Advances in the Detection of Toxic Algae Using Electrochemical Biosensors

 and
and 
Abstract
:
1. Introduction
2. Material and Methods
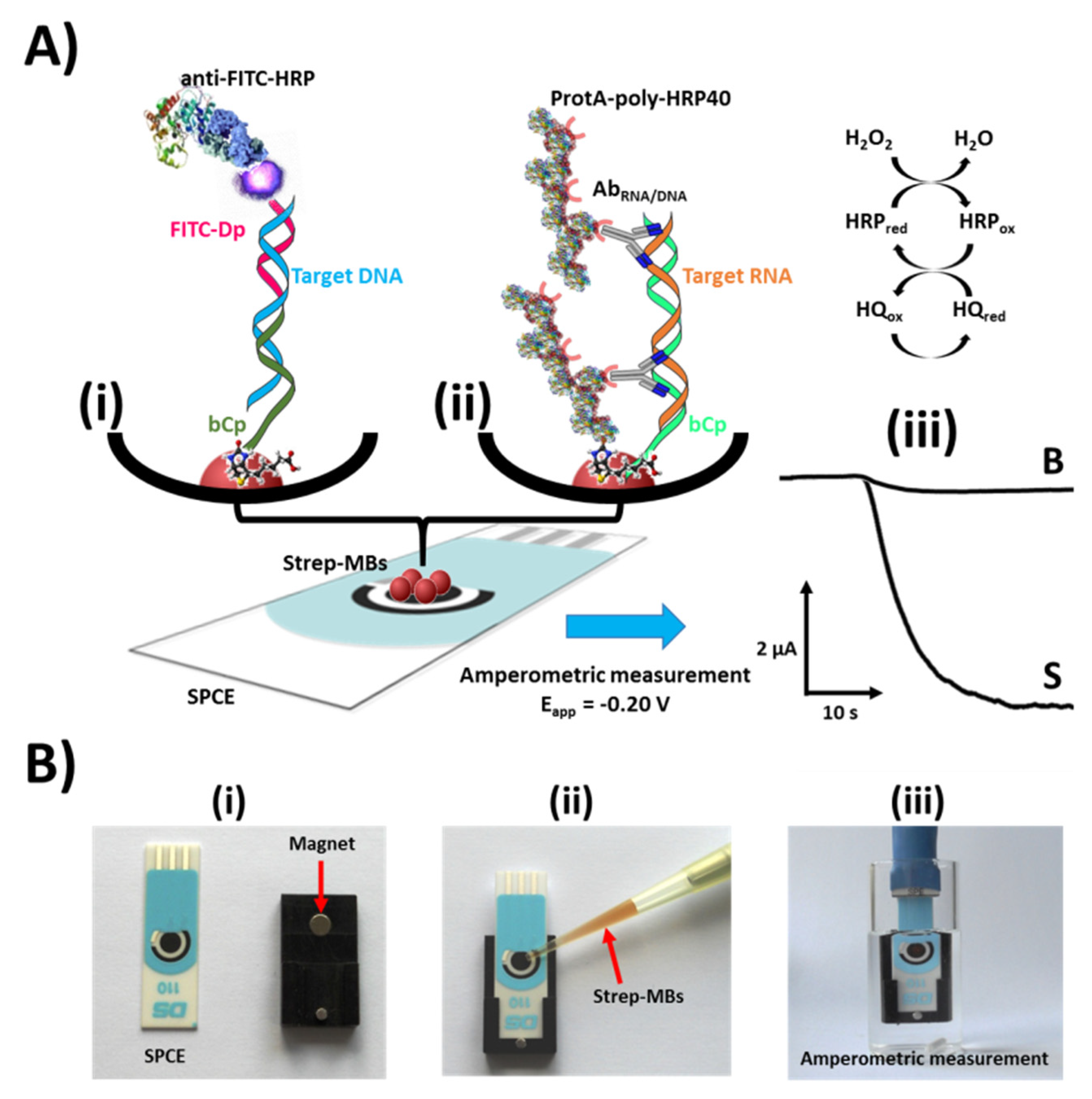
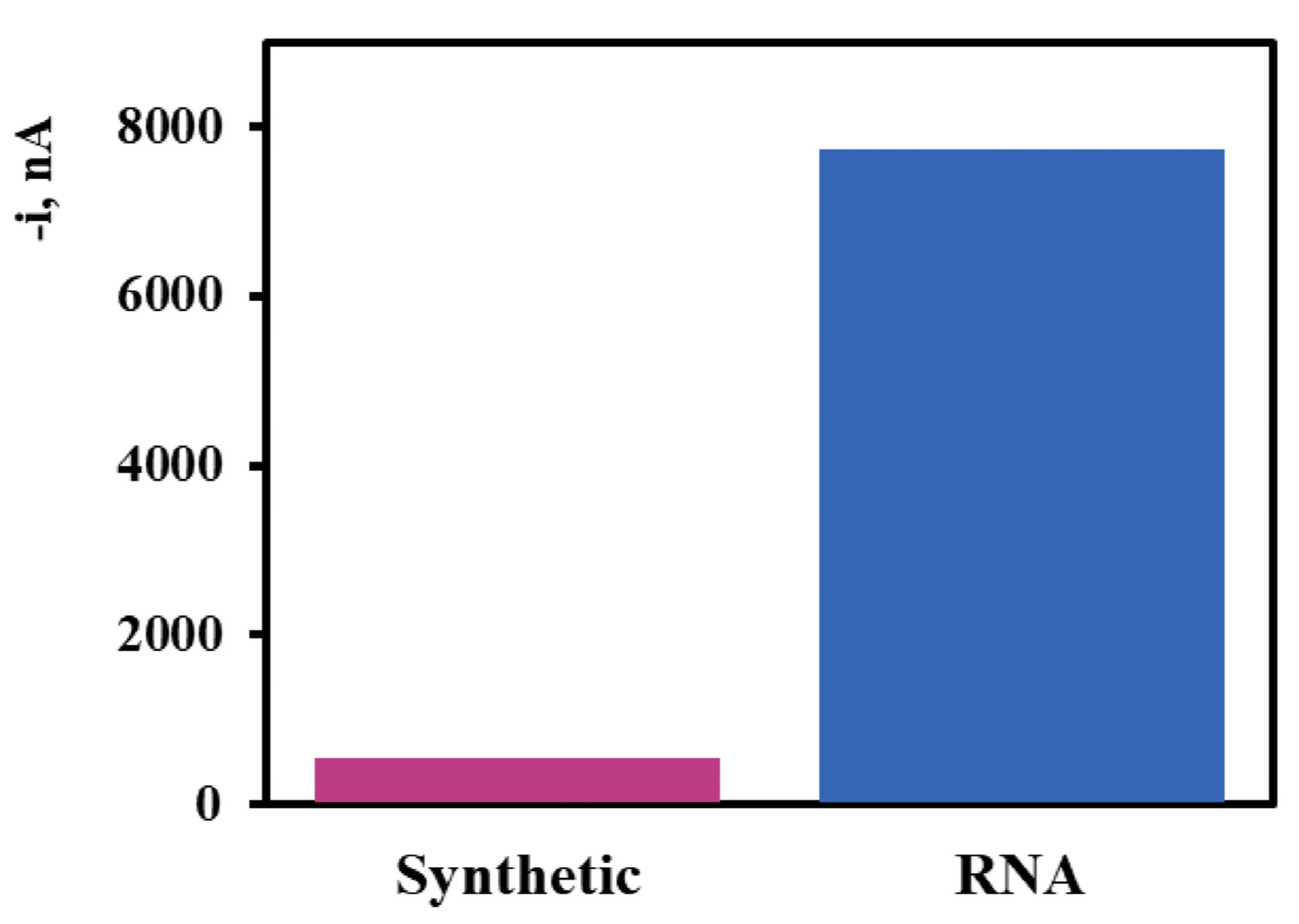
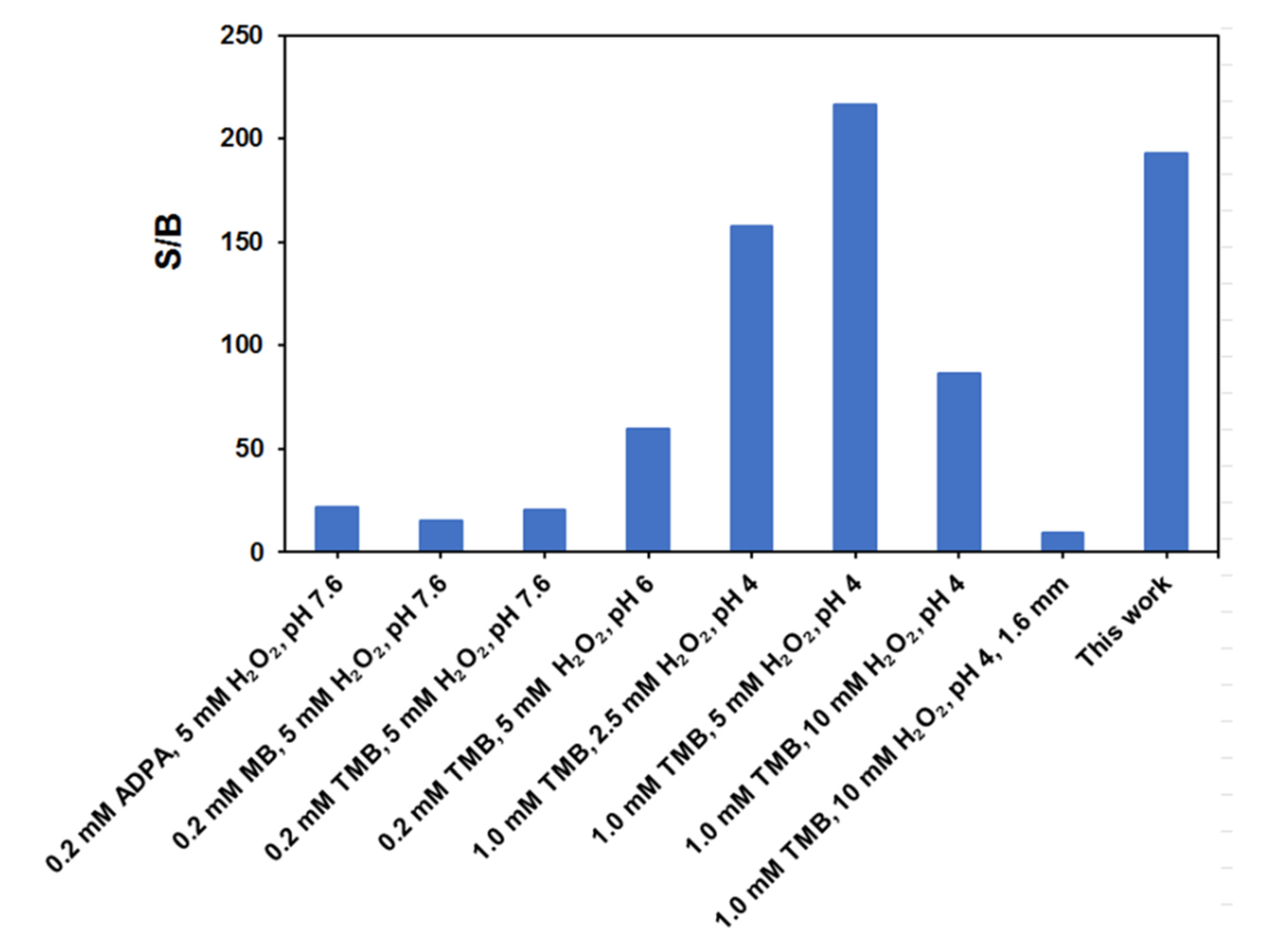
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aligizaki, K.; Nikolaidis, G. Morphological identification of two tropical dinoflagellates of the genera Gambierdiscus and Sinophysis in the Mediterranean Sea. J. Biol. Res. Thessa. 2008, 9, 75–82. [Google Scholar]
- Otero, P.; Pérez, S.; Alfonso, A.; Vale, C.; Rodríguez, P.; Gouveia, N.N.; Gouveia, N.; Delgado, J.; Vale, P.; Hirama, M.; et al. First toxin profile of ciguateric fish in Madeira Arquipelago (Europe). Anal. Chem. 2010, 82, 6032–6039. [Google Scholar] [CrossRef] [PubMed]
- Correia, F.S. Um caso raro de intoxicação alimentar colectiva. Bol. Inst. Sup. Higiene 1946, 3, 216–221. [Google Scholar]
- Vale, P.; Sampayo, M.A.M. Domoic acid in Portuguese shellfish and fish. Toxicon 2001, 39, 893–904. [Google Scholar] [CrossRef]
- David, H.; Laza-Martinez, A.; Orive, E.; Silva, A.; Moita Vale, P.; Sampayo, M.A.D.M. First confirmation of human diarrhoeic poisonings by okadaic acid esters after ingestion of razor clams (Solen marginatus) and green crabs (Carcinus maenas) in Aveiro lagoon, Portugal and detection of okadaic acid esters in phytoplankton. Toxicon 2002, 40, 989–996. [Google Scholar]
- Uemura, D.; Hirata, Y.; Iwashita, T.; Naoki, H. Studies on palytoxins. Tetrahedron 1985, 41, 1007–1017. [Google Scholar] [CrossRef]
- Moore, R.E.; Bartolini, G. Structure of palytoxin. J. Am. Chem. Soc. 1981, 103, 2491–2494. [Google Scholar] [CrossRef]
- Deeds, J.R.; Schwartz, M.D. Human risk associated with palytoxin exposure. Toxicon 2010, 56, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Aligizaki, K.; Katikou, P.; Nikolaidis, G.; Panou, A. First episode of shellfish contamination by palytoxin-like compounds from Ostreopsis species (Aegean Sea, Greece). Toxicon 2008, 51, 418–427. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative palytoxin and its new analogue, ovatoxin-a, in Ostreopsis ovata collected along the Ligurian coasts during the 2006 toxic outbreak. J. Am. Soc. Mass Spectrom. 2008, 19, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Pfannkuchen, M.; Godrijan, J.; Pfannkuchen, D.M.; Iveša, L.; Kružić, P.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; et al. Toxin-producing Ostreopsis cf. ovata are likely to bloom undetected along coastal areas. Environ. Sci. Technol. 2012, 46, 5574–5582. [Google Scholar] [CrossRef] [PubMed]
