
Molecular Cloning and Functional Identification of the Antimicrobial Peptide Gene Ctri9594 from the Venom of the Scorpion Chaerilus tricostatus

Abstract
:1. Introduction
2. Results
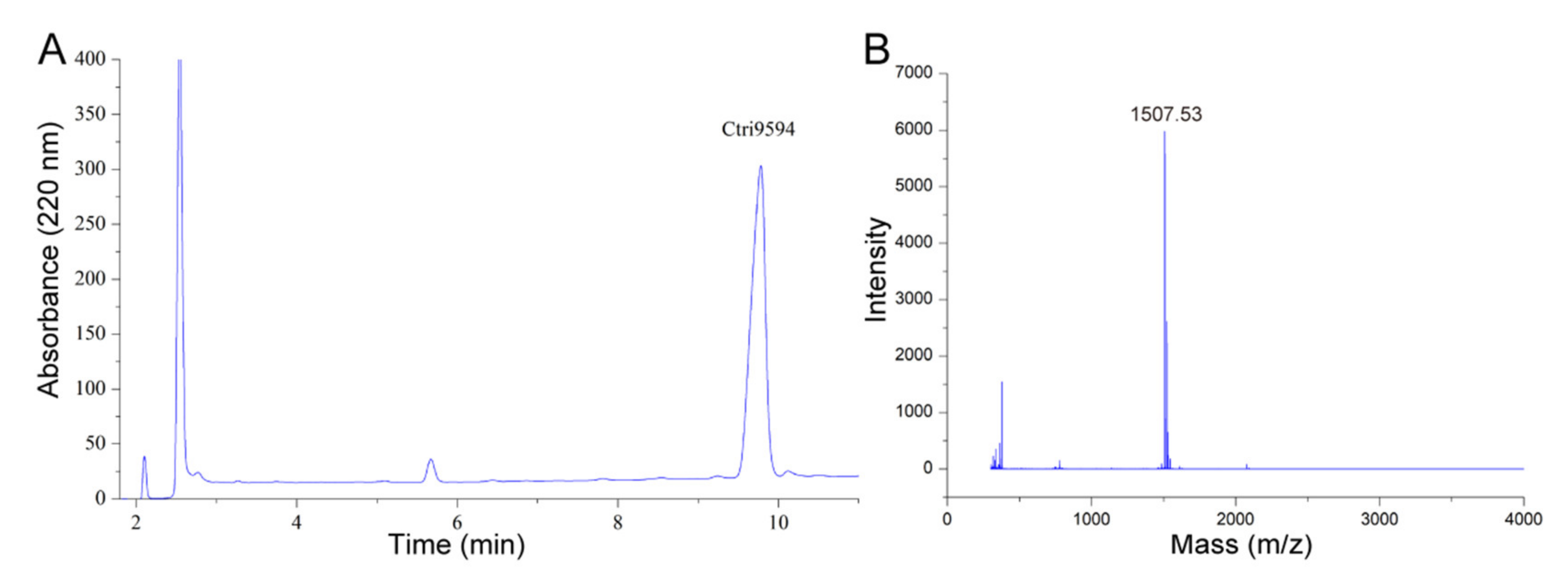
2.1. Cloning and Characterization of a Precursor Nucleotide Sequence Encoding an AMP from cDNA Library of the Venom Gland of the Scorpion Chaerilus tricostatus
2.2. Precursor Amino Acid Sequence Analysis of the Antimicrobial Peptide Ctri9594 from the Venom of the Scorpion Chaerilus tricostatus
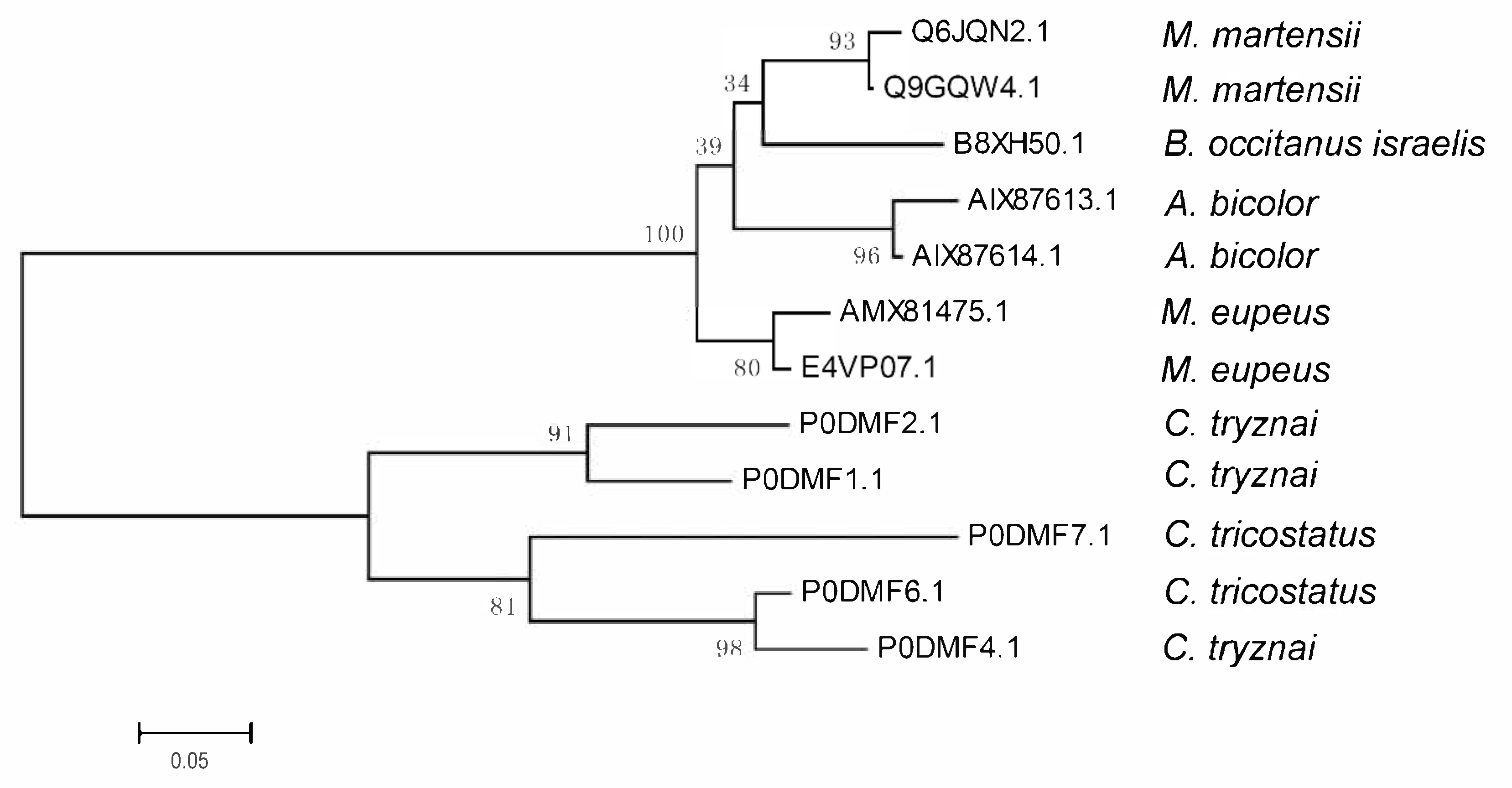
2.3. Comparison and Evolutionary Analysis of Ctri9594 and Its Homologs
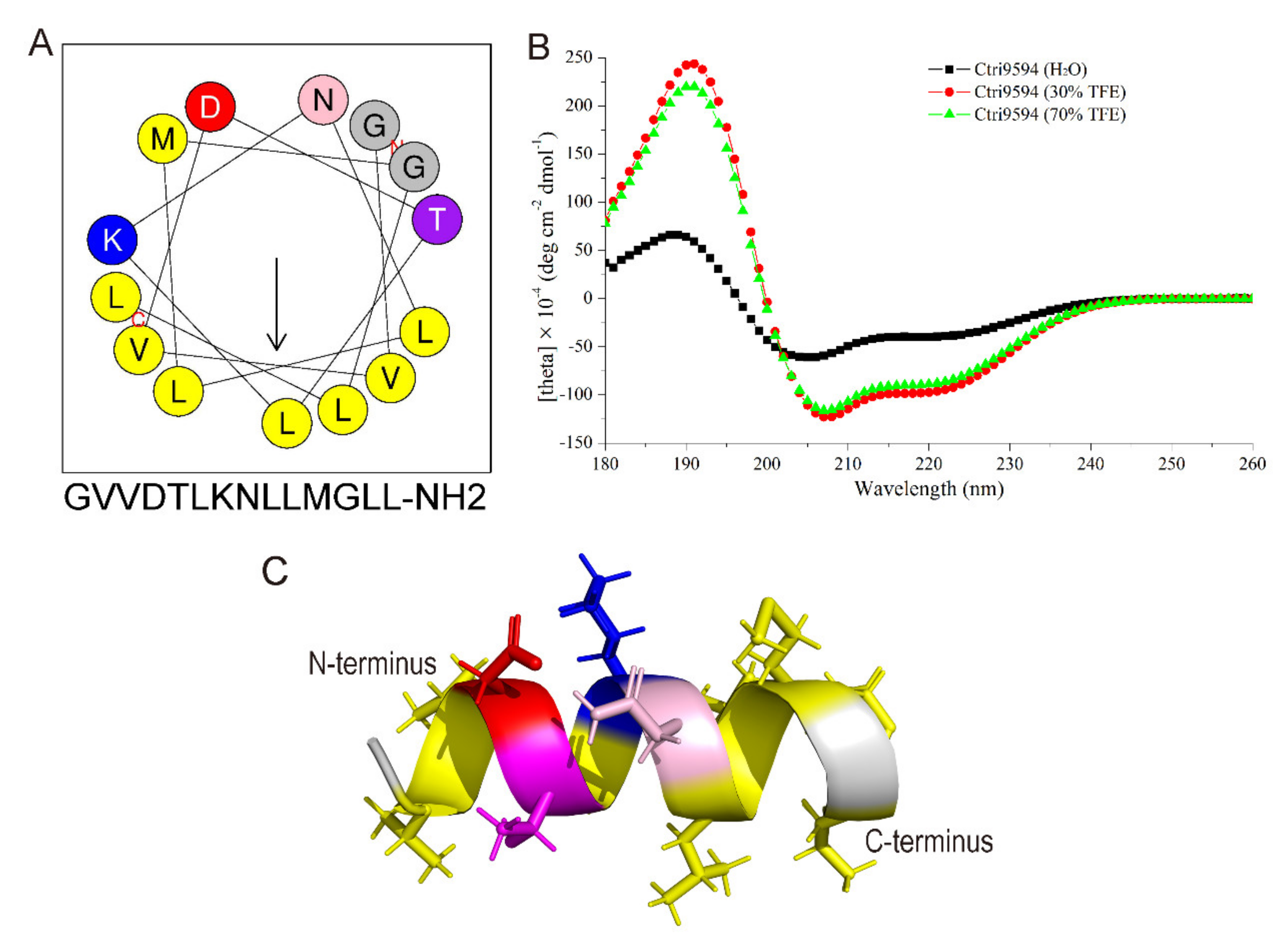
2.4. Structural Analysis of the Chemically Synthesized Ctri9594 Mature Peptide
2.5. Antimicrobial Activity of the Chemically Synthesized Ctri9594 Mature Peptide
3. Discussion
4. Materials and Methods
4.1. Screening cDNA Library with PCR Strategy
4.2. cDNA Sequencing and Sequence Analysis
4.3. Chemical Synthesis
4.4. Structure Prediction and CD Analysis
4.5. Antimicrobial Assay
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Furtado, A.A.; Daniele-Silva, A.; Silva-Júnior, A.A.D.; Fernandes-Pedrosa, M.F. Biology, venom composition, and scorpionism induced by brazilian scorpion Tityus stigmurus (Thorell, 1876) (Scorpiones: Buthidae): A mini-review. Toxicon 2020, 185, 36–45. [Google Scholar] [CrossRef]
- Dueñas-Cuellar, R.A.; Santana, C.J.C.; Magalhães, A.C.M.; Pires, O.R., Jr.; Fontes, W.; Castro, M.S. Scorpion toxins and ion channels: Potential applications in cancer therapy. Toxins 2020, 12, 326. [Google Scholar] [CrossRef]
- Yacoub, T.; Rima, M.; Karam, M.; Fajloun, J.S.A.Z. Antimicrobials from venomous animals: An overview. Molecules 2020, 25, 2402. [Google Scholar] [CrossRef] [PubMed]
