Dopamine and Striatal Neuron Firing Respond to Frequency-Dependent DBS Detected by Microelectrode Arrays in the Rat Model of Parkinson’s Disease

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Apparatus
2.3. MEA Fabrication Procedures
2.4. In Vivo Testing of MEAs
2.5. Data Acquisition and Analysis
3. Results
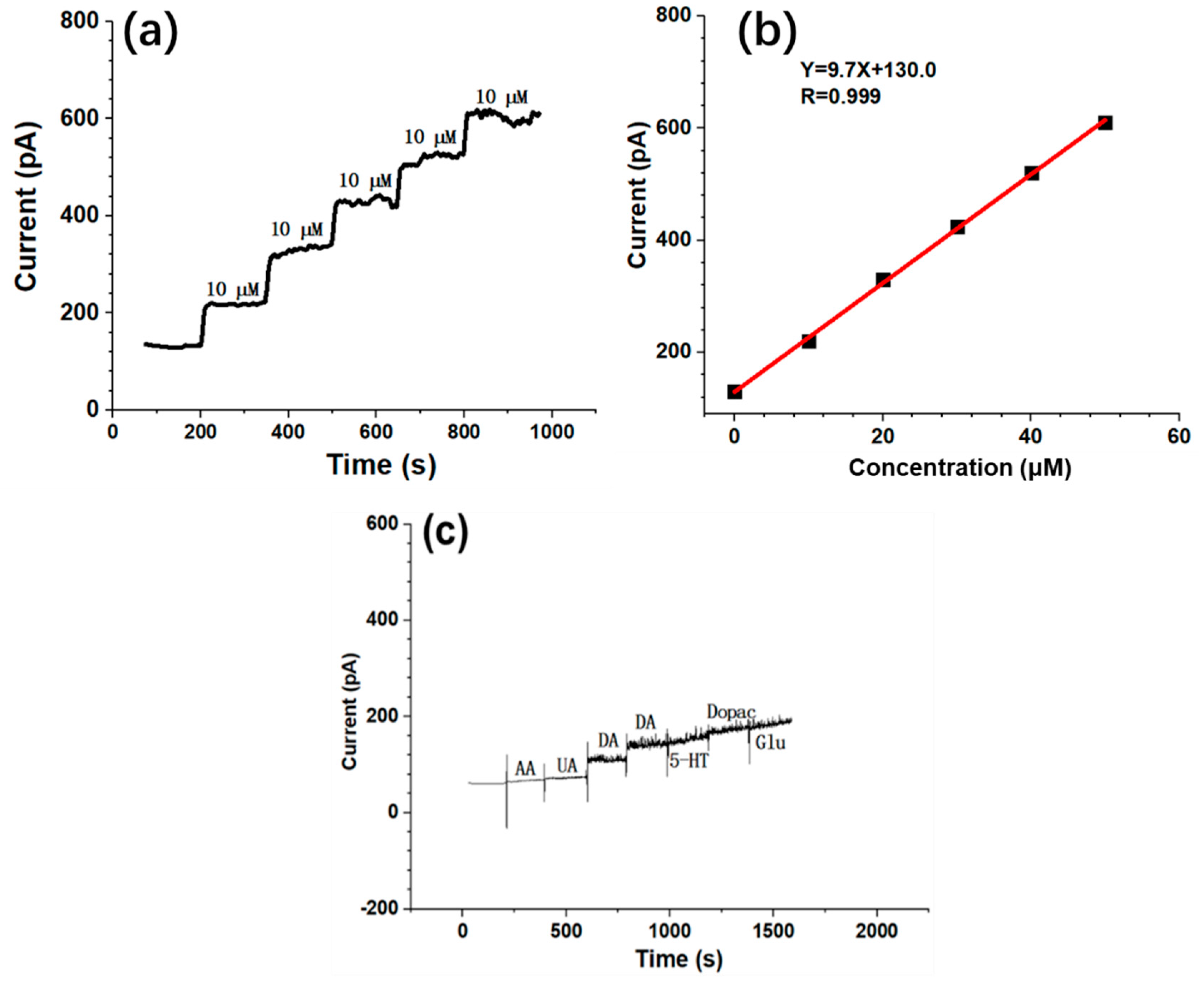
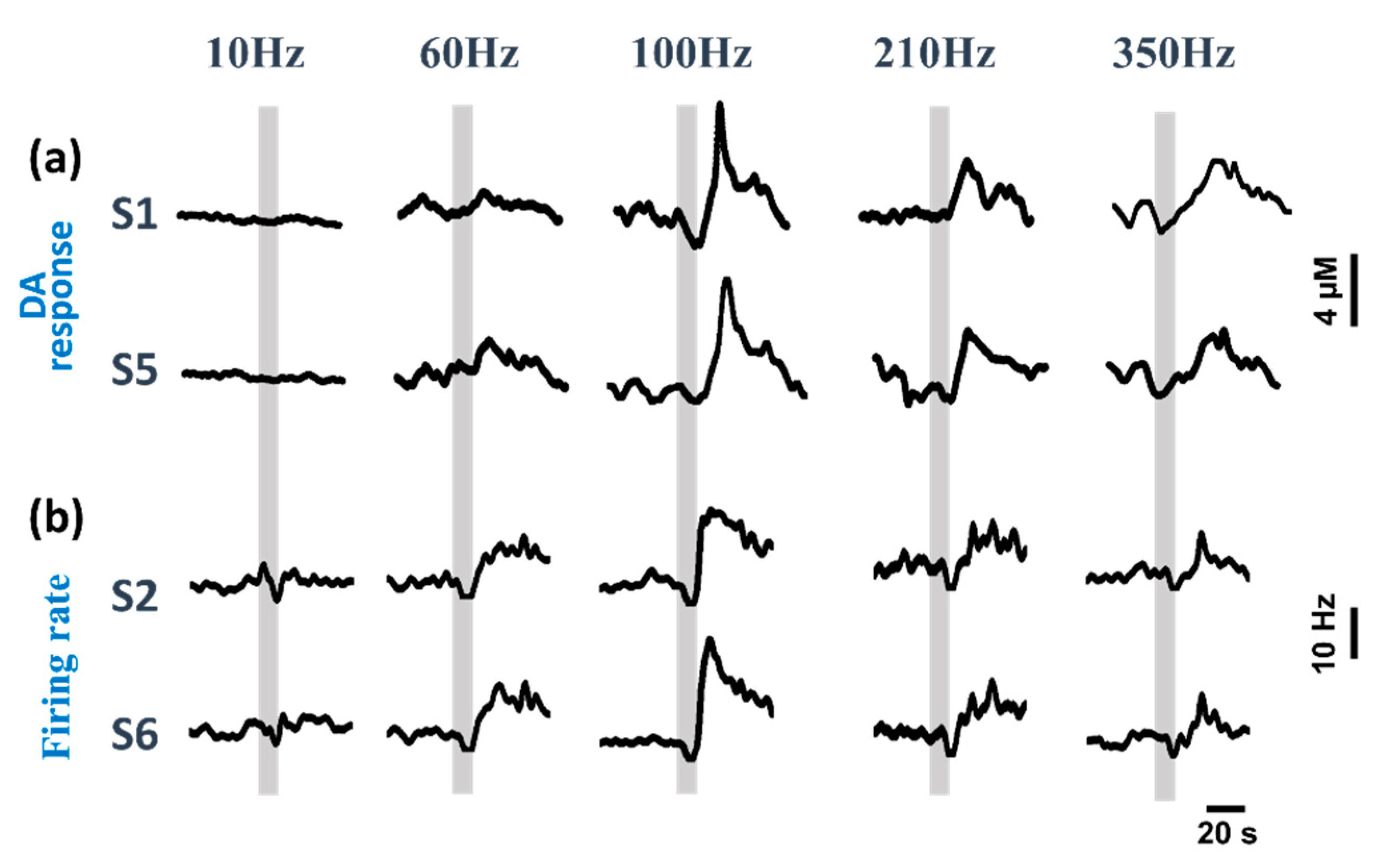
3.1. Neural Spike Firing Variations during DBS
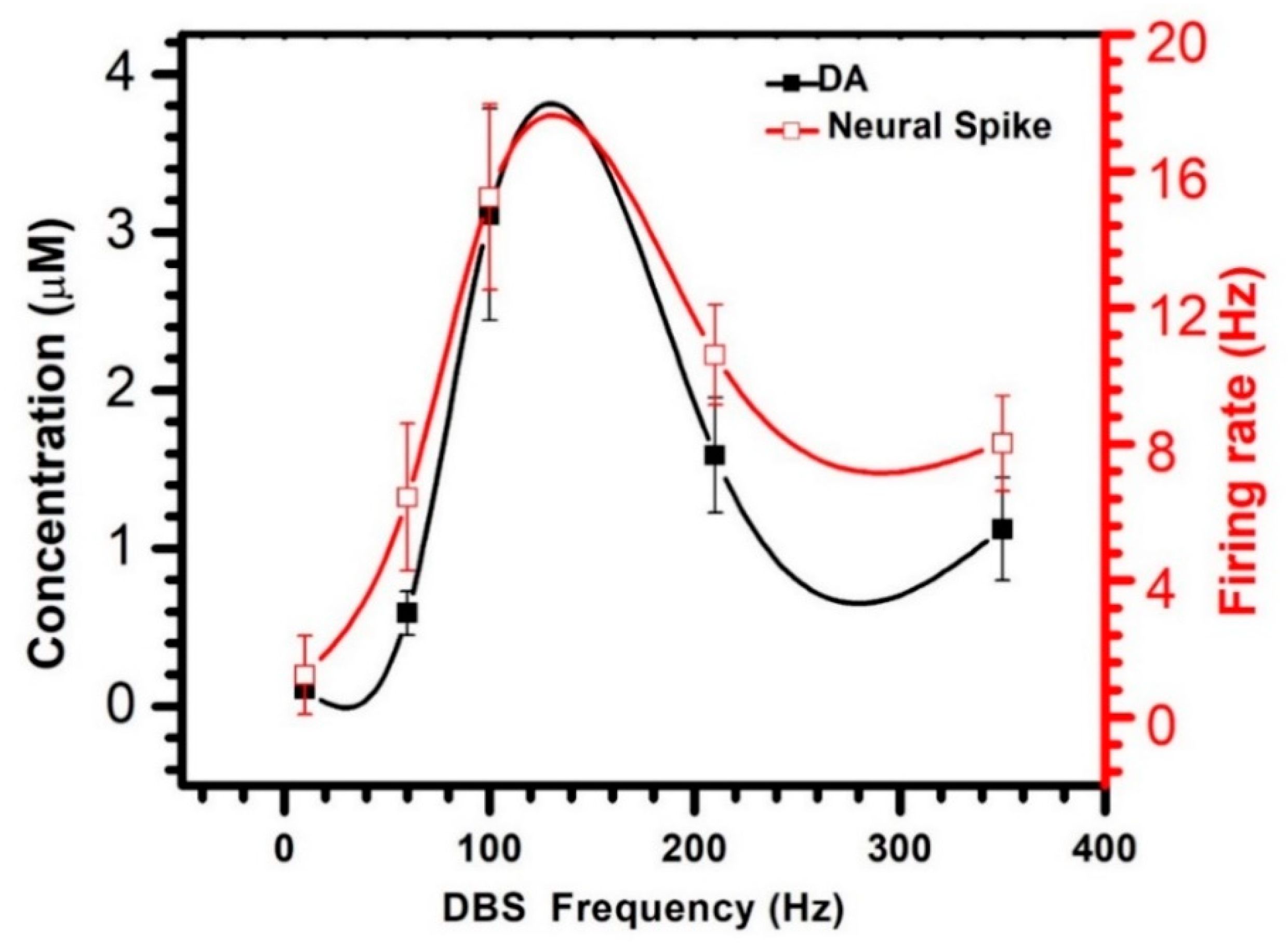
3.2. Dual-Mode Signals Variations during DBS
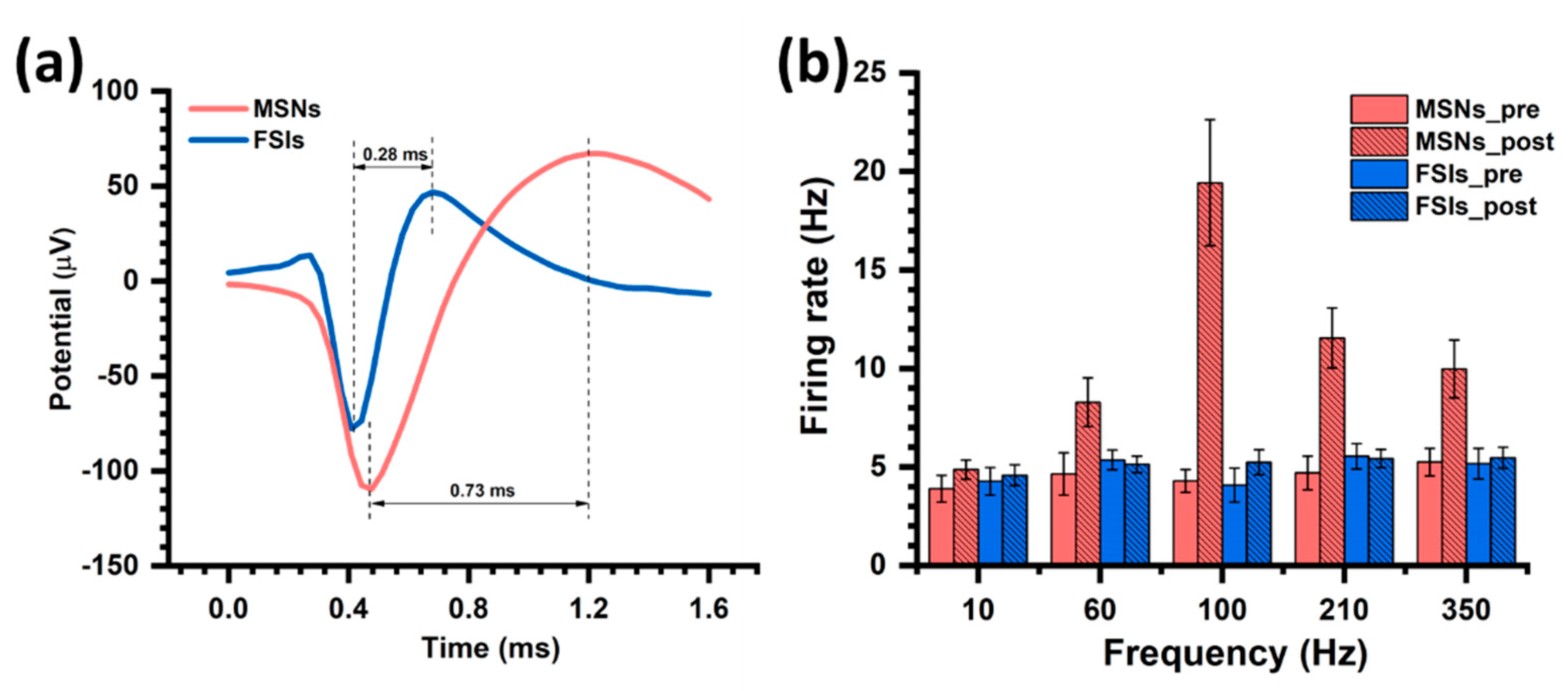
3.3. MSNs and FSIs Respond Different to DBS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andoskin, P.; Emelyanov, A.; Nikolaev, M.; Senkevich, K.; Shilin, V.; Yakimovskiy, A.; Timofeeva, A.; Pchelina, S. Parkinson’s disease (PD) is the most common neurodegenerative disease. Sci. Notes I. P. Pavlov St. Petersburg State Med. Univ. 2015, 22, 14–17. [Google Scholar] [CrossRef] [Green Version]
- Petersson, P.; Kuhn, A.A.; Neumann, W.J.; Fuentes, R. Basal ganglia oscillations as biomarkers for targeting circuit dysfunction in Parkinson’s disease. Prog. Brain. Res. 2020, 252, 525–557. [Google Scholar] [CrossRef] [PubMed]
- Halje, P.; Brys, I.; Mariman, J.J.; da Cunha, C.; Fuentes, R.; Petersson, P. Oscillations in cortico-basal ganglia circuits: Implications for Parkinson’s disease and other neurologic and psychiatric conditions. J. Neurophysiol. 2019, 122, 203–231. [Google Scholar] [CrossRef] [PubMed]
