
Effects of Carbon Nanoparticles and Chromium Combined Exposure in Native (Ruditapes decussatus) and Invasive (Ruditapes philippinarum) Clams
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Description
2.2. Experimental Conditions
2.3. Carbon Nanotubes Characterization
2.4. Chromium Determination in Seawater and Organisms
2.5. Biomarker Responses
2.6. Statistical Analysis
3. Results
3.1. Carbon Nanotubes Characterization
3.2. Chromium in Seawater and Organisms
3.3. Biological Responses
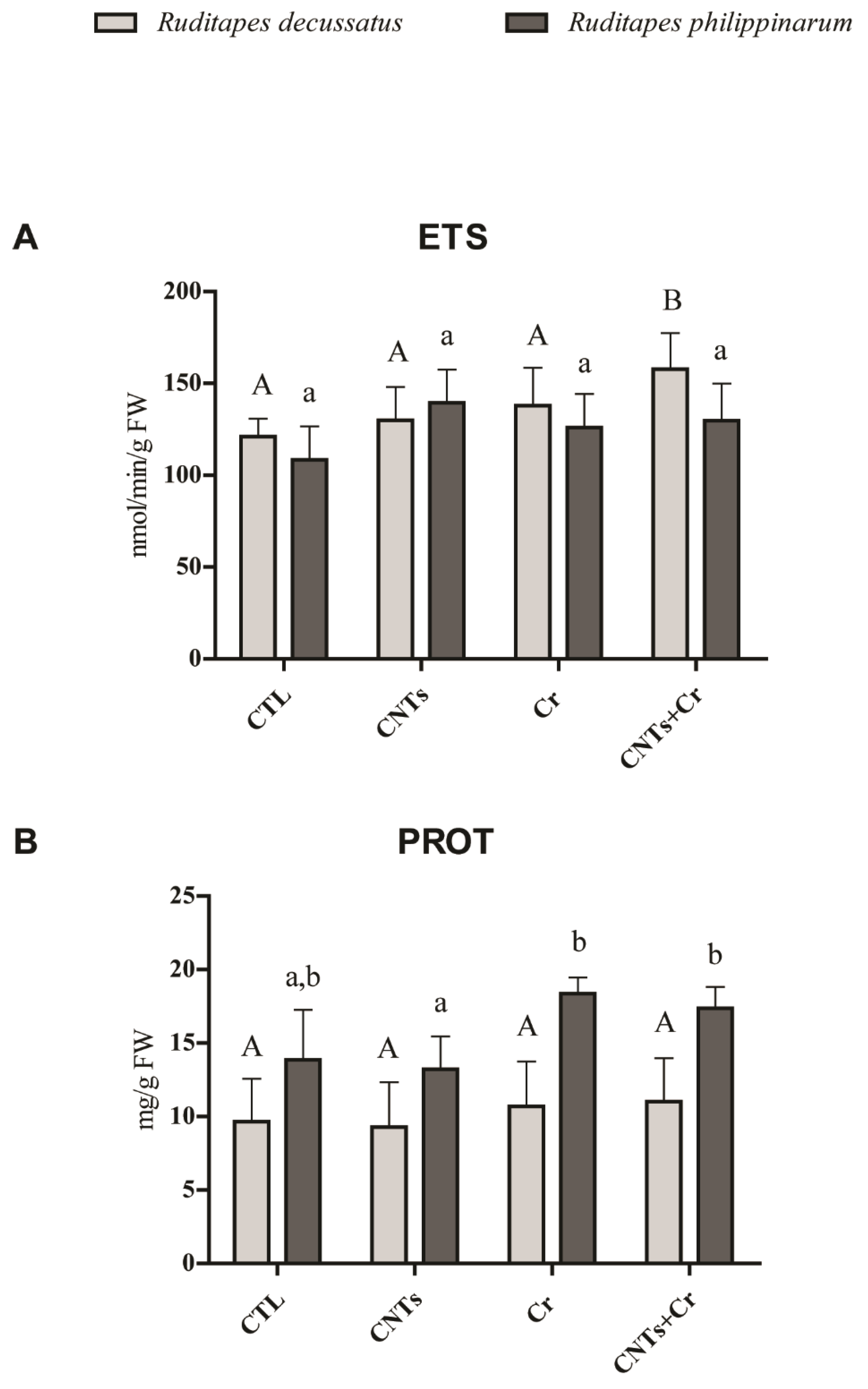
3.3.1. Metabolic Activity and Energy Reserves
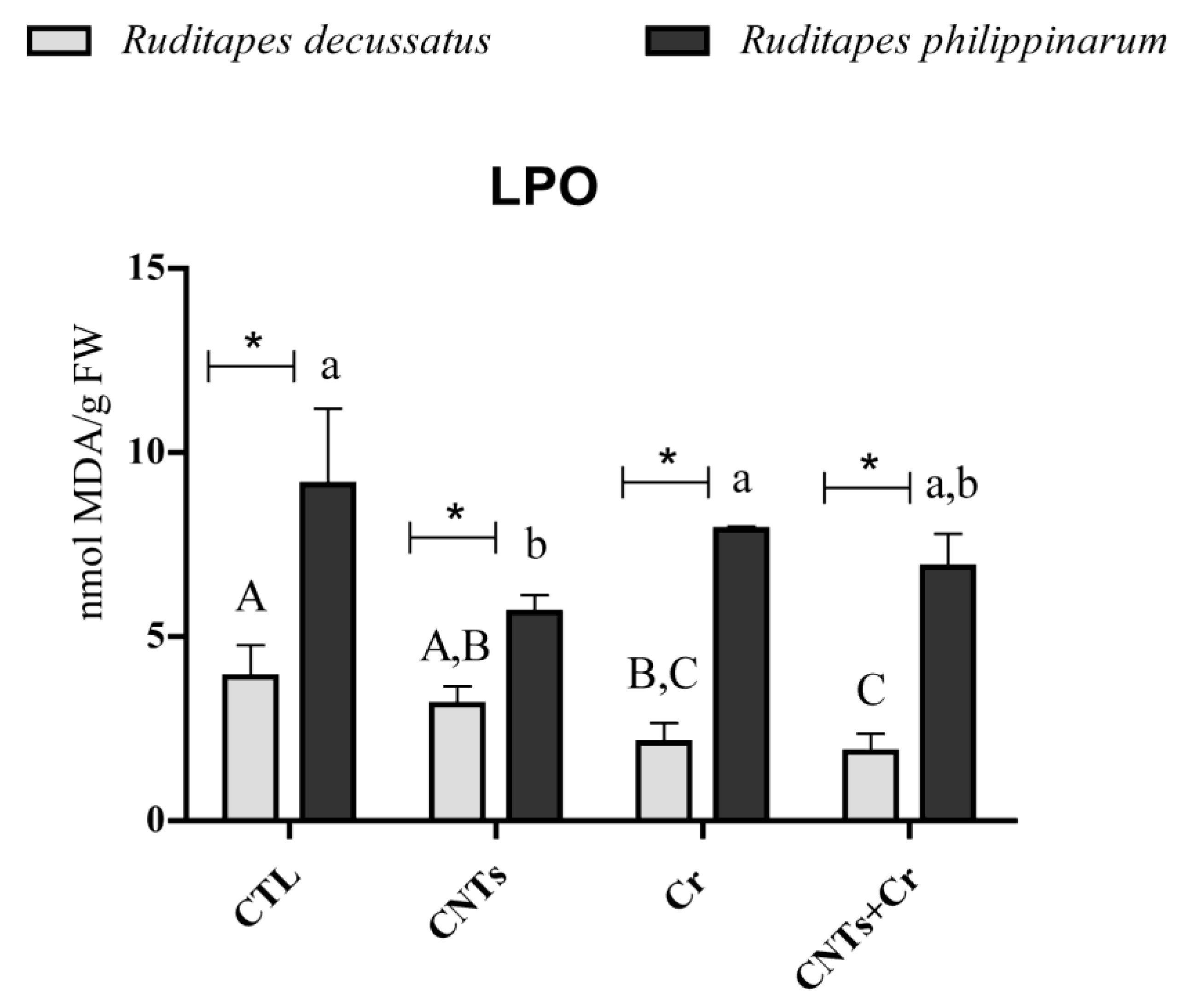
3.3.2. Indicators of Cellular Damage
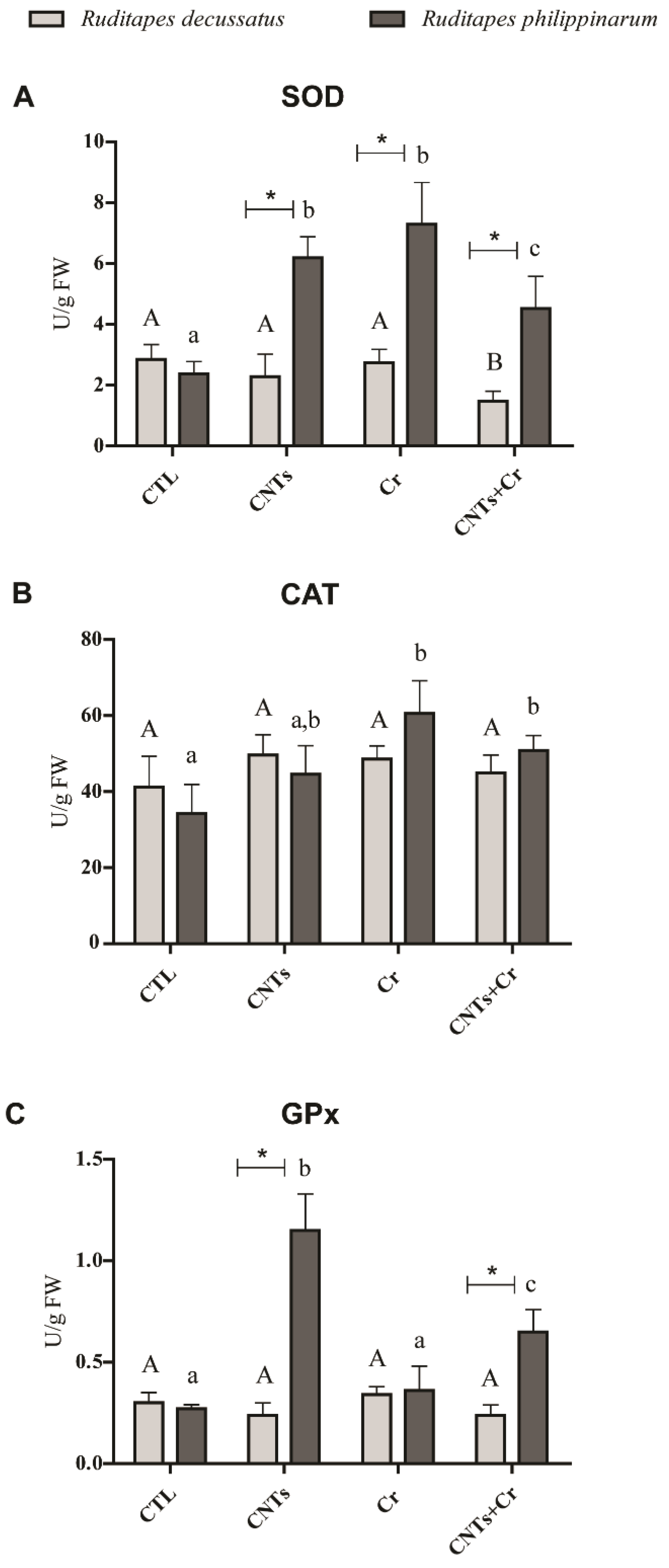
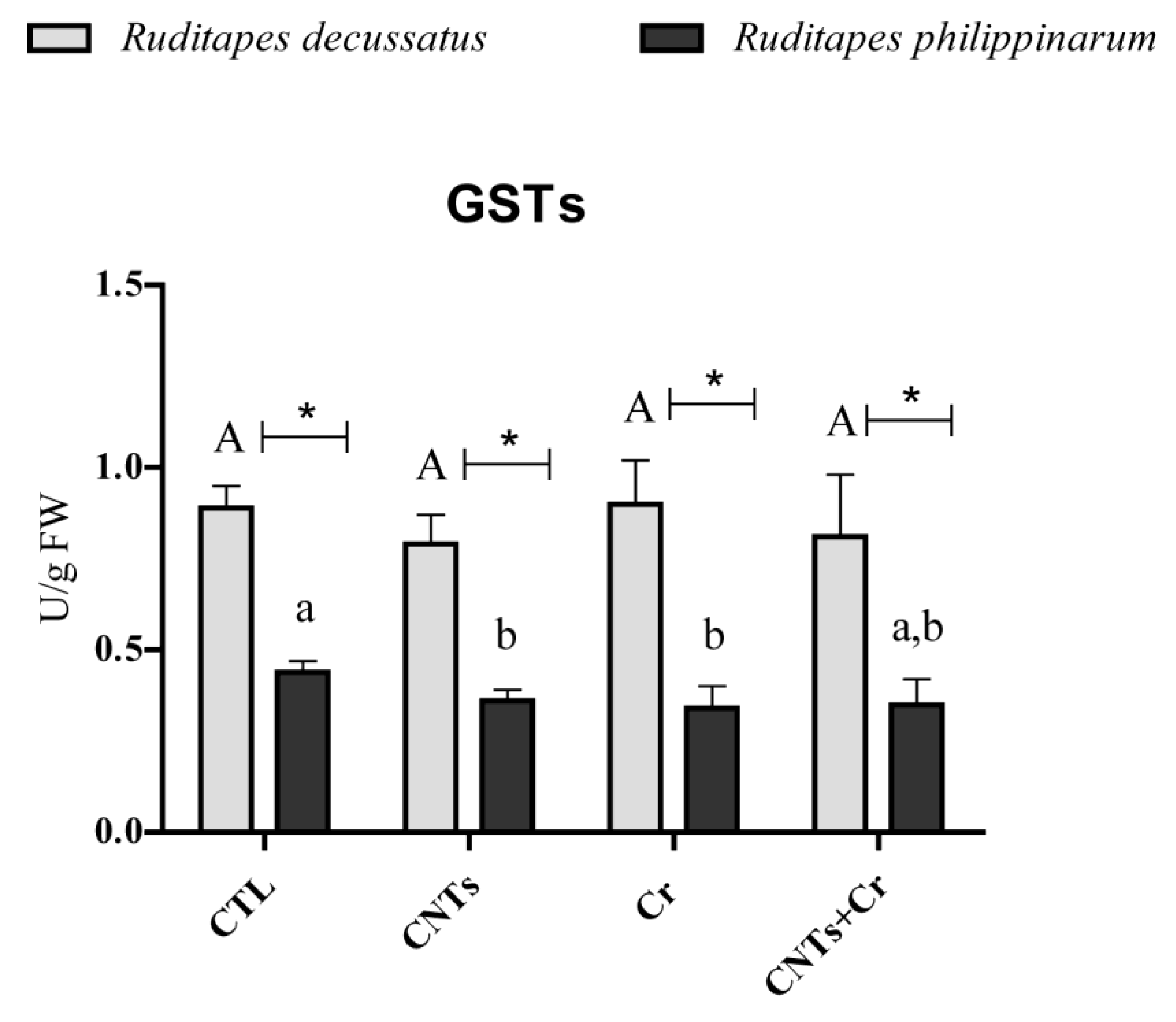
3.3.3. Antioxidant and Biotransformation Defenses
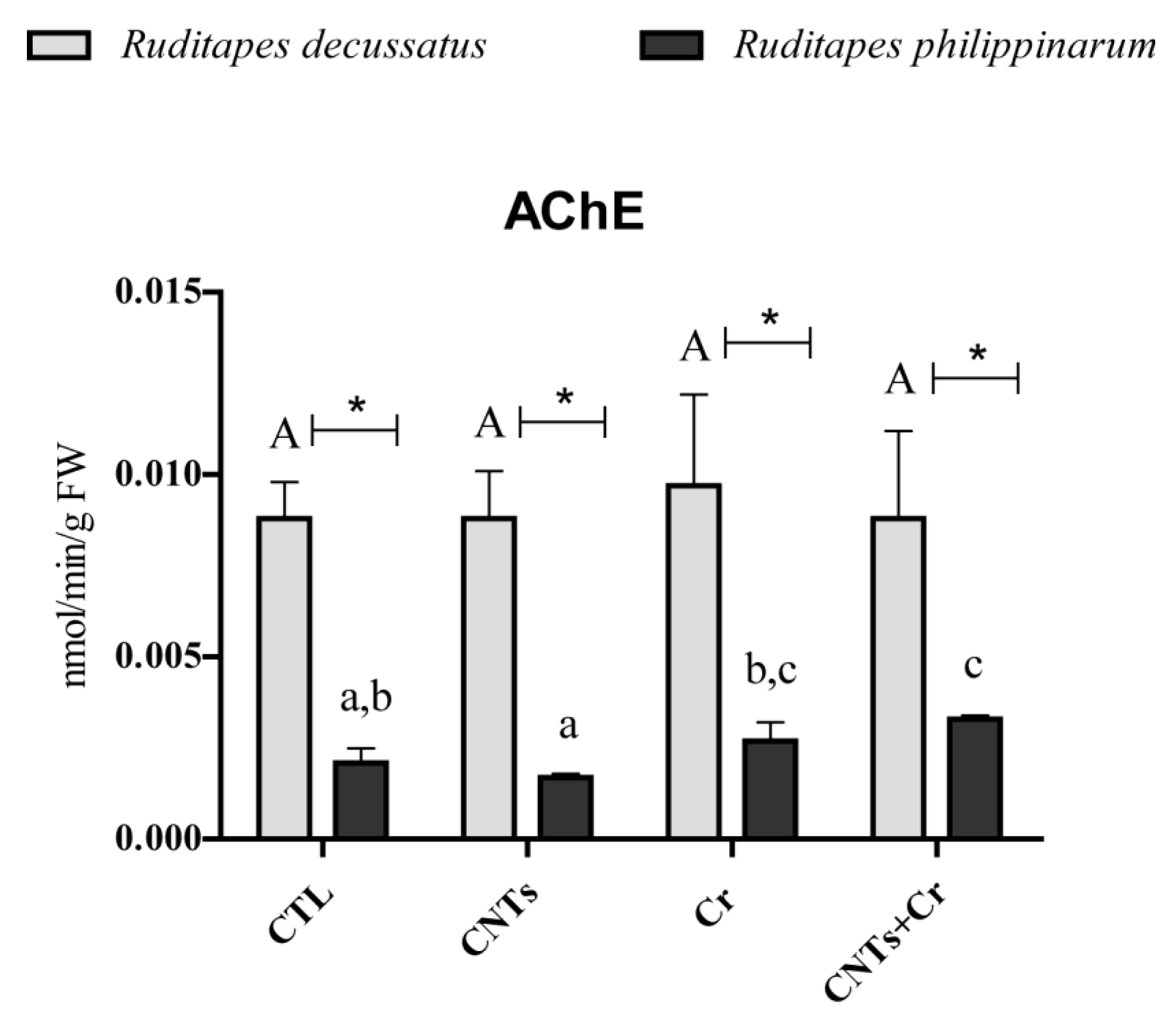