- Cohu, S.; Lemée, R. Vertical distribution of the toxic epibenthic dinoflagellates Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis in the NW Mediterranean Sea. Cah. Biol. Mar. 2012, 53, 373–380. [Google Scholar]
- Ruiz-Valdepeñas Montiel, V.; Gutiérrez, M.L.; Torrente-Rodríguez, R.M.; Povedano, E.; Vargas, E.; Reviejo, Á.J.; Linacero, R.; Gallego, F.J.; Campuzano, S.; Pingarrón, J.M. Disposable amperometric polymerase chain reaction-free biosensor for direct detection of adulteration with horsemeat in raw lysates targeting Mitochondrial DNA. Anal. Chem. 2017, 89, 9474–9482. [Google Scholar]
- Ruiz-Valdepeñas Montiel, V.; Povedano, E.; Vargas, E.; Torrente-Rodríguez, R.M.; Pedrero, M.; Reviejo, A.J.; Campuzano, S.; Pingarrón, J.M. Comparison of different strategies for the development of highly sensitive electrochemical nucleic acid biosensors using neither nanomaterials nor nucleic acid amplification. ACS Sens. 2018, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Barros, M.A.; Barros, M.F.; Martín-Pedraza, L.; Vargas, E.; Benedé, S.; Villalba, M.; Rocha, J.M.; Campuzano, S.; Pingarrón, J.M. Direct PCR-free electrochemical biosensing of plant-food derived nucleic acids in genomic DNA extracts. Application to the determination of the key allergen Sola l 7 in tomato seeds. Biosens. Bioelectron. 2019, 137, 171–177. [Google Scholar] [CrossRef]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhukumar Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; Förster, W.; et al. ARB: A software environment for sequence data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [Green Version]
- Siegmund, K.; Ahlborn, C.; Richert, C. ChipCheckII—Predicting binding curves for multiple analyte strands on small DNA microarrays. Nucl. Acids 2008, 27, 376–388. Available online: chip.chemie.uni-stuttgart.de/software.html (accessed on 14 December 2020). [CrossRef]
- BO Santé–Protection Sociale–Solidarité. no 2010/8 du 15 Septembre 2010. p. 264. Available online: https://solidarites-sante.gouv.fr/fichiers/bo/2010/10-08/ste_20100008_0001_p000.pdf (accessed on 14 December 2020).
