mater. 2021, 6, 2754–2766. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Yan, Y.; Wu, Z.; Wei, Y.; Song, H.; Zhu, L.; Zhao, C.; Xu, N.; Fu, J.; Huo, K. Resveratrol-loaded titania nanotube coatings promote osteogenesis and inhibit inflammation through reducing the reactive oxygen species production via regulation of NF-κB signaling pathway. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 131, 112513. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | MIC (μg/mL) | MBC (μg/mL) | ||
|---|---|---|---|---|
| KR−1 | KR−2 | KR−1 | KR−2 | |
| S. gordonii | 125 | 50 | 250 | 150 |
| F. nucleatum | 100 | 50 | 200 | 100 |
| Gene (Mouse) | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| CD80 | CCTCAAGTTTCCATGTCCAAGGC | GAGGAGAGTTGTAACGGCAAGG |
| iNOS | GAGACAGGGAAGTCTGAAGCAC | CCAGCAGTAGTTGCTCCTCTTC |
| IL1β | TGGACCTTCCAGGATGAGGACA | GTTCATCTCGGAGCCTGTAGTG |
| TNF-α | GGTGCCTATGTCTCAGCCTCTT | GCCATAGAACTGATGAGAGGGAG |
| CD206 | GTTCACCTGGAGTGATGGTTCTC | AGGACATGCCAGGGTCACCTTT |
| CD163 | GGCTAGACGAAGTCATCTGCAC | CTTCGTTGGTCAGCCTCAGAGA |
| Arg-1 | CATTGGCTTGCGAGACGTAGAC | GCTGAAGGTCTCTTCCATCACC |
| ALP | CCAGAAAGACACCTTGACTGTGG | TCTTGTCCGTGTCGCTCACCAT |
| Runx-2 | CCTGAACTCTGCACCAAGTCCT | TCATCTGGCTCAGATAGGAGGG |
| COL1α1 | CCTCAGGGTATTGCTGGACAAC | CAGAAGGACCTTGTTTGCCAGG |
| OCN | GCAATAAGGTAGTGAACAGACTCC | CCATAGATGCGTTTGTAGGCGG |
| GAPDH | CATCACTGCCACCCAGAAGACTG | ATGCCAGTGAGCTTCCCGTTCAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Zhang, Q.; Hu, M.; Zhao, B.; Liu, Z.; Li, C.; Zhang, X. Study on Optimizing Novel Antimicrobial Peptides with Bifunctional Activity to Prevent and Treat Peri-Implant Disease. Antibiotics 2022, 11, 1482. https://doi.org/10.3390/antibiotics11111482
Yu S, Zhang Q, Hu M, Zhao B, Liu Z, Li C, Zhang X. Study on Optimizing Novel Antimicrobial Peptides with Bifunctional Activity to Prevent and Treat Peri-Implant Disease. Antibiotics. 2022; 11(11):1482. https://doi.org/10.3390/antibiotics11111482
Chicago/Turabian StyleYu, Shuipeng, Qian Zhang, Meilin Hu, Borui Zhao, Zhiyang Liu, Changyi Li, and Xi Zhang. 2022. "Study on Optimizing Novel Antimicrobial Peptides with Bifunctional Activity to Prevent and Treat Peri-Implant Disease" Antibiotics 11, no. 11: 1482. https://doi.org/10.3390/antibiotics11111482