
The Small Metal-Binding Protein SmbP Simplifies the Recombinant Expression and Purification of the Antimicrobial Peptide LL-37

 , and
, and 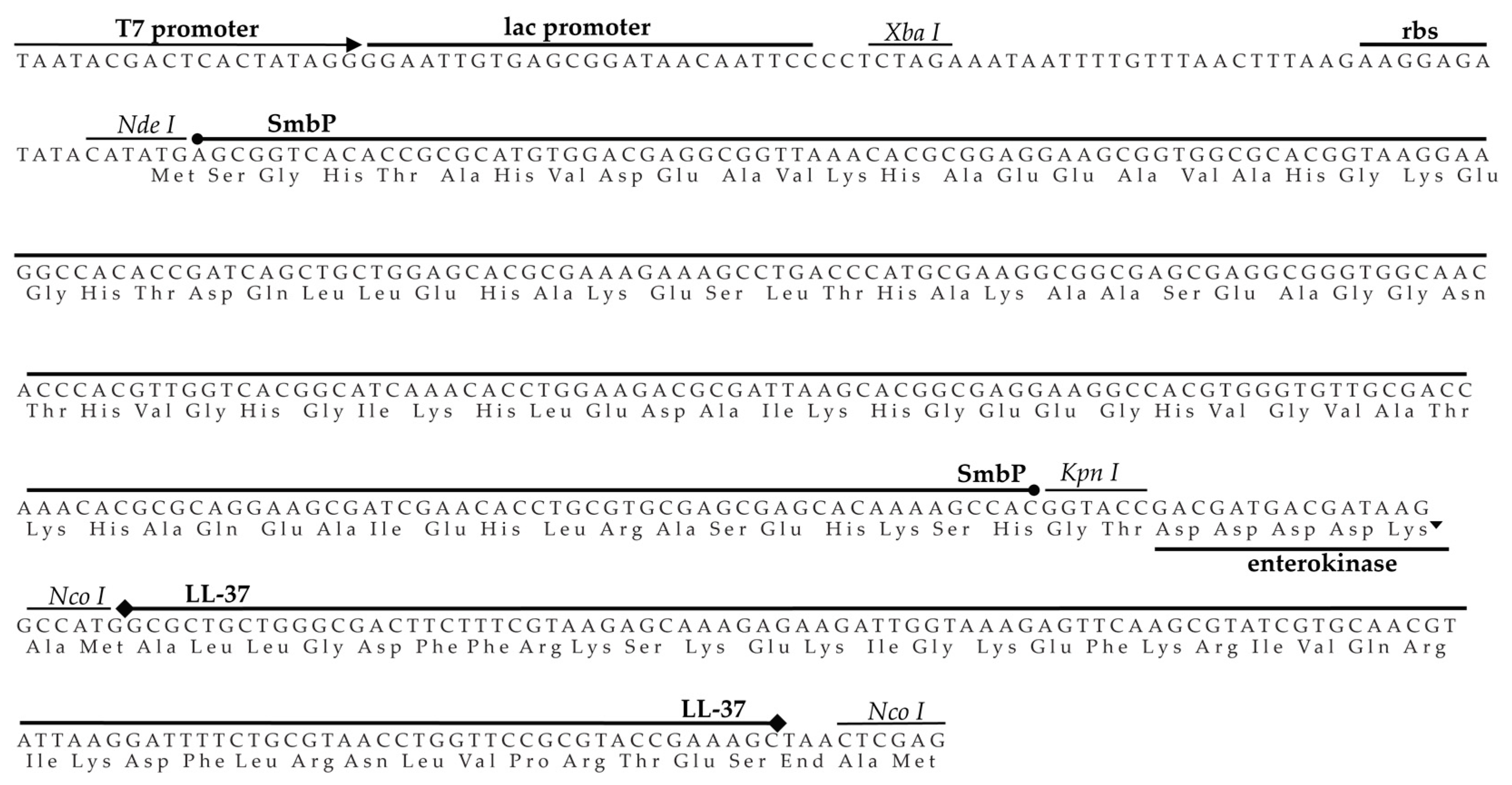{kind=link}
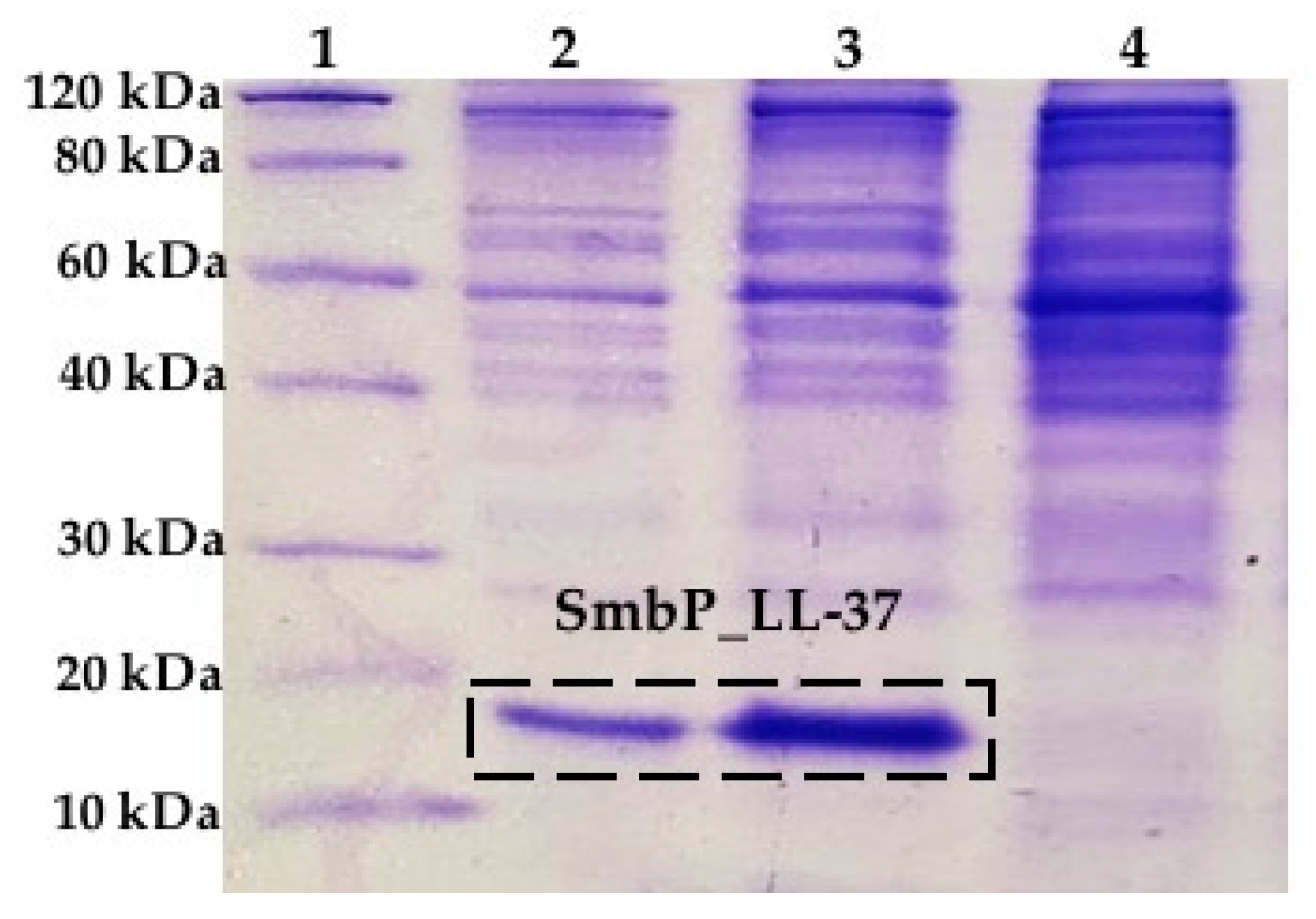{kind=link}
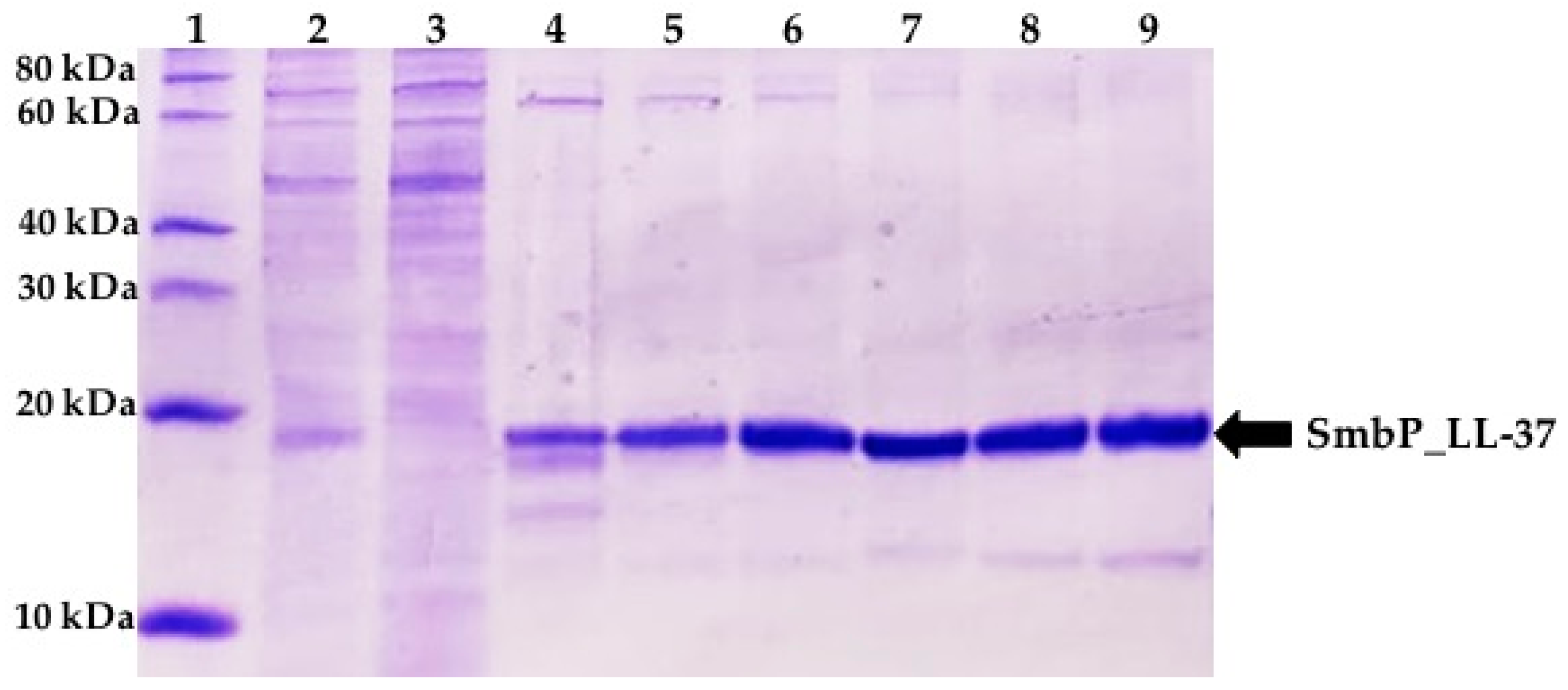{kind=link}
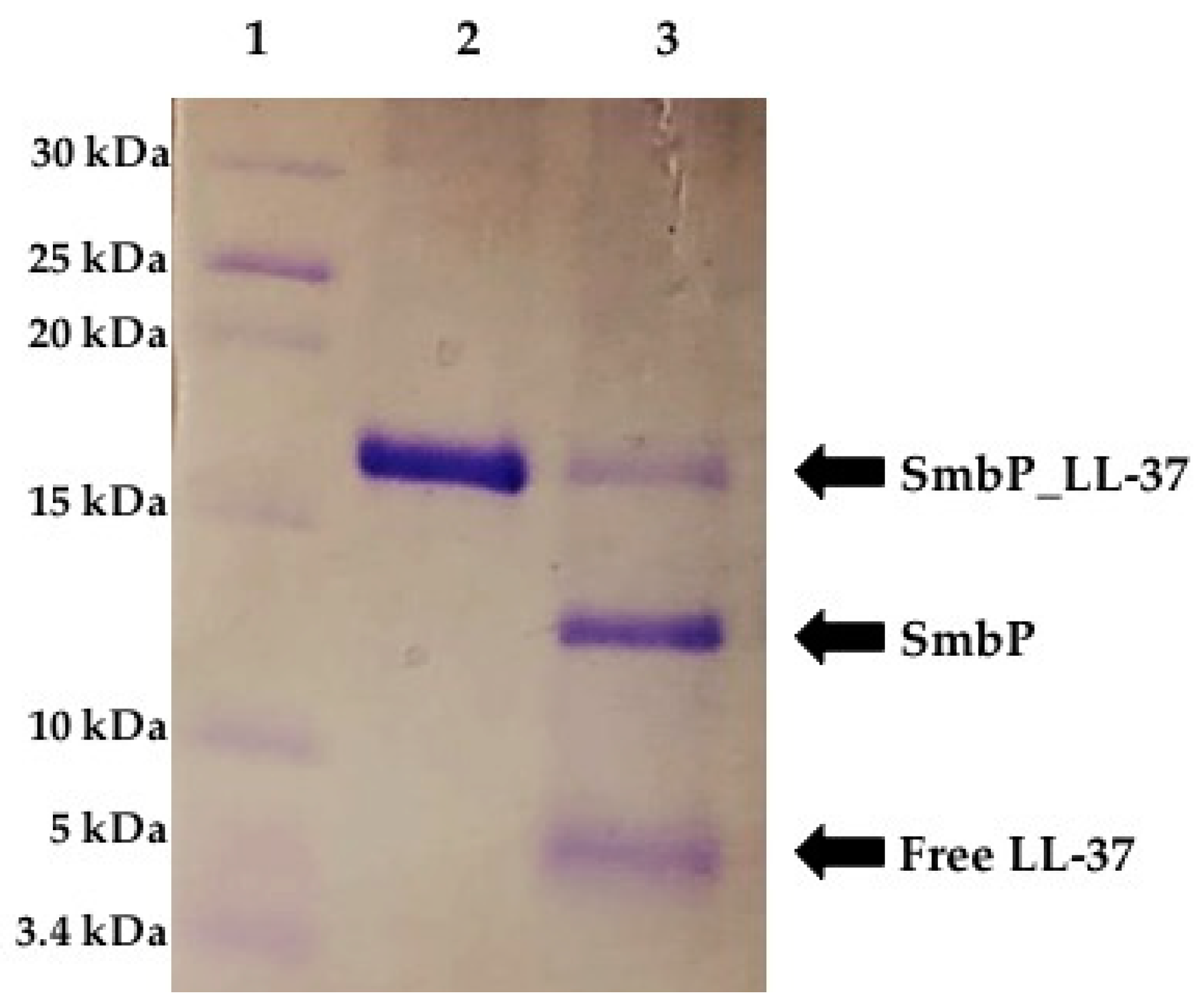{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
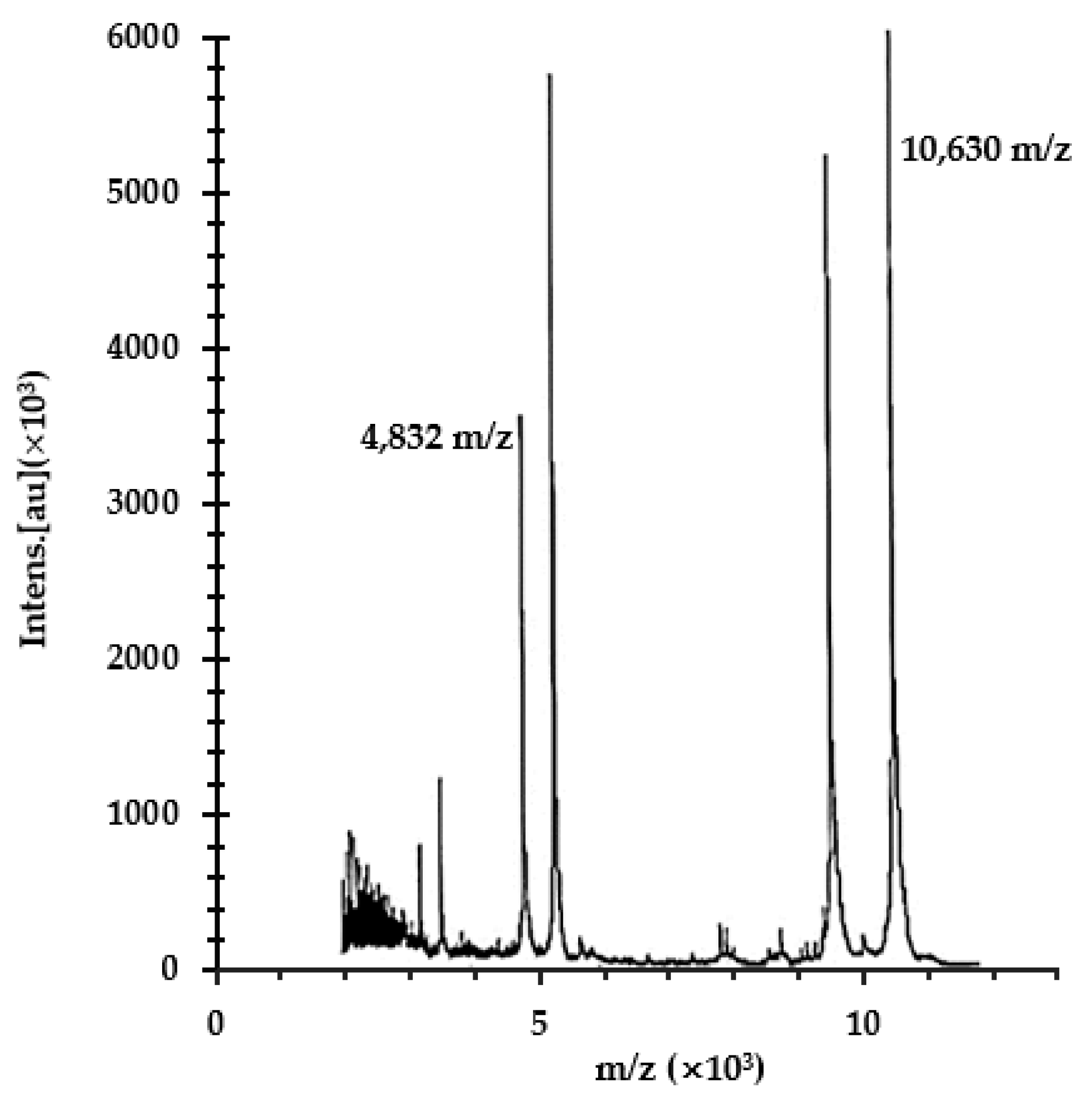
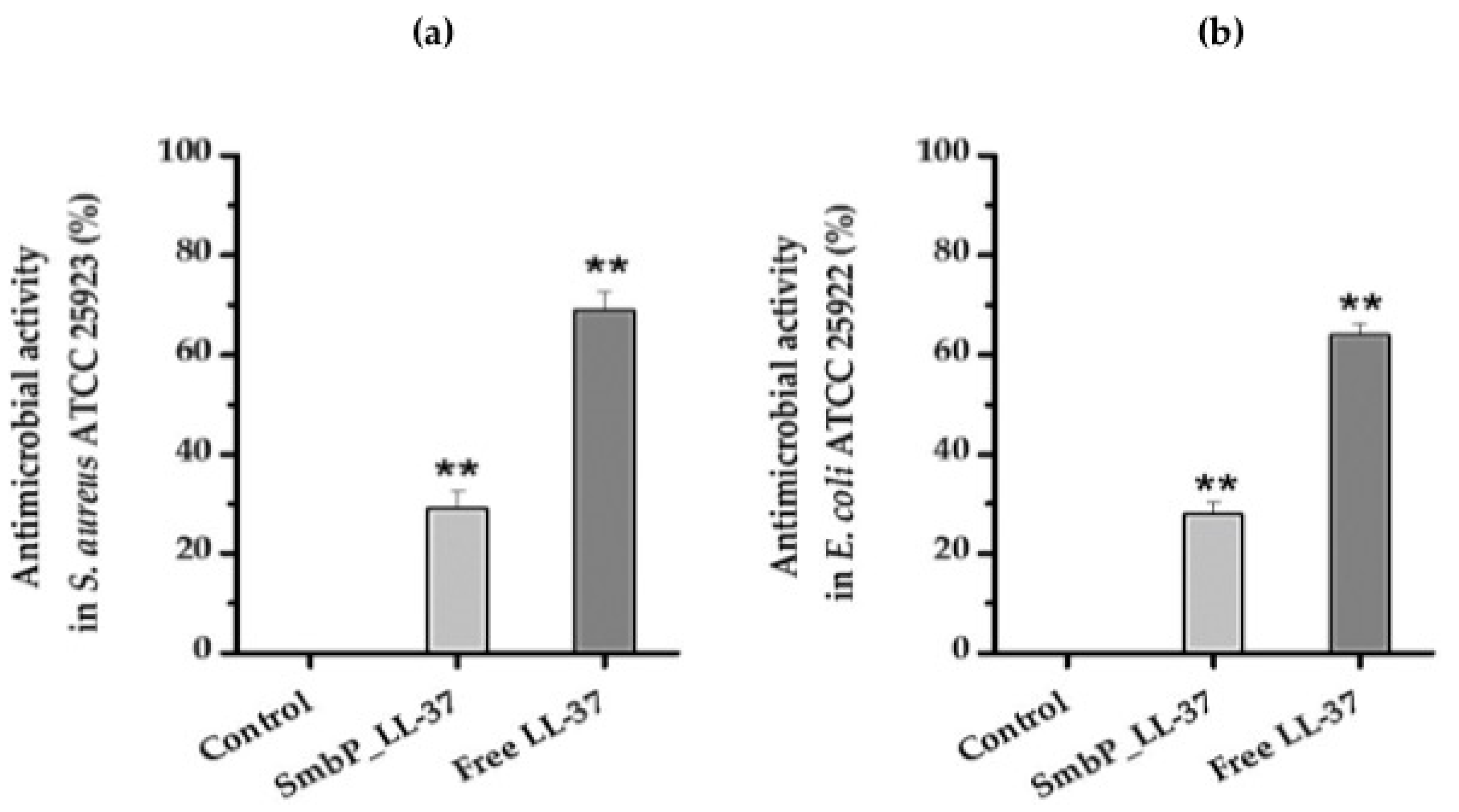
2. Results and Discussion
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barney, B.M.; LoBrutto, R.; Francisco, W.A. Characterization of a small metal binding protein from Nitrosomonas europaea. Biochemistry 2004, 43, 11206–11213. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Cortez, T.; Morones-Ramirez, J.R.; Balderas-Renteria, I.; Zarate, X. Expression and purification of recombinant proteins in Escherichia coli tagged with a small metal-binding protein from Nitrosomonas europaea. Protein Expr. Purif. 