- Funari, E.; Manganelli, M.; Emanuela, T. Ostreospis cf. ovata blooms in coastal water: Italian guidelines to assess and manage the risk associated to bathing waters and recreational activities. Harmful Algae 2015, 50, 45–56. [Google Scholar]
- Lemée, R.; Mangialajo, L.; Cohu, S.; Amzil, Z.; Blanfune, A.; Chomerat, N.; Ganzin, N.; Gasparini, S.; Grossel, H.; Guidi-Guivard, L.; et al. Interactions between scientists, managers and policy makers in the framework of the French MediOs Project on Ostreopsis (2008–2010). Crypt. Algol. 2012, 33, 137–142. [Google Scholar] [CrossRef]
- ICES. ICES CM 2015/SSGEPD. In Proceedings of the Interim Report of the ICES—IOC Working Group on Harmful Algal Bloom Dynamics (WGHABD), Lisbon, Portugal, 13–18 April 2015; Volume 17, p. 77. Available online: https://meetings.pices.int/publications/scientific-reports (accessed on 14 December 2020).
- Rodriquez, F.; Fraga, S.; Ramilo, I.; Rial, P.; Figueroa, R.I.; Riobo, P.; Braso, I. Canary Islands (NE Atlantic) as a biodiversity ‘hotspot’ of Gambierdiscus: Implications for future trends of ciguatera in the area. Harmful Algae 2017, 67, 131–143. [Google Scholar] [CrossRef]
- David, H.; Laza-Martínez, A.; Miguel, I.; Orive, E. Broad distribution of Coolia monotis and restricted distribution of Coolia cf. canariensis (Dinophyceae) on the Atlantic coast of the Iberian Peninsula. Phycologia 2014, 53, 342–352. [Google Scholar]
- Accoroni, S.; Totti, C. The toxic benthic dinoflagellates of the genus Ostreopsis in temperate areas: A review. Adv. Oceanogr. Limnol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zang, H. Morphotypes of Prorocentrum lima (Dinophyceae) from Hainan Island, South China Sea: Morphological and molecular characterization. Phycologia 2015, 54, 503–516. [Google Scholar] [CrossRef]
- Nishimura, T.; Uchida, H.; Noguchi, R.; Oikawa, H.; Suzuki, T.; Funaki, H.; Ihara, C.; Hagino, K.; Arimitsui, S.; Tanii, Y.; et al. Abundance of the benthic dinoflagellate Prorocentrum and the diversity, distribution, and diarrhetic shellfish toxin production of Prorocentrum lima complex and P. caipirignum in Japan. Harmful Algae 2020, 96, 101687. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.