- Conde, R.; Zamudio, F.Z.; Rodriguez, M.H.; Possani, L.D. Scorpine, an anti-malaria and anti-bacterial agent purified from scorpion venom. FEBS Lett. 2000, 471, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E.F.; Diego-Garcia, E.; Rodriguez de la Vega, R.C.; Possani, L.D. Transcriptome analysis of the venom gland of the Mexican scorpion Hadrurus gertschi (Arachnida: Scorpiones). BMC Genom. 2007, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Tytgat, J. The scorpine family of defensins: Gene structure, alternative polyadenylation and fold recognition. Cell Mol. Life Sci. 2004, 61, 1751–1763. [Google Scholar] [CrossRef]
- Zeng, X.C.; Corzo, G.; Hahin, R. Scorpion venom peptides without disulfide bridges. IUBMB Life 2005, 57, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.C.; Wang, S.X.; Zhu, Y.; Zhu, S.Y.; Li, W.X. Identification and functional characterization of novel scorpion venom peptides with no disulfide bridge from Buthus martensii Karsch. Peptides 2004, 25, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Yasuda, A.; Naoki, H.; Corzo, G.; Andriantsiferana, M.; Nakajima, T. IsCT, a novel cytotoxic linear peptide from scorpion Opisthacanthus madagascariensis. Biochem. Biophys. Res. Commun. 2001, 286, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; He, Y.; Zhao, R.; Wu, Y.; Li, W.; Cao, Z. Extreme diversity of scorpion venom peptides and proteins revealed by transcriptomic analysis: Implication for proteome evolution of scorpion venom arsenal. J. Proteom. 2012, 75, 1563–1576. [Google Scholar] [CrossRef]