2016, 118, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.D.; Morones-Ramirez, J.R.; Balderas-Renteria, I.; Casillas-Vega, N.G.; Galbraith, D.W.; Zarate, X. Optimizing Periplasmic Expression in Escherichia coli for the Production of Recombinant Proteins Tagged with the Small Metal-Binding Protein SmbP. Mol. Biotechnol. 2019, 61, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Perez-Perez, D.A.; Pioquinto-Avila, E.; Arredondo-Espinoza, E.; Morones-Ramirez, J.R.; Balderas-Renteria, I.; Zarate, X. Engineered small metal-binding protein tag improves the production of recombinant human growth hormone in the periplasm of Escherichia coli. FEBS Open Bio 2020, 10, 546–551. [Google Scholar] [CrossRef] [Green Version]
- Montfort-Gardeazabal, J.M.; Claudio, P.C.M.-S.; Casillas-Vega, N.G.; Zarate, X. Expression and Purification of the VpDef Defensin in Escherichia coli using the Small Metal-Binding Proteins CusF3H+ and SmbP. Protein Pept. Lett. 2020, 28, 108–114. [Google Scholar] [CrossRef]
- Montfort-Gardeazabal, J.M.; Balderas-Renteria, I.; Casillas-Vega, N.G.; Zarate, X. Expression and purification of the antimicrobial peptide Bin1b in Escherichia coli tagged with the fusion proteins CusF3H+ and SmbP. Protein Expr. Purif. 2021, 178, 105784. [Google Scholar] [CrossRef]