- Ghika, J.; Villemure, J.G.; Burkhard, P.; Russmanm, H.; Bogousslavsky, J.; Vingerhoets, F. GPi-DBS further improves a PD patient with STN-DBS and medication. Move. Disord. 2002, 17, S197. [Google Scholar]
- Villemure, J.G.; Vingerhoets, F.; Temperli, P.; Pollo, C.; Ghika, J. Effect of bilateral subthalamic deep brain stimulation (STN-DBS) after bilateral contemporaneous pallidal DBS (GPi-DBS) in Parkinson’s disease (PD). Neurology 2000, 54, A186. [Google Scholar]
- Lattanzi, S.; Silvestrini, M. Optimal achieved blood pressure in acute intracerebral hemorrhage: INTERACT2. Neurology 2015, 85, 557–558. [Google Scholar] [CrossRef] [Green Version]
- Miravite, J.; Deik, A.; Swan, M.; Sarva, H.; Kopell, B.H.; Severt, W.L. Parkinsonism and dystonia in Lubag disease respond well to high pulse width/low-frequency globus pallidus interna DBS. Neurol. Clin. Pract. 2015, 5, 480–483. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Padmanaban, M.; Bloom, L.; MacCracken, E.; Bertacchi, B.; Dachman, A.; Warnke, P. Effect of low versus high frequency stimulation on freezing of gait and other axial symptoms in Parkinson patients with bilateral STN DBS: A mini-review. Transl. Neurodegener. 2017, 6, 13. [Google Scholar] [CrossRef]
- Okun, M.S.; Wu, S.S.; Fayad, S.; Ward, H.; Bowers, D.; Rosado, C.; Bowen, L.; Jacobson, C.; Butson, C.; Foote, K.D. Acute and Chronic Mood and Apathy Outcomes from a randomized study of unilateral STN and GPi DBS. PLoS ONE 2014, 9, e114140. [Google Scholar] [CrossRef]