-H.; Davidson, N. RNA:DNA hybrids are more stable than DNA:DNA duplexes in concentration perchlorate and trichloroacetate solution. Nucl. Acid Res. 1978, 5, 1627–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metfies, K.; Hujic, S.; Lange, M.; Medlin, L.K. Electrochemical detection of the toxic dinoflagellate A. ostenfeldii with a DNA biosensor. Biosens. Bioelectron. 2005, 20, 1349–1357. [Google Scholar] [CrossRef] [Green Version]
- Diercks, S.; Metfies, K.; Jäckel, S.; Medlin, L.K. The ALGADEC devise: Development and optimisation of a semi-automated rRNA biosensor for the detection of toxic algae. Harmful Algae 2011, 10, 395–401. [Google Scholar] [CrossRef]
- Orozco, J.; Villa, E.; Manes, C.-L.; Medlin, L.K.; Guillebault, D. Electrochemical RNA genosensors for toxic algal species: Enhancing selectivity and sensitivity. Talanta 2016, 161, 560–566. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Set No. | Probe Name | Target | Gene |
|---|---|---|---|
| 1 | AlOstS01C | Alexandrium ostenfeldii | SSU |
| AlOstS01S | SSU | ||
| AlOstS02C | SSU | ||
| AlOstS02S | SSU | ||
| AlOstS03C | SSU | ||
| AlOstS03S | SSU | ||
| 3 | ClDinoS01C | Class Dinoflagellata | SSU |
| ClDinoS01S | SSU | ||
| 4 | GaustD301C | Gambierdiscus australis | LSU D1-3 |
| GaustD301S | LSU D1-3 | ||
| 5 | GaustD801C | LSU D8-10 | |
| GaustD801S | LSU D8-10 | ||
| 6 | GSilvD301C | Gambierdiscus silvae | LSU D1-3 |
| GSilvD301S | LSU D1-3 | ||
| 7 | GExceD301C | Gambierdiscus excentricus | LSU D1-3 |
| GExceD301S | LSU D1-3 | ||
| 8 | GExceD801C | LSU D8-10 | |
| GExceD801S | LSU D8-10 | ||
| 9 | CMonoS01C | Coolia monotis | SSU |
| CMonoS01S | SSU | ||
| 10 | CtropD01C | Coolia tropicalis | LSU D1-3 |
| CtropD01S | LSU D1-3 | ||
| 11 | CCanCd1D01C | Coolia cf. canariensis | LSU D1-3 |
| CCanCd1D01S | LSU D1-3 | ||
| 12 | CCanCD2D01C | Coolia canariensis | LSU D1-3 |
| CCanCD2D01S | LSU D1-3 | ||
| 13 | CspD01C | Coolia sp. | |
| CspD01S | LSU D1-3 | ||
| 14 | CPalmCld1D01C | Coolia palmeriensis Clade 1 | LSU D1-3 |
| CPalmCld1D01S | LSU D1-3 | ||
| 15 | C.palmCLD2D01C | Coolia palmeriensis Clade 2 | LSU D1-3 |
| C.palmCLD2D01S | LSU D1-3 | ||
| 16 | CooMalCD1D01C | Coolia maliaensis | LSU D1-3 |
| CooMalCD1D01S | LSU D1-3 | ||