- Nizet, V.; Ohtake, T.; Lauth, X.; Trowbridge, J.; Rudisill, J.; Dorschner, R.A.; Pestonjamasp, V.; Piraino, J.; Huttner, K.; Gallo, R.L. Innate antimicrobial peptide protects the skin from invasive bacterial infection. Nature 2001, 414, 454–457. [Google Scholar] [CrossRef]
- Chromek, M.; Slamová, Z.; Bergman, P.; Kovács, L.; Podracká, L.; Ehrén, I.; Hökfelt, T.; Gudmundsson, G.H.; Gallo, R.L.; Agerberth, B.; et al. The antimicrobial peptide cathelicidin protects the urinary tract against invasive bacterial infection. Nat. Med. 2006, 12, 636–641. [Google Scholar] [CrossRef]
- Kang, J.; Dietz, M.J.; Li, B. Antimicrobial peptide LL-37 is bactericidal against Staphylococcus aureus biofilms. PLoS ONE 2019, 14, e0216676. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; McLean, D.T.F.; Linden, G.J.; McAuley, D.F.; McMullan, R.; Lundy, F.T. The naturally occurring host defense peptide, LL-37, and its truncated mimetics KE-18 and KR-12 have selected biocidal and antibiofilm activities against Candida albicans, Staphylococcus aureus, and Escherichia coli in vitro. Front. Microbiol. 2017, 8, 544. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Watson, K.M.; Buckheit, R.W. Anti-human immunodeficiency virus type 1 activities of antimicrobial peptides derived from human and bovine cathelicidins. Antimicrob. Agents Chemother. 2008, 52, 3438–3440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wetering, S.; Mannesse-Lazeroms, S.P.; Van Sterkenburg, M.A.; Daha, M.R.; Dijkman, J.H.; Hiemstra, P.S. Effect of defensins on interleukin-8 synthesis in airway epithelial cells. Am. J. Physiol. 1997, 272, L888–L896. [Google Scholar] [CrossRef]
- Hase, K.; Murakami, M.; Iimura, M.; Cole, S.P.; Horibe, Y.; Ohtake, T.; Obonyo, M.; Gallo, R.L.; Eckmann, L.; Kagnoff, M.F. Expression of LL-37 by human gastric epithelial cells as a potential host defense mechanism against Helicobacter pylori. Gastroenterology 2003, 125, 1613–1625. [Google Scholar] [CrossRef]
- Vandamme, D.; Landuyt, B.; Luyten, W.; Schoofs, L. A comprehensive summary of LL-37, the factoctum human cathelicidin peptide. Cell. Immunol. 2012, 280, 22–35. [Google Scholar] [CrossRef]
- Scheper, H.; Wubbolts, J.M.; Verhagen, J.A.M.; de Visser, A.W.; van der Wal, R.J.P.; Visser, L.G.; de Boer, M.G.J.; Nibbering, P.H. SAAP-148 Eradicates MRSA Persisters Within Mature Biofilm Models Simulating Prosthetic Joint Infection. Front. Microbiol. 2021, 12, 99. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Wang, G. Cloning, expression, isotope labeling, and purification of human antimicrobial peptide LL-37 in Escherichia coli for NMR studies. Protein Expr. Purif. 2006, 47, 498–505. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Li, H.; Lockridge, O.; Wang, G. A novel method for purifying recombinant human host defense cathelicidin LL-37 by utilizing its inherent property of aggregation. Protein Expr. Purif. 2007, 54, 157–165. [Google Scholar] [CrossRef]
- Ramos, R.; Domingues, L.; Gama, M. Escherichia coli expression and purification of LL37 fused to a family III carbohydrate-binding module from Clostridium thermocellum. Protein Expr. Purif. 2010, 71, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.Y.; Henzler-Wildman, K.A.; Ramamoorthy, A. Expression and purification of a recombinant LL-37 from Escherichia coli. Biochim. Biophys. Acta Biomembr. 2006, 1758, 1351–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y. Recombinant production of antimicrobial peptides in Escherichia coli: A review. Protein Expr. Purif. 2011, 80, 260–267. [Google Scholar] [CrossRef]