- Summerson, S.R.; Kemere, C.T.; Aazhang, B. Current amplitude-dependent modulation of rotational behavior with GPi stimulation in the rodent model of Parkinson’s Disease. In Proceedings of the 2013 35th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Montréal, QC, Canada, 3–7 July 2013; pp. 783–786. [Google Scholar]
- Shah, V.V.; Goyal, S.; Palanthandalam-Madapusi, H.J. A Possible Explanation of How High-Frequency Deep Brain Stimulation Suppresses Low-Frequency Tremors in Parkinson’s Disease. IEEE Trans. Neural. Syst. Rehabil. Eng. 2017, 25, 2498–2508. [Google Scholar] [CrossRef]
- Witt, K.; Sauer, T.; Deuschl, G. EP 32. Frequency of subthalamic nucleus stimulation (STN) in Parkinson’s disease (PD) modifies response accuracy in a decision making task. Clin. Neurophysiol. 2016, 127, e189. [Google Scholar] [CrossRef]
- Ostrem, J.L.; Markun, L.C.; Glass, G.A.; Racine, C.A.; Volz, M.M.; Heath, S.L.; de Hemptinne, C.; Starr, P.A. Effect of frequency on subthalamic nucleus deep brain stimulation in primary dystonia. Parkinson. Relat. Disord. 2014, 20, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Bloom, L.; Padmanaban, M.; Bertacchi, B.; Kang, W.; MacCracken, E.; Dachman, A.; Vigil, J.; Satzer, D.; Zadikoff, C.; et al. Long-term effect of low frequency stimulation of STN on dysphagia, freezing of gait and other motor symptoms in PD. J. Neurol. Neurosurg. Psychiatry 2018, 89, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Hwynn, N.; Tagliati, M.; Alterman, R.L.; Limotai, N.; Zeilman, P.; Malaty, I.A.; Foote, K.D.; Morishita, T.; Okun, M.S. Improvement of both dystonia and tics with 60 Hz pallidal deep brain stimulation. Int. J. Neurosci. 2012, 122, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Florin, E.; Reck, C.; Burghaus, L.; Lehrke, R.; Gross, J.; Sturm, V.; Fink, G.R.; Timmermann, L. Ten Hertz thalamus stimulation increases tremor activity in the subthalamic nucleus in a patient with Parkinson’s disease. Clin. Neurophysiol. 2008, 119, 2098–2103. [Google Scholar] [CrossRef]
- de la Fuente-Fernandez, R. Imaging of Dopamine in PD and Implications for Motor and Neuropsychiatric Manifestations of PD. Front. Neurol. 2013, 4, 90. [Google Scholar] [CrossRef] [Green Version]
- Kawahata, I.; Kawahata, I.; Hasegawa, K.; Kase, R. Music and Dopamine–Potential in Movement Disorders. J. Brain Nerves 2020, 1–4. [Google Scholar] [CrossRef]
- Berke, J.D. Uncoordinated firing rate changes of striatal fast-spiking interneurons during behavioral task performance. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 10075–10080. [Google Scholar] [CrossRef] [Green Version]
- Kravitz, A.V.; Freeze, B.S.; Parker, P.R.; Kay, K.; Thwin, M.T.; Deisseroth, K.; Kreitzer, A.C. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature 2010, 466, 622–626. [Google Scholar] [CrossRef] [Green Version]
