Charge-Sensitive Optical Detection of Binding Kinetics between Phage-Displayed Peptide Ligands and Protein Targets



Abstract
:1. Introduction
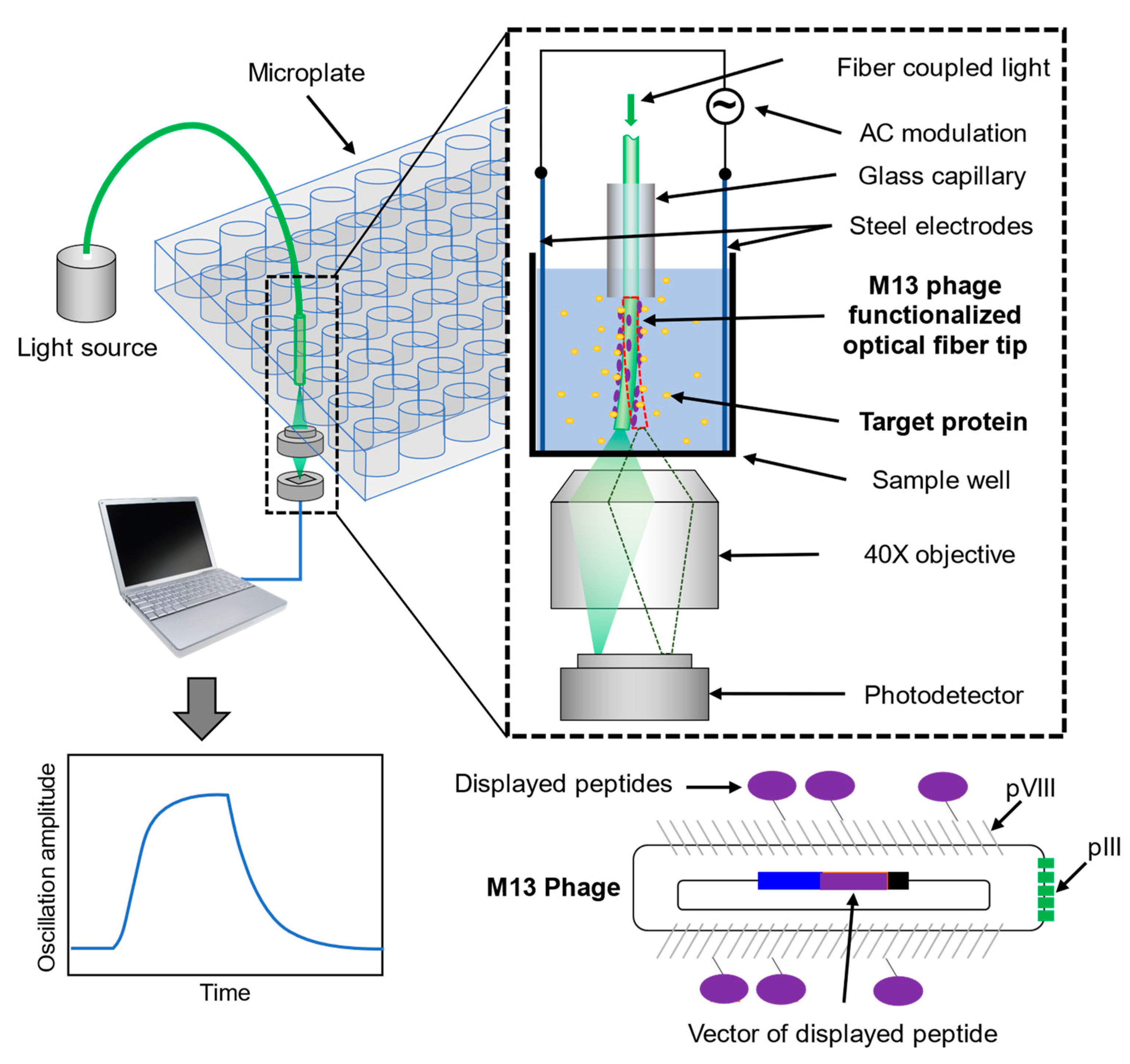
2. Detection Principle
3. Materials and Methods
3.1. Materials
3.2. Generation of M13 Phage Displayed Peptides
3.3. Charge-Sensitive Optical Detection Setup
3.4. Surface Functionalization
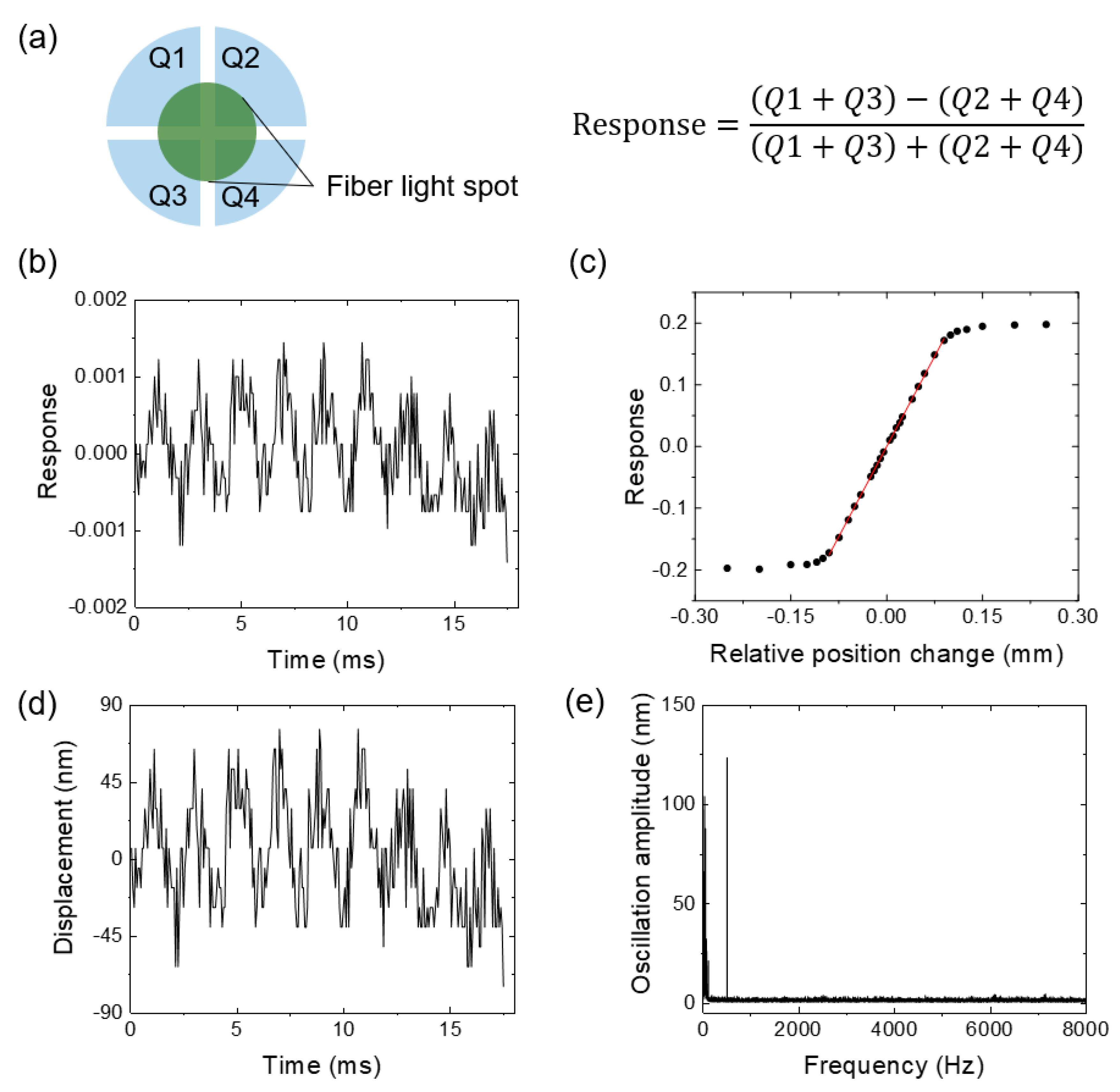
3.5. Calibration Curve
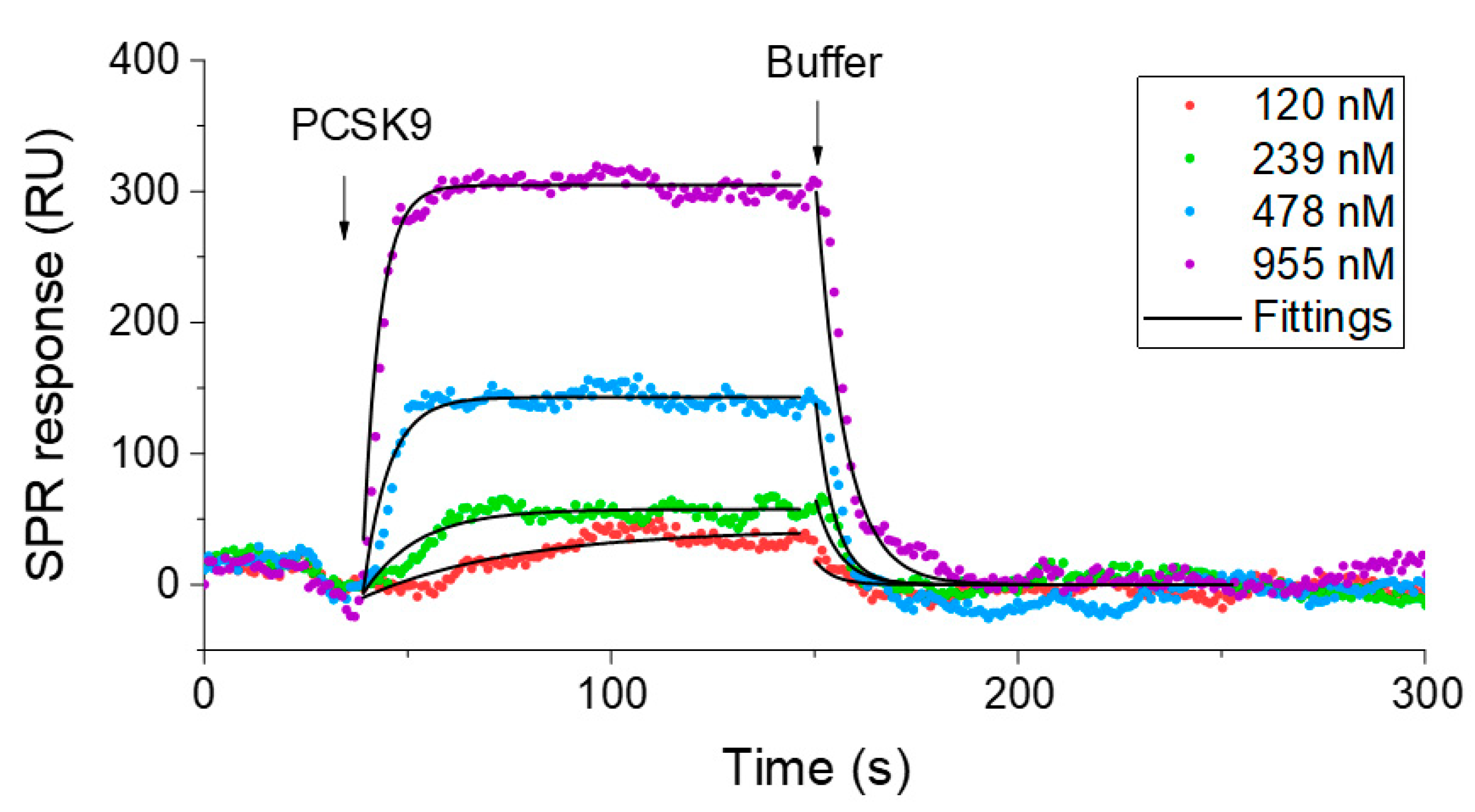
3.6. SPR Measurement
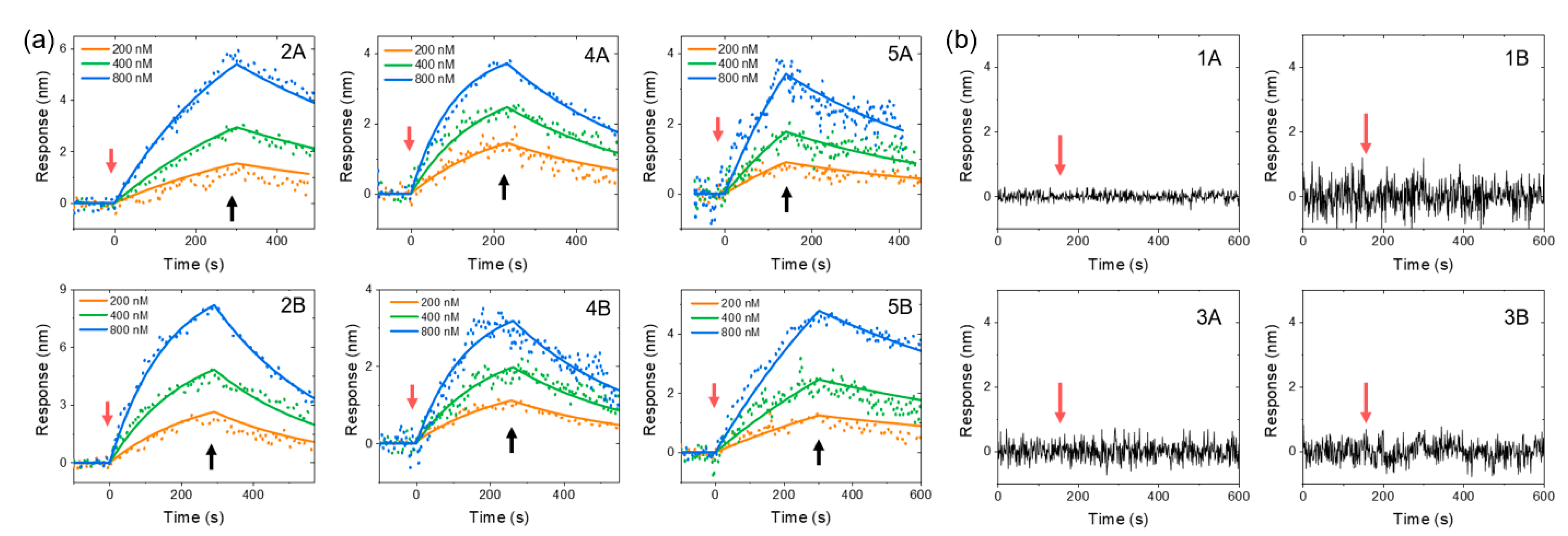
4. Results and Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Zhang, H.; Ghosh, D.; Williams, R.O. Just how prevalent are peptide therapeutic products? A critical review. Int. J. Pharm. 2020, 587, 119491. [Google Scholar] [CrossRef] [PubMed]
- Vlieghe, P.; Lisowski, V.; Martinez, J.; Khrestchatisky, M. Synthetic therapeutic peptides: Science and market. Drug Discov. Today 2010, 15, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, J.; Marasco, W.A. Phage and Yeast Display. Microbiol. Spectr. 2015, 3, AID-0028-2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonikian, R.; Zhang, Y.; Boone, C.; Sidhu, S.S. Identifying specificity profiles for peptide recognition modules from phage-displayed peptide libraries. Nat. Protoc. 2007, 2, 1368–1386. [Google Scholar] [CrossRef] [PubMed]