- Lourenço, W.R.; Ythier, E.; Cloudsley-Thompson, J.L. Observations on the life history of Chaerilus philippinus Lourenço & Ythier, 2008 (Scorpiones, Chaerilidae) from the Philippines. Comptes Rendus Biol. 2008, 331, 896–900. [Google Scholar]
- He, Y.; Zhao, R.; Di, Z.; Li, Z.; Xu, X.; Hong, W.; Wu, Y.; Zhao, H.; Li, W.; Cao, Z. Molecular diversity of Chaerilidae venom peptides reveals the dynamic evolution of scorpion venom components from Buthidae to non-Buthidae. J. Proteom. 2013, 89, 1–14. [Google Scholar] [CrossRef]
- Fan, Z.; Cao, L.; He, Y.; Hu, J.; Di, Z.; Wu, Y.; Li, W.; Cao, Z. Ctriporin, a new anti-methicillin-resistant Staphylococcus aureus peptide from the venom of the scorpion Chaerilus tricostatus. Antimicrob. Agents Chemother. 2011, 55, 5220–5229. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Feng, J.; Yu, C.; Li, Z.; Wu, Y.; Cao, Z.; Li, W.; He, X.; Xiang, M.; Han, S. Identification of a new specific Kv1.3 channel blocker, Ctri9577, from the scorpion Chaerilus tricostatus. Peptides 2012, 36, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Chen, J.; Hao, J.; Zhang, J.; Huang, X.; Hu, F.; Wu, Z.; Liu, Y.; Li, W.; Cao, Z.; et al. Discovery of three toxin peptides with Kv1.3 channel and IL-2 cytokine-inhibiting activities from Non-Buthidae scorpions, Chaerilus tricostatus and Chaerilus tryznai. Peptides 2017, 91, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Douglass, J.; Civelli, O.; Herbert, E. Polyprotein gene expression: Generation of diversity of neuroendocrine peptides. Annu. Rev. Biochem. 1984, 53, 665–715. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, K.; Zhu, W.; Ye, X.; Ding, L.; Jiang, H.; Li, F.; Chen, Z.; Luo, X. Two new cationic α-helical peptides identified from the venom gland of Liocheles australasiae possess antimicrobial activity against methicillin-resistant staphylococci. Toxicon 2021, 196, 63–73. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Raja, Z.; Humblot, V.; Piesse, C.; Foulon, T.; Sereno, D.; Oury, B.; Ladram, A. Functional characterization of temporin-SHe, a new broad-spectrum antibacterial and Leishmanicidal temporin-SH paralog from the sahara frog (Pelophylax saharicus). Int. J. Mol. Sci. 2020, 21, 6713. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368, eaau5480. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E. Peptide antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
- Borah, A.; Deb, B.; Chakraborty, S. A crosstalk on antimicrobial peptides. Int. J. Pept. Res. Ther. 2021, 27, 229–244. [Google Scholar] [CrossRef]
- Juretić, D.; Simunić, J. Design of alpha-helical antimicrobial peptides with a high selectivity index. Expert Opin. Drug Discov. 2019, 14, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Berditsch, M.; Hawecker, J.; Ardakani, M.F.; Gerthsen, D.; Ulrich, A.S. Damage of the bacterial cell envelope by antimicrobial peptides gramicidin S and PGLa as revealed by transmission and scanning electron microscopy. Antimicrob. Agents Chemother. 2010, 54, 3132–3142. [Google Scholar] [CrossRef] [Green Version]
- Zelezetsky, I.; Tossi, A. Alpha-helical antimicrobial peptides--using a sequence template to guide structure-activity relationship studies. Biochim. Biophys. Acta. 2006, 1758, 1436–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.Y.; You, R.I.; Lai, M.J.; Lin, N.T.; Chen, L.K.; Chang, K.C. Highly potent antimicrobial modified peptides derived from the Acinetobacter baumannii phage endolysin LysAB2. Sci. Rep. 2017, 7, 11477. [Google Scholar] [CrossRef] [Green