- Galati, S.; Mazzone, P.; Fedele, E.; Pisani, A.; Peppe, A.; Pierantozzi, M.; Brusa, L.; Tropepi, D.; Moschella, V.; Raiteri, M.; et al. Biochemical and electrophysiological changes of substantia nigra pars reticulata driven by subthalamic stimulation in patients with Parkinson’s disease. Eur. J. Neurosci. 2006, 23, 2923–2928. [Google Scholar] [CrossRef]
- Sato, N.; Haruta, M.; Ohta, Y.; Sasagawa, K.; Ohta, J.; Pewnim, N.; Jongprateep, O. Fe2O3/MWCNTs modified microdialysis electrode for dopamine detection. Mater. Res. Express 2019, 7. [Google Scholar] [CrossRef]
- Xiao, G.; Song, Y.; Zhang, Y.; Xing, Y.; Zhao, H.; Xie, J.; Xu, S.; Gao, F.; Wang, M.; Xing, G.; et al. Microelectrode Arrays Modified with Nanocomposites for Monitoring Dopamine and Spike Firings under Deep Brain Stimulation in Rat Models of Parkinson’s Disease. ACS Sens. 2019, 4, 1992–2000. [Google Scholar] [CrossRef]
- Xiao, G.; Xu, S.; Song, Y.; Zhang, Y.; Li, Z.; Gao, F.; Xie, J.; Sha, L.; Xu, Q.; Shen, Y.; et al. In situ detection of neurotransmitters and epileptiform electrophysiology activity in awake mice brains using a nanocomposites modified microelectrode array. Sens. Actuat. B Chem. 2019, 288, 601–610. [Google Scholar] [CrossRef]
- Albert, G.C.; Cook, C.M.; Prato, F.S.; Thomas, A.W. Deep brain stimulation, vagal nerve stimulation and transcranial stimulation: An overview of stimulation parameters and neurotransmitter release. Neurosci. Biobehav. Rev. 2009, 33, 1042–1060. [Google Scholar] [CrossRef] [PubMed]