- Plikusiene, I.; Balevicius, Z.; Ramanaviciene, A.; Talbot, J.; Mickiene, G.; Balevicius, S.; Stirke, A.; Tereshchenko, A.; Tamosaitis, L.; Zvirblis, G.; et al. Evaluation of affinity sensor response kinetics towards dimeric ligands linked with spacers of different rigidity: Immobilized recombinant granulocyte colony-stimulating factor based synthetic receptor binding with genetically engineered dimeric analyte derivatives. Biosens. Bioelectron. 2020, 156, 112112. [Google Scholar] [PubMed]
- Hunt, A.C.; Case, J.B.; Park, Y.-J.; Cao, L.; Wu, K.; Walls, A.C.; Liu, Z.; Bowen, J.E.; Yeh, H.-W.; Saini, S.; et al. Multivalent designed proteins neutralize SARS-CoV-2 variants of concern and confer protection against infection in mice. Sci. Transl. Med. 2022, 14, eabn1252. [Google Scholar] [CrossRef] [PubMed]
- Homola, J.; Yee, S.S.; Gauglitz, G. Surface plasmon resonance sensors: Review. Sens. Actuators B Chem. 1999, 54, 3–15. [Google Scholar] [CrossRef]
- Wartchow, C.A.; Podlaski, F.; Li, S.; Rowan, K.; Zhang, X.; Mark, D.; Huang, K.S. Biosensor-based small molecule fragment screening with biolayer interferometry. J. Comput.-Aided Mol. Des. 2011, 25, 669–676. [Google Scholar] [CrossRef]