| 16’ | CooMalCD1D02C | LSU D1-3 | |
| CooMalCD1D02S | LSU D1-3 | ||
| 17 | GNOstreS01C | Genus Ostreopsis | LSU D1-3 |
| GNOstreS01S | LSU D1-3 | ||
| 18 | Ostsp1D01C | Ostreopsis sp. 1 | LSU D1-3 |
| Ostsp1D01S | LSU D1-3 | ||
| 19 | Ostsp1D03C | LSU D1-3 | |
| Ostsp1D03S | LSU D1-3 | ||
| 20 | Ostsp2D01C | Ostreopsis sp. 2 | LSU D1-3 |
| Ostsp2D01S | LSU D1-3 | ||
| 21 | Ostsp3D01C | Ostreopsis sp. 3 | LSU D1-3 |
| Ostsp3D01S | LSU D1-3 | ||
| 22 | Ostsp4D01C | Ostreopsis sp. 4 | LSU D1-3 |
| Ostsp4D01S | LSU D1-3 | ||
| 23 | Ostsp5D01C | Ostreopsis sp. 5 | LSU D1-3 |
| Ostsp5D01S | LSU D1-3 | ||
| 24 | Ostsp6D01C | Ostreopsis sp. 6 | LSU D1-3 |
| Ostsp6D01S | LSU D1-3 | ||
| 25 | Ostsp7D01C | Ostreopsis sp. 7 | LSU D1-3 |
| Ostsp7D01S | LSU D1-3 | ||
| 26 | Ostsp8D01C | Ostreopsis sp. 8 | LSU D1-3 |
| Ostsp8D01S | LSU D1-3 | ||
| 27 | Ostsp9D01C | Ostreopsis sp. 9 | LSU D1-3 |
| Ostsp9D01S | LSU D1-3 | ||
| 28 | OstlentD01C | Ostreopsis lentilosus | LSU D1-3 |
| OstlentD01S | LSU D1-3 | ||
| 29 | OstsiamD01C | Ostreopsis siamensis | LSU D1-3 |
| OstsiamD01S | LSU D1-3 | ||
| 30 | OstrhodD01C | Ostreopsis rhodensis | LSU D1-3 |
| OstrhodD01S | LSU D1-3 | ||
| 31 | OstovatD01C | Ostreopsis ovata | LSU D1-3 |
| OstovatD01S | LSU D1-3 | ||
| OstovatD02Competitor | LSU D1-3 | ||
| 32 | OstCfOvCld2D01C | Ostreopsis cf. ovata clade 2 | LSU D1-3 |
| OstCfOvCld2D01S | LSU D1-3 | ||
| OstCfOvCld2D02Comp | LSU D1-3 | ||
| 33 | OstCf.AD01C | Ostreopsis sp. 10 | LSU D1-3 |
| OstCf.AD01S | LSU D1-3 | ||
| 34 | OstCf.BD8/03C | Ostreopsis sp. 11 | LSU D1-3 |
| OstCf.BD8/03S | LSU D1-3 | ||
| 35 | OstCf.DD07C | Ostreopsis sp. 12 | LSU D1-3 |
| OstCf.DD07S | LSU D1-3 | ||
| 36 | LinpolyD01C | Lingulodinium polyheldrum | LSU D1-3 |
| LinpolyD01S | LSU D1-3 | ||
| 37 | VulrugD01C | Vulcanodinium rugosum | LSU D1-3 |
| Vul rugD01S | LSU D1-3 | ||
| 38 | UnkPLMo1D01C | Prorocentrum lima morpho 1 | LSU D1-3z |
| UnkPLMo1D01S | LSU D1-3 | ||
| 39 | UnkPLMo2D02C | Prorocentrum lima morpho 2 | LSU D1-3 |
| UnkPLMo2D02S | LSU D1-3 | ||
| 40 | UnkPlMo3D01C | Prorocentrum lima morpho 3 | LSU D1-3 |
| UnkPlMo3D01S | LSU D1-3 | ||
| 41 | UnkPlMo4D01S | Prorocentrum lima morpho 4 | LSU D1-3 |