3.3.4. Neurotoxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Velma, V.; Vutukuru, S.S.; Tchounwou, P.B. Ecotoxicology of Hexavalent Chromium in Freshwater Fish: A Critical Review. Rev. Environ. Health 2009, 24, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Charoula, M.; Georgios, S.; Anna, B.; Elisavet, A. Correlation of Hexavalent Chromium Concentration to Groundwater Hydrochemical Zones Chemistry. Groundw. Sustain. Dev. 2021, 15, 100672. [Google Scholar] [CrossRef]
- Chiarelli, R.; Roccheri, M.C. Marine Invertebrates as Bioindicators of Heavy Metal Pollution. Open J. Met. 2014, 4, 93–106. [Google Scholar] [CrossRef]
- Urrutia, C.; Rudolph, A.; Lermanda, M.P.; Ahumada, R. Assessment of EDTA in Chromium (III-VI) Toxicity on Marine Intertidal Crab (Petrolisthes Laevigatus). Bull. Environ. Contam. Toxicol. 2008, 80, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Ciacci, C.; Barmo, C.; Gallo, G.; Maisano, M.; Cappello, T.; D’Agata, A.; Leonzio, C.; Mauceri, A.; Fasulo, S.; Canesi, L. Effects of Sublethal, Environmentally Relevant Concentrations of Hexavalent Chromium in the Gills of Mytilus Galloprovincialis. Aquat. Toxicol. 2012, 120–121, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Ciacci, C.; Barmo, C.; Fabbri, R.; Canonico, B.; Gallo, G.; Canesi, L. Immunomodulation in Mytilus Galloprovincialis by Non-Toxic Doses of Hexavalent Chromium. Fish Shellfish Immunol. 2011, 31, 1026–1033. [Google Scholar] [CrossRef]
- Emmanouil, C.; Sheehan, T.M.T.; Chipman, J.K. Macromolecule Oxidation and DNA Repair in Mussel (Mytilus Edulis L.) Gill Following Exposure to Cd and Cr(VI). Aquat. Toxicol. 2007, 82, 27–35. [Google Scholar] [CrossRef]