- Lunder, M.; Bratkovič, T.; Anderluh, G.; Štrukelj, B.; Kreft, S. Affinity ranking of phage-displayed peptides: Enzyme-linked immunosorbent assay versus surface plasmon resonance. Acta Chim. Slov. 2008, 55, 233–235. [Google Scholar]
- Lasonder, E.; Schellekens, G.A.; Welling, G.W. A fast and sensitive method for the evaluation of binding of phage clones selected from a surface displayed library. Nucleic Acids Res. 1994, 22, 545–546. [Google Scholar] [CrossRef] [Green Version]
- Shabani, A.; Zourob, M.; Allain, B.; Marquette, C.A.; Lawrence, M.F.; Mandeville, R. Bacteriophage-modified microarrays for the direct impedimetric detection of bacteria. Anal. Chem. 2008, 80, 9475–9482. [Google Scholar] [CrossRef] [PubMed]
- Tolba, M.; Ahmed, M.U.; Tlili, C.; Eichenseher, F.; Loessner, M.J.; Zourob, M. A bacteriophage endolysin-based electrochemical impedance biosensor for the rapid detection of Listeria cells. Analyst 2012, 137, 5749–5756. [Google Scholar] [CrossRef] [PubMed]
- Tlili, C.; Sokullu, E.; Safavieh, M.; Tolba, M.; Ahmed, M.U.; Zourob, M. Bacteria Screening, Viability, And Confirmation Assays Using Bacteriophage-Impedimetric/Loop-Mediated Isothermal Amplification Dual-Response Biosensors. Anal. Chem. 2013, 85, 4893–4901. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Shan, X.; Wang, S.; Zhang, P.; Tao, N. Detection of molecular binding via charge-induced mechanical response of optical fibers. Chem. Sci. 2014, 5, 4375–4381. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Guan, Y.; Wang, S.; Xu, H.; Tao, N. Study of Small-Molecule-Membrane Protein Binding Kinetics with Nanodisc and Charge-Sensitive Optical Detection. Anal. Chem. 2016, 88, 2375–2379. [Google Scholar] [CrossRef] [Green Version]
- Liang, R.; Ma, G.; Jing, W.; Wang, Y.; Yang, Y.; Tao, N.; Wang, S. Charge-Sensitive Optical Detection of Small Molecule Binding Kinetics in Normal Ionic Strength Buffer. ACS Sens. 2020, 6, 364–370. [Google Scholar] [CrossRef]
- Zhang, Y.; Ultsch, M.; Skelton, N.J.; Burdick, D.J.; Beresini, M.H.; Li, W.; Kong-Beltran, M.; Peterson, A.; Quinn, J.; Chiu, C.; et al. Discovery of a cryptic peptide-binding site on PCSK9 and design of antagonists. Nat. Struct. Mol. Biol. 2017, 24, 848–856. [Google Scholar] [CrossRef]
- Kunkel, T.A.; Roberts, J.D.; Zakour, R.A. Rapid and efficient site-specific mutagenesis without phenotypic selection. Methods Enzym. 1987, 154, 367–382. [Google Scholar]
- Costet, P.; Krempf, M.; Cariou, B. PCSK9 and LDL cholesterol: Unravelling the target to design the bullet. Trends Biochem. Sci. 2008, 33, 426–434. [Google Scholar] [CrossRef]
- Horton, J.D.; Cohen, J.C.; Hobbs, H.H. PCSK9: A convertase that coordinates LDL catabolism. J. Lipid Res. 2009, 50, S172–S177. [Google Scholar] [CrossRef] [Green Version]