- Deng, T.; Ge, H.; He, H.; Liu, Y.; Zhai, C.; Feng, L.; Yi, L. The heterologous expression strategies of antimicrobial peptides in microbial systems. Protein Expr. Purif. 2017, 140, 52–59. [Google Scholar] [CrossRef]
- Liu, W.; Dong, S.L.; Xu, F.; Wang, X.Q.; Withers, T.R.; Yu, H.D.; Wang, X. Effect of intracellular expression of antimicrobial peptide LL-37 on growth of Escherichia coli strain TOP10 under aerobic and anaerobic conditions. Antimicrob. Agents Chemother. 2013, 57, 4707–4716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Lugo, J.J.; Santos, B.D.; Perez-Perez, D.A.; Montfort-Gardeazabal, J.M.; McEvoy, M.M.; Zarate, X. Expression and Purification of Recombinant Proteins in Escherichia coli Tagged with the Metal-Binding Proteins SmbP and CusF3H+ BT—Protein Downstream Processing: Design, Development, and Application of High and Low-Resolution Method; Labrou, N.E., Ed.; Springer: New York, NY, USA, 2021; pp. 329–344. ISBN 978-1-0716-0775-6. [Google Scholar]
- Li, Y. A novel protocol for the production of recombinant LL-37 expressed as a thioredoxin fusion protein. Protein Expr. Purif. 2012, 81, 201–210. [Google Scholar] [CrossRef]
- Rasigade, J.-P.; Vandenesch, F. Staphylococcus aureus: A pathogen with still unresolved issues. Infect. Genet. Evol. 2014, 21, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Braz, V.S.; Melchior, K.; Moreira, C.G. Escherichia coli as a Multifaceted Pathogenic and Versatile Bacterium. Front. Cell. Infect. Microbiol. 2020, 10, 548492. [Google Scholar] [CrossRef] [PubMed]
- Bommarius, B.; Jenssen, H.; Elliott, M.; Kindrachuk, J.; Pasupuleti, M.; Gieren, H.; Jaeger, K.-E.; Hancock, R.E.W.; Kalman, D. Cost-effective expression and purification of antimicrobial and host defense peptides in Escherichia coli. Peptides 2010, 31, 1957–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y. Production of human antimicrobial peptide LL-37 in Escherichia coli using a thioredoxin–SUMO dual fusion system. Protein Expr. Purif. 2013, 87, 72–78. [Google Scholar] [CrossRef]