| UnkPlMo4D01C | LSU D1-3 | ||
| 42 | UnkPlMo5D02C | Prorocentrum lima morpho 5 | LSU D1-3 |
| UnkPlMo5D02S | LSU D1-3 | ||
| 43 | unkMorC1D01C | Prorocentrum lima Unknown morphotype clade 1 | LSU D1-3 |
| unkMorC1D01S | LSU D1-3 | ||
| 44 | unkMorC1D02C | LSU D1-3 | |
| unkMorC1D02S | LSU D1-3 | ||
| 45 | unkMorC1D03C | LSU D1-3 | |
| unkMorC1D03S | LSU D1-3 | ||
| 46 | UnkMorC2D03C | Prorocentrum lima Unknown morphotype clade 2 | LSU D1-3 |
| unkMorC2D03S | |||
| 75 | PlimaD01C | Prorocentrum lima all clades | LSU D1-3 |
| PlimaD01S | LSU D1-3 | ||
| 76 | UnkPLMo2D01C | Prorocentrum lima morpho 2 | LSU D1-3 |
| 77 | Plima?D01C | ? Prorocentrum lima | LSU D1-3 |
| 78 | AminD01* | Alexandrium minutum | LSU D1-3 |
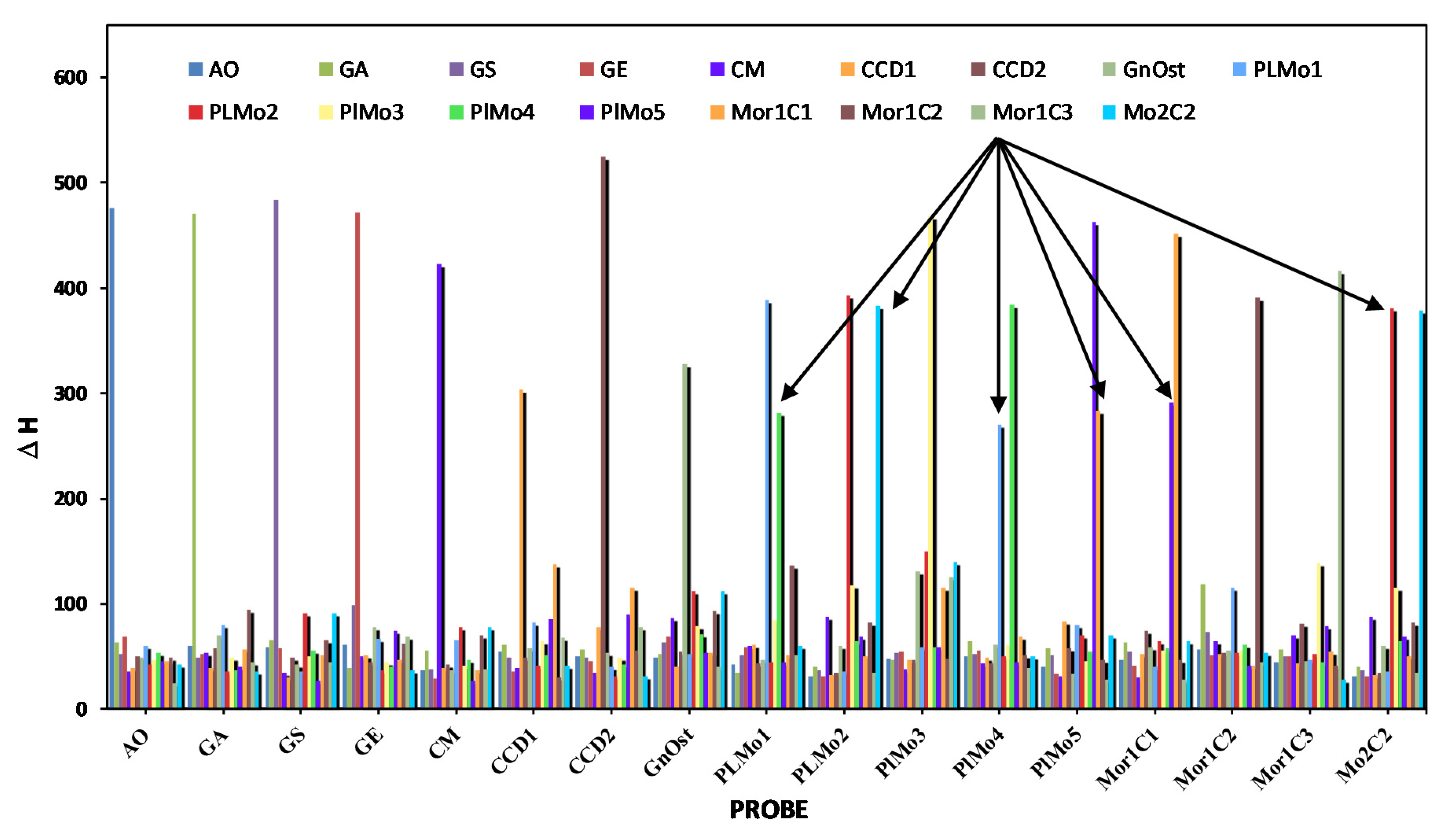
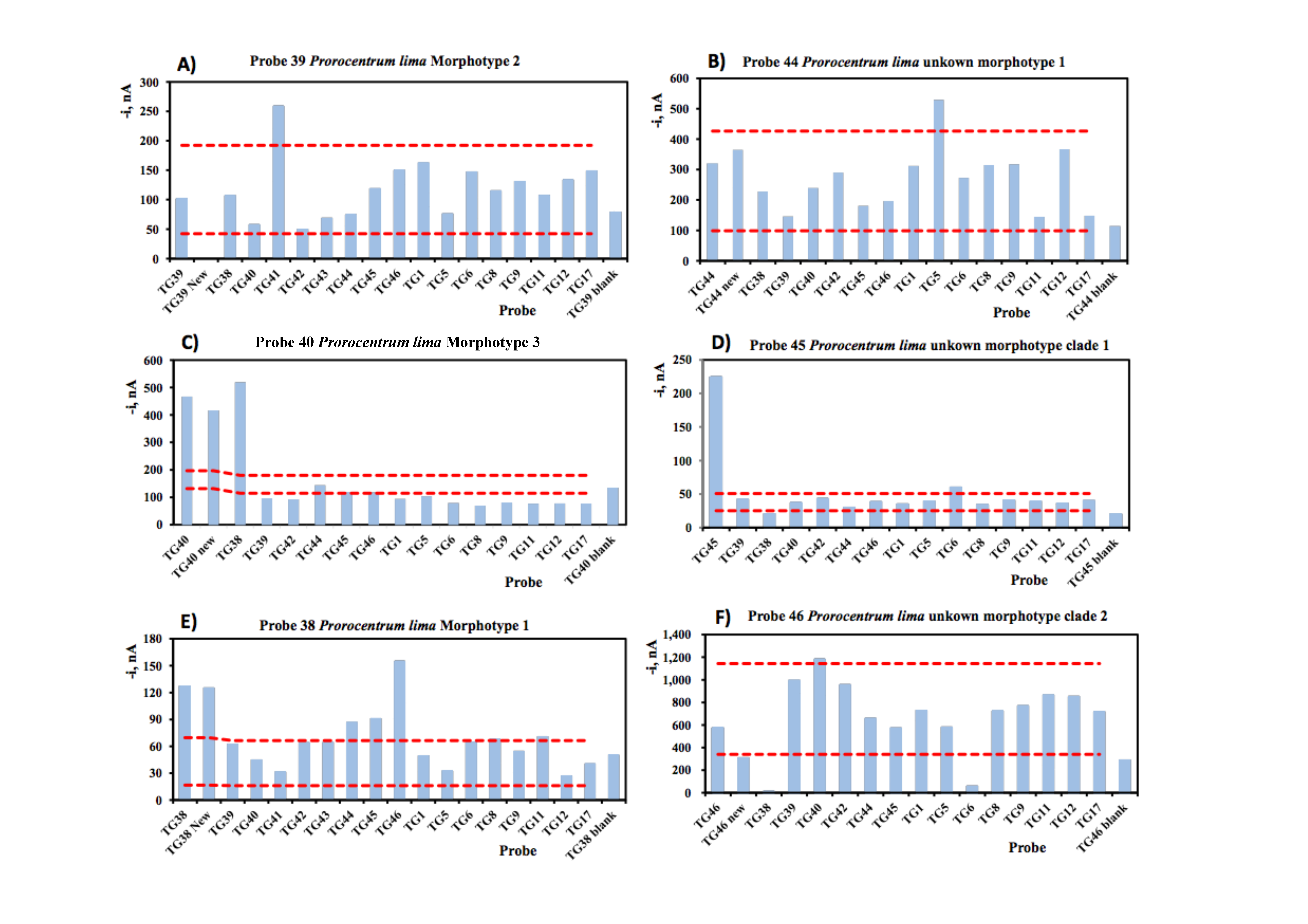
| 1 | 5 | 6 | 8 | 9 | 11 | 12 | 17 | 38 | 39 | 40 | 41 | 42 | 43 | 44 | 45 | 46 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AO | GA | GS | GE | CM | CCD1 | CCD2 | GnOst | PLMo1 | PLMo2 | PlMo3 | PlMo4 | PlMo5 | Mor1C1 | Mor1C2 | Mor1C3 | Mo2C2 | |
| AO | −475.7 | −60.5 | −59.4 | −61.6 | −36.7 | −54.9 | −50.7 | −48.6 | −42.3 | −31.3 | −48 | −50.6 | −40.4 | −47.3 | −57.1 | −44.3 | −31.3 |
| GA | −63.5 | −470.2 | −65.9 | −39.5 | −55.5 | −60.8 | −57.3 | −52.2 | −34.6 | −40.2 | −46.7 | −64.4 | −57.7 | −63.6 | −119.1 | −56.5 | −40.2 |
| GS | −52.2 | −48.7 | −484.1 | −98.6 | −38 | −49.1 | −48.6 | −63.3 | −50.8 | −37.1 | −53.2 | −51.9 | −51.5 | −54.6 | −73.8 | −50.1 | −37.1 |
| GE | −69.1 | −52.9 | −58.1 | −472 | −29.5 | −35.7 | −45.3 | −69.4 | −59.1 | −31.3 | −54.1 | −56.2 | −33.2 | −41.4 | −51.4 | −50.7 | −31.3 |
| CM | −35.8 | −53.3 | −34.6 | −50.7 | −422.6 | −38.7 | −35.2 | −87.1 | −60.4 | −87.9 | −38.3 | −44.1 | −31 | −30.6 | −64.4 | −70.2 | −87.9 |
| Ccf.Can | −38.7 | −39.6 | −30.7 | −51.6 | −39.1 | −303.8 | −77.3 | −40.4 | −60.8 | −33 | −46.7 | −49.2 | −83.3 | −52.2 | −52.8 | −43.6 | −33 |
| CCan | −50.1 | −57.8 | −48.6 | −44.2 | −42 | −48.7 | −524.6 | −55.1 | −44.1 | −34.3 | −46.4 | −44.1 | −58.3 | −74.6 | −53.5 | −80.8 | −34.3 |
| GnOst | −49.4 | −69.8 | −42.4 | −77.3 | −36.9 | −58.3 | −53.9 | −328.3 | −46.5 | −60 | −130.6 | −61.6 | −33.1 | −59.3 | −55.2 | −46.2 | −60 |