- Seidah, N.G. PCSK9 as a therapeutic target of dyslipidemia. Expert Opin. Targets 2009, 13, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.W.; Lagace, T.A.; Garuti, R.; Zhao, Z.; McDonald, M.; Horton, J.D.; Cohen, J.C.; Hobbs, H.H. Binding of proprotein convertase subtilisin/kexin type 9 to epidermal growth factor-like repeat A of low density lipoprotein receptor decreases receptor recycling and increases degradation. J. Biol. Chem. 2007, 282, 18602–18612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Eigenbrot, C.; Zhou, L.; Shia, S.; Li, W.; Quan, C.; Tom, J.; Moran, P.; Di Lello, P.; Skelton, N.J.; et al. Identification of a small peptide that inhibits PCSK9 protein binding to the low density lipoprotein receptor. J. Biol. Chem. 2014, 289, 942–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Displayed Peptide Sequence | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample ID | Sample Name | Note | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | Signal Peptide | ||
| 1A, 1B | no SG pep2-8AA-2202 | negative control | T | V | A | T | S | A | E | E | Y | L | D | W | V | MKIKTGARILALSALTTMMFSASAYA | ||
| 2A, 2B | no SG pep2-8-2202 | positive | T | V | F | T | S | W | E | E | Y | L | D | W | V | MKIKTGARILALSALTTMMFSASAYA | ||