- Wightman, R.M.; Zimmerman, J.B. Control of dopamine extracellular concentration in rat striatum by impulse flow and uptake. Brain. Res. Brain. Res. Rev. 1990, 15, 135–144. [Google Scholar] [CrossRef]
- Li, Q.; Ke, Y.; Chan, D.C.; Qian, Z.M.; Yung, K.K.; Ko, H.; Arbuthnott, G.W.; Yung, W.H. Therapeutic deep brain stimulation in Parkinsonian rats directly influences motor cortex. Neuron 2012, 76, 1030–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenenberger, P.; O’Neill, J.; Csicsvari, J. Activity-dependent plasticity of hippocampal place maps. Nat. Commun. 2016, 7, 11824. [Google Scholar] [CrossRef] [PubMed]
- English, D.F.; McKenzie, S.; Evans, T.; Kim, K.; Yoon, E.; Buzsaki, G. Pyramidal Cell-Interneuron Circuit Architecture and Dynamics in Hippocampal Networks. Neuron 2017, 96, 505–520.e507. [Google Scholar] [CrossRef] [Green Version]
- DeLong, M.R. Primate models of movement disorders of basal ganglia origin. Trends Neurosci. 1990, 13, 281–285. [Google Scholar] [CrossRef]
- Alexander, G.E.; Crutcher, M.D. Functional architecture of basal ganglia circuits: Neural substrates of parallel processing. Trends Neurosci. 1990, 13, 266–271. [Google Scholar] [CrossRef]
- Pisani, A.; Bonsi, P.; Centonze, D.; Calabresi, P.; Bernardi, G. Activation of D2-like dopamine receptors reduces synaptic inputs to striatal cholinergic interneurons. J. Neurosci. 2000, 20, RC69. [Google Scholar] [CrossRef] [PubMed]