| PLMo1 | −60.2 | −79.9 | −36 | −66.7 | −65.4 | −82 | −39.8 | −51.9 | −389.1 | −36.1 | −58.9 | −269.8 | −80.4 | −40.1 | −115.2 | −47.2 | −36.1 |
| PLMo2 | −43 | −35.3 | −91.5 | −36.7 | −77.6 | −41.3 | −31.5 | −111.6 | −44.7 | −392.9 | −149.9 | −49.9 | −70 | −64.4 | −53.6 | −51.9 | −381.2 |
| PlMo3 | −47.2 | −48.8 | −49.8 | −45.1 | −41.5 | −65.1 | −49 | −79.3 | −84.2 | −118.2 | −467.8 | −61.3 | −45.3 | −56.2 | −55.4 | −138.3 | −114.9 |
| PlMo4 | −53.3 | −36.8 | −55.9 | −39.9 | −46.4 | −51.5 | −42.5 | −71.1 | −281.6 | −65 | −58.9 | −384.1 | −54.8 | −57.4 | −61.3 | −44.8 | −65 |
| PlMo5 | −45.8 | −40.3 | −26.8 | −74.6 | −27 | −85.5 | −90.3 | −53 | −44.2 | −69 | −58.6 | −44.2 | −463.2 | −291 | −41.4 | −79.1 | −69 |
| Mor1C1 | −45.8 | −56.5 | −51.3 | −47 | −37 | −137.1 | −114.9 | −53 | −50.9 | −50.5 | −115.8 | −69.1 | −283.5 | −452 | −41.4 | −54.7 | −50.5 |
| Mor1C2 | −49.1 | −94.4 | −66.1 | −62.3 | −69.7 | −30.3 | −55.6 | −93.2 | −136.4 | −82.3 | −48 | −51.3 | −46.4 | −46.4 | −391.1 | −41.7 | −82.3 |
| Mor1C3 | −24.5 | −45.2 | −44.8 | −69.5 | −38.5 | −67.5 | −77.3 | −40 | −51.3 | −34.3 | −125.9 | −38.9 | −27.7 | −27.7 | −44.9 | −416.3 | −34.3 |
| Mo2C2 | −43 | −35.3 | −91.5 | −36.7 | −77.6 | −41.3 | −31.5 | −111.6 | −60.1 | −382.5 | −139.5 | −49.9 | −70 | −64.4 | −53.6 | −28.2 | −378.6 |
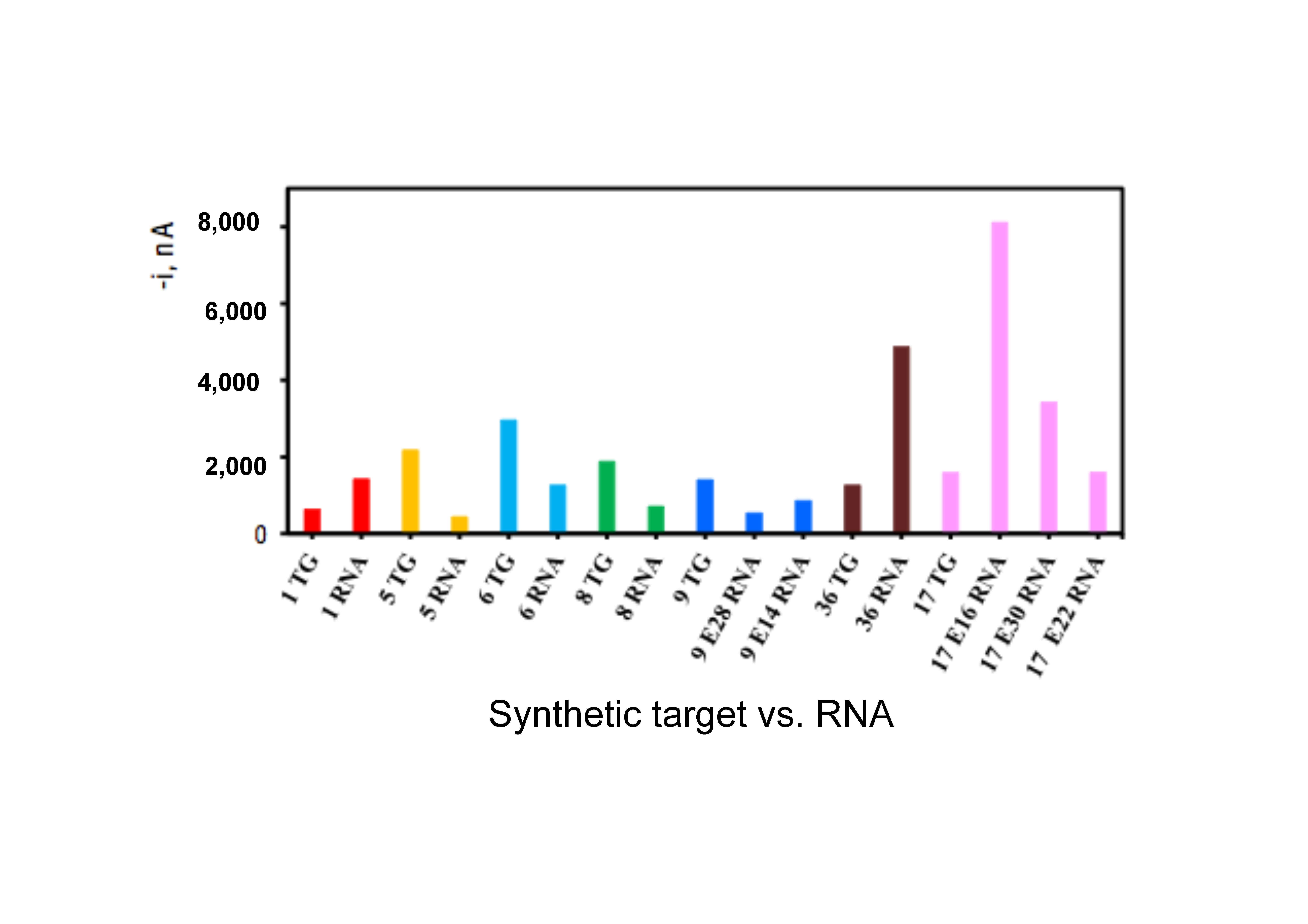
| Species/Strain from IEO VIGO and AWI | Number of Cells for 1 pM of RNA | Maximum Number of Cells Allowed (Per L) Before Fisheries Are Closed or More Intensive Toxin Testing is Required or Beaches Closed to Swimming |
|---|---|---|
| Prorocentrum lima/2V | 444 | 100 (UK) 500 (Spain) |
| P. lima/PL27V | 154 | 100 (UK) 500 (Spain) |
| P. lima/PL7V | 254 | 100 (UK) 500 (Spain) |
| P. lima/PLMA01 | 10 | 100 (UK) 500 (Spain) |
| Lingulodinium/1204 | 142 | 10,000 (0.000 (France)) |
| Coolia sp./VGO782 | 34 | N/A |
| Coolia sp./941 | 32 | N/A |
| Coolia malayensis/1163 | 117 | N/A |
| Coolia monotis/CMIV | 19 | N/A |
| Coolia monotis/VGO831 | 174 | N/A |
| Coolia tropicalis/923 | 57 | N/A |
| Ostreopsis cf ovata/1107 | 26 | 10-30,000 Italy 1, France |
| Ostreopsis cf ovata/614 | 11 | 10–30,000 Italy, France |
| Ostreopsis cf ovata/820 | 43 | 10–30,000 Italy, France |