| 3A, 3B | no SG pep2-8AA-2478 | negative control | T | V | A | T | S | A | E | E | Y | L | D | W | V | MKKNIAFLLASMFVFSIATNAYA | ||
| 4A, 4B | no SG pep2-8-2478 | positive | T | V | F | T | S | W | E | E | Y | L | D | W | V | MKKNIAFLLASMFVFSIATNAYA | ||
| 5A, 5B | SG pep2-8-2478 | positive | S | G | T | V | F | T | S | W | E | E | Y | L | D | W | V | MKKNIAFLLASMFVFSIATNAYA |
| Sample | Description | |||
|---|---|---|---|---|
| 2A | no SG pep2-8-2202 | |||
| 2B | no SG pep2-8-2202 | |||
| 4A | no SG pep2-8-2478 | |||
| 4B | no SG pep2-8-2478 | |||
| 5A | SG pep2-8-2478 | |||
| 5B | SG pep2-8-2478 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, R.; Zhang, Y.; Ma, G.; Wang, S. Charge-Sensitive Optical Detection of Binding Kinetics between Phage-Displayed Peptide Ligands and Protein Targets. Biosensors 2022, 12, 394. https://doi.org/10.3390/bios12060394
Liang R, Zhang Y, Ma G, Wang S. Charge-Sensitive Optical Detection of Binding Kinetics between Phage-Displayed Peptide Ligands and Protein Targets. Biosensors. 2022; 12(6):394. https://doi.org/10.3390/bios12060394
Chicago/Turabian StyleLiang, Runli, Yingnan Zhang, Guangzhong Ma, and Shaopeng Wang. 2022. "Charge-Sensitive Optical Detection of Binding Kinetics between Phage-Displayed Peptide Ligands and Protein Targets" Biosensors 12, no. 6: 394. https://doi.org/10.3390/bios12060394