- Guo, C.; Diao, H.; Lian, Y.; Yu, H.; Gao, H.; Zhang, Y.; Lin, D. Recombinant expression and characterization of an epididymis-specific antimicrobial peptide BIN1b/SPAG11E. J. Biotechnol. 2009, 139, 33–37. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez-Perez, D.A.; Villanueva-Ramirez, T.d.J.; Hernandez-Pedraza, A.E.; Casillas-Vega, N.G.; Gonzalez-Barranco, P.; Zarate, X. The Small Metal-Binding Protein SmbP Simplifies the Recombinant Expression and Purification of the Antimicrobial Peptide LL-37. Antibiotics 2021, 10, 1271. https://doi.org/10.3390/antibiotics10101271
Perez-Perez DA, Villanueva-Ramirez TdJ, Hernandez-Pedraza AE, Casillas-Vega NG, Gonzalez-Barranco P, Zarate X. The Small Metal-Binding Protein SmbP Simplifies the Recombinant Expression and Purification of the Antimicrobial Peptide LL-37. Antibiotics. 2021; 10(10):1271. https://doi.org/10.3390/antibiotics10101271
Chicago/Turabian StylePerez-Perez, David A., Teresa de J. Villanueva-Ramirez, Adriana E. Hernandez-Pedraza, Nestor G. Casillas-Vega, Patricia Gonzalez-Barranco, and Xristo Zarate. 2021. "The Small Metal-Binding Protein SmbP Simplifies the Recombinant Expression and Purification of the Antimicrobial Peptide LL-37" Antibiotics 10, no. 10: 1271. https://doi.org/10.3390/antibiotics10101271