| Ostreopsis cf ovata/884 | 18 | 10–30,000 Italy, France |
| Ostreopsis cf ovata/1196 | 14 | 10–30,000 Italy, France |
| Ostreopsis cf ovata/1068 | 33 | 10–30,000 Italy, France |
| Ostreopsis cf ovata/898 | 22 | 10–30,000 Italy, France |
| Ostreopsis cf. ovata/693 | 12 | 10–30,000 Italy, France |
| Ostreopsis fattorussoi/1795 | 45 | 10–30,000 Italy, France |
| Alexandrium ostenfeldii | 56 | 40 (UK), 200 (Aus) |
| Alexandrium minutum | 41 | Presence (Northern Europe), 40–1000 (Southern Europe), 200 (Aus) |
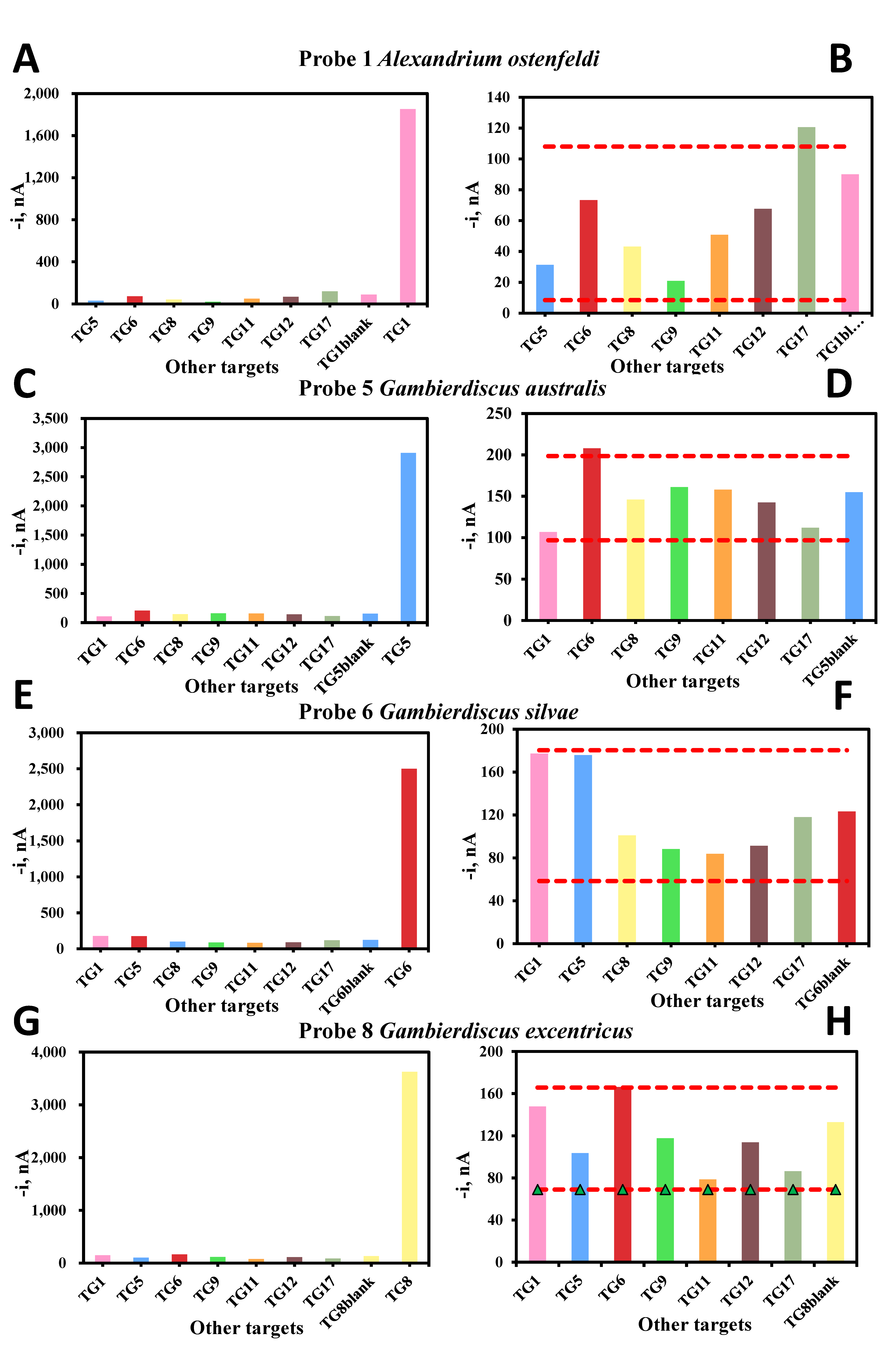
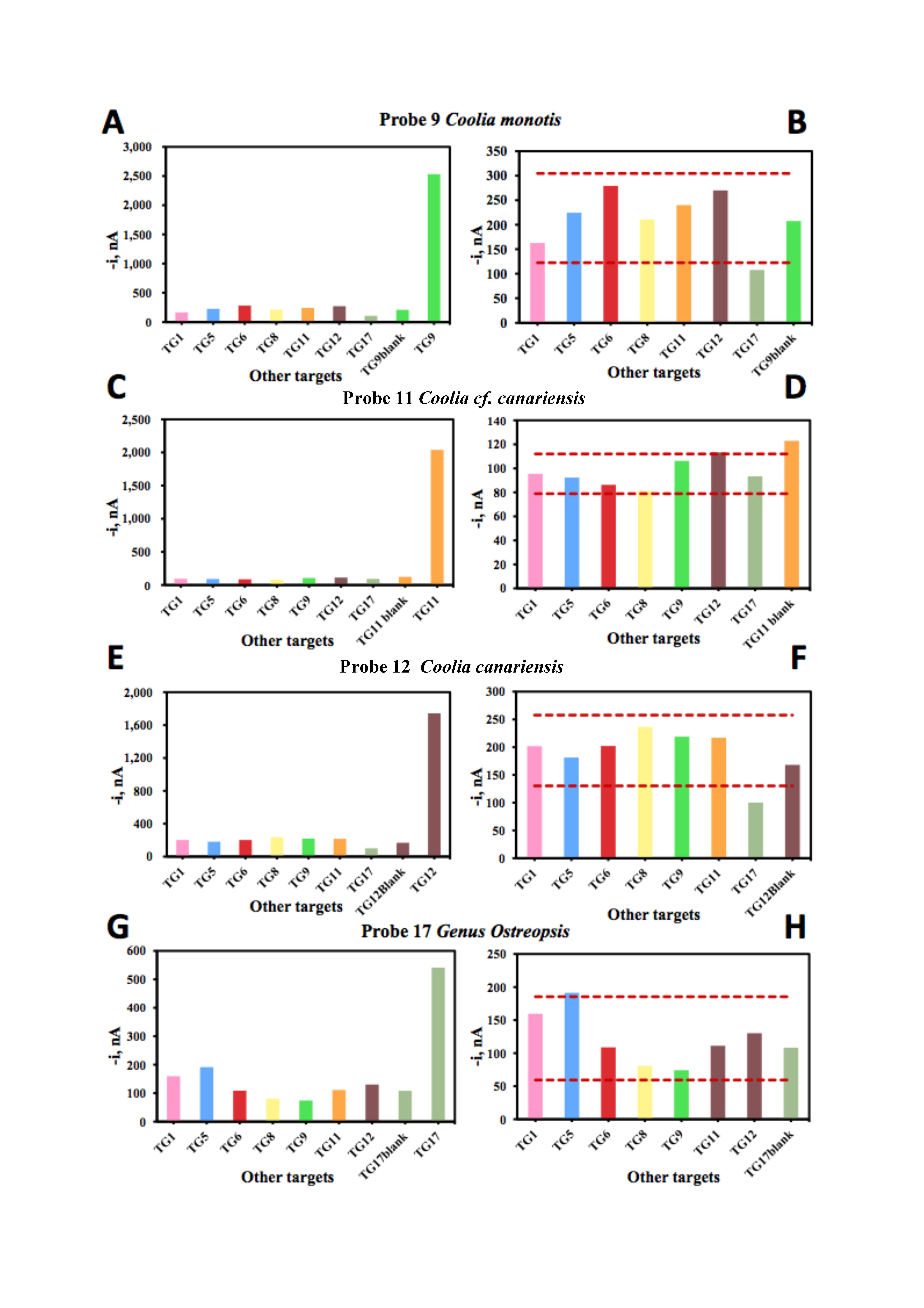
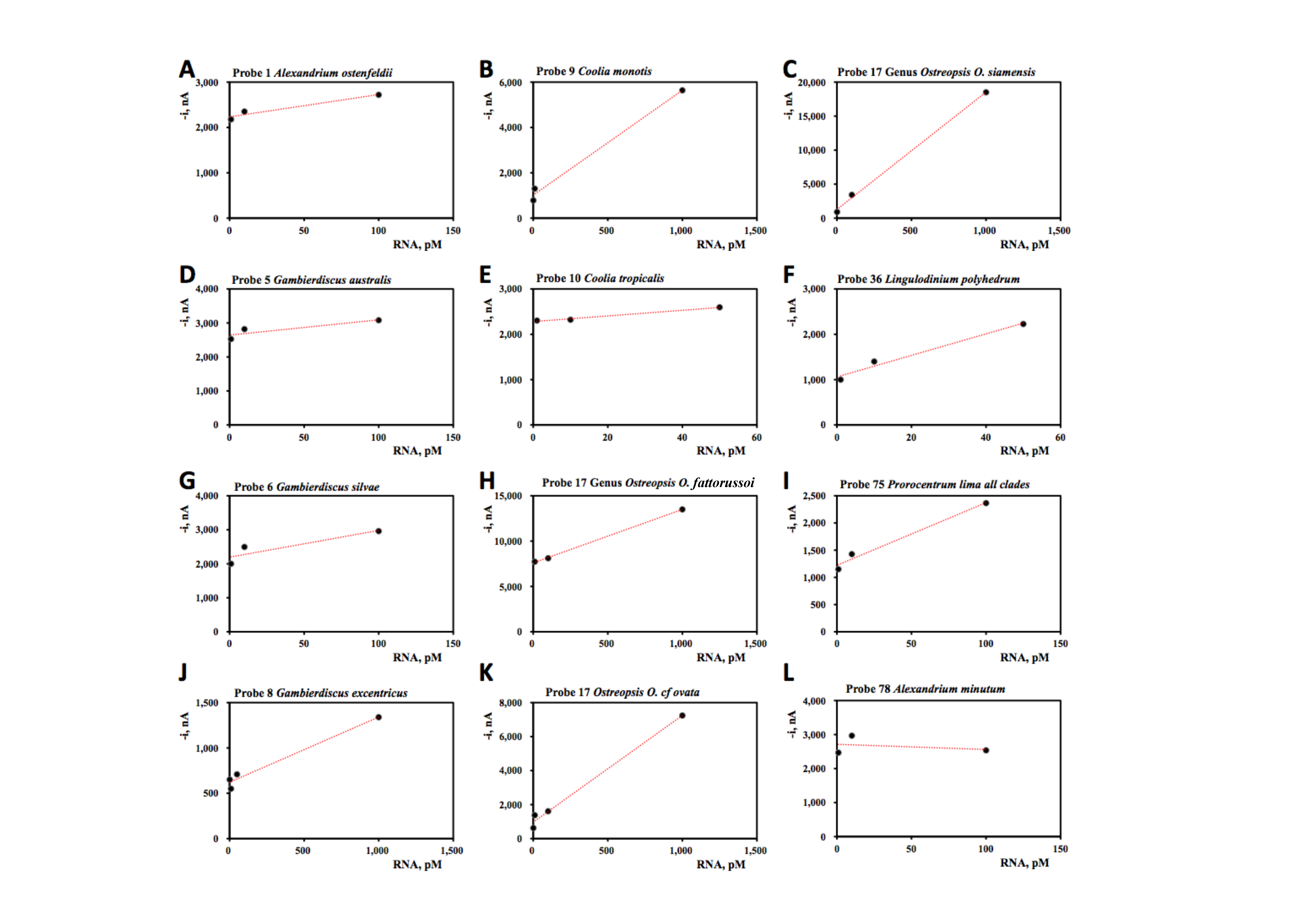
| Probe Number | Species Name | Slope, pM nA−1 | Intercept, nA | R2 |
|---|---|---|---|---|
| 1 | Alexandrium ostenfeldii | 4.9 ± 1.1 | 2233 ± 69 | 0.94 |
| 5 | Gambierdiscus australis | 4.5 ± 2.3 | 2642 ± 134 | 0.88 |
| 6 | Gambierdiscus silvae | 7.9 ± 3.9 | 2194 ± 227 | 0.80 |
| 8 | Gambierdiscus | 0.72 ± 0.08 | 621 ± 41 | 0.98 |
| 9 | Coolia monotis | 4.6 ± 0.4 | 1014 ± 240 | 0.99 |
| 10 | Coolia tropicalis | 6.2 ± 0.8 | 2280 ± 22 | 0.99 |
| 17 | Genus Osteopsis, O. fattorussoi | 5.9 ±0.1 | 7599 ± 79 | 0.99 |
| Genus Ostreopsis, O. cf. ovata | 6.3 ± 0.4 | 968 ± 209 | 0.99 | |
| Genus Ostreopsis, O. siamensis | 17 ± 1 | 1270 ± 432 | 0.99 | |
| 36 | Lingulodinium polyhedrum | 23.7 ± 3.6 | 1059 ± 107 | 0.99 |
| 75 | Prorocentrum lima all clades | 11 ± 2 | 1221 ± 93 | 0.98 |
| 78 | Alexandrium minutum | −1.5 ± 4.6 | 2713 ± 273 | 0.096 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medlin, L.K.; Gamella, M.; Mengs, G.; Serafín, V.; Campuzano, S.; M. Pingarrón, J. Advances in the Detection of Toxic Algae Using Electrochemical Biosensors. Biosensors 2020, 10, 207. https://doi.org/10.3390/bios10120207
Medlin LK, Gamella M, Mengs G, Serafín V, Campuzano S, M. Pingarrón J. Advances in the Detection of Toxic Algae Using Electrochemical Biosensors. Biosensors. 2020; 10(12):207. https://doi.org/10.3390/bios10120207
Chicago/Turabian StyleMedlin, Linda K., Maria Gamella, Gerardo Mengs, Verónica Serafín, Susana Campuzano, and José M. Pingarrón. 2020. "Advances in the Detection of Toxic Algae Using Electrochemical Biosensors" Biosensors 10, no. 12: 207. https://doi.org/10.3390/bios10120207