- Aosaki, T.; Kiuchi, K.; Kawaguchi, Y. Dopamine D1-like receptor activation excites rat striatal large aspiny neurons in vitro. J. Neurosci. 1998, 18, 5180–5190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindenbach, D.; Bishop, C. Critical involvement of the motor cortex in the pathophysiology and treatment of Parkinson’s disease. Neurosci. Biobehav. Rev. 2013, 37, 2737–2750. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Lei, H.; Xu, Q. Neuronal activity pattern defects in the striatum in awake mouse model of Parkinson’s disease. Behav. Brain Res. 2018, 341, 135–145. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, G.; Song, Y.; Zhang, Y.; Xing, Y.; Xu, S.; Wang, M.; Wang, J.; Chen, D.; Chen, J.; Cai, X. Dopamine and Striatal Neuron Firing Respond to Frequency-Dependent DBS Detected by Microelectrode Arrays in the Rat Model of Parkinson’s Disease. Biosensors 2020, 10, 136. https://doi.org/10.3390/bios10100136
Xiao G, Song Y, Zhang Y, Xing Y, Xu S, Wang M, Wang J, Chen D, Chen J, Cai X. Dopamine and Striatal Neuron Firing Respond to Frequency-Dependent DBS Detected by Microelectrode Arrays in the Rat Model of Parkinson’s Disease. Biosensors. 2020; 10(10):136. https://doi.org/10.3390/bios10100136
Chicago/Turabian StyleXiao, Guihua, Yilin Song, Yu Zhang, Yu Xing, Shengwei Xu, Mixia Wang, Junbo Wang, Deyong Chen, Jian Chen, and Xinxia Cai. 2020. "Dopamine and Striatal Neuron Firing Respond to Frequency-Dependent DBS Detected by Microelectrode Arrays in the Rat Model of Parkinson’s Disease" Biosensors 10, no. 10: 136. https://doi.org/10.3390/bios10100136