- Barmo, C.; Ciacci, C.; Fabbri, R.; Olivieri, S.; Bianchi, N.; Gallo, G.; Canesi, L. Pleiotropic Effects of Hexavalent Chromium (CrVI) in Mytilus Galloprovincialis Digestive Gland. Chemosphere 2011, 83, 1087–1095. [Google Scholar] [CrossRef]
- Dey, S.K.; Roy, S.; Chatterjee, A.K. Effect of Chromium on Certain Aspects of Metabolic Toxicities. Toxicol. Mech. Methods 2003, 13, 89–95. [Google Scholar] [CrossRef]
- Dahbi, S.; Azzi, M.; Saib, N.; de la Guardia, M.; Faure, R.; Durand, R. Removal of Trivalent Chromium from Tannery Waste Waters Using Bone Charcoal. Anal. Bioanal. Chem. 2002, 374, 540–546. [Google Scholar] [CrossRef]
- Salnikow, K.; Zhitkovich, A. Genetic and Epigenetic Mechanisms in Metal Carcinogenesis and Cocarcinogenesis: Nickel, Arsenic, and Chromium. Chem. Res. Toxicol. 2008, 21, 28–44. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Guidelines for Drinking-Water Quality, 4th ed.; WHO: Geneva, Switzerland, 2011; p. 216. ISBN 978-92-4-154995-0. [Google Scholar]
- Bakshi, A.; Panigrahi, A.K. A Comprehensive Review on Chromium Induced Alterations in Fresh Water Fishes. Toxicol. Rep. 2018, 5, 440–447. [Google Scholar] [CrossRef]
- Junaid, M.; Hashmi, M.Z.; Malik, R.N.; Pei, D.S. Toxicity and Oxidative Stress Induced by Chromium in Workers Exposed from Different Occupational Settings around the Globe: A Review. Environ. Sci. Pollut. Res. 2016, 23, 20151–20167. [Google Scholar] [CrossRef]
- Belay, A.A. Impacts of Chromium from Tannery Effluent and Evaluation of Alternative Treatment Options. J. Environ. Prot. 2010, 01, 53–58. [Google Scholar] [CrossRef]
- Ngoc, N.T.M.; Chuyen, N.V.; Thao, N.T.T.; Duc, N.Q.; Trang, N.T.T.; Binh, N.T.T.; Sa, H.C.; Tran, N.B.; Ba, N.V.; Khai, N.V.; et al. Chromium, Cadmium, Lead, and Arsenic Concentrations in Water, Vegetables, and Seafood Consumed in a Coastal Area in Northern Vietnam. Environ. Health Insights 2020, 14, 1178630220921410. [Google Scholar] [CrossRef]
- Rajeshkumar, S.; Liu, Y.; Zhang, X.; Ravikumar, B.; Bai, G.; Li, X. Studies on Seasonal Pollution of Heavy Metals in Water, Sediment, Fish and Oyster from the Meiliang Bay of Taihu Lake in China. Chemosphere 2018, 191, 626–638. [Google Scholar] [CrossRef]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, Applications and Toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- De Volder, M.F.L.; Tawfick, S.H.; Baughman, R.H.; Hart, A.J. Carbon Nanotubes: Present and Future Commercial