Version]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Liu, G.; Yang, F.; Li, F.; Li, Z.; Lang, Y.; Shen, B.; Wu, Y.; Li, W.; Harrison, P.L.; Strong, P.N.; et al. Therapeutic potential of a scorpion venom-derived antimicrobial peptide and its homologs against antibiotic-resistant gram-positive bacteria. Front. Microbiol. 2018, 9, 1159. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (μg/mL) | |||
|---|---|---|---|---|
| Ctri9594 | Amp | Kan | Amb | |
| Gram-positive bacteria | ||||
| Bacillus thuringensis AB92037 | 25 | >100 | 4 | ND |
| Bacillus subtilis AB91021 | 25 | >100 | 20 | ND |
| Staphylococcus aureus AB94004 | 25 | 2 | 50 | ND |
| Micrococcus luteus AB93113 | 12.5 | 10 | 20 | ND |
| Gram-negative bacteria | ||||
| Escherichia coli AB94012 | >100 | ND | 60 | ND |
| Pseudomonas aeruginosa AB93066 | >100 | ND | 60 | ND |
| Fungus | ||||
| Candida albicans AY93025 | >100 | ND | ND | 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, D.; Cao, Z.; Zhang, R.; Li, W. Molecular Cloning and Functional Identification of the Antimicrobial Peptide Gene Ctri9594 from the Venom of the Scorpion Chaerilus tricostatus. Antibiotics 2021, 10, 896. https://doi.org/10.3390/antibiotics10080896
He D, Cao Z, Zhang R, Li W. Molecular Cloning and Functional Identification of the Antimicrobial Peptide Gene Ctri9594 from the Venom of the Scorpion Chaerilus tricostatus. Antibiotics. 2021; 10(8):896. https://doi.org/10.3390/antibiotics10080896
Chicago/Turabian StyleHe, Dangui, Zhijian Cao, Ruhong Zhang, and Wenhua Li. 2021. "Molecular Cloning and Functional Identification of the Antimicrobial Peptide Gene Ctri9594 from the Venom of the Scorpion Chaerilus tricostatus" Antibiotics 10, no. 8: 896. https://doi.org/10.3390/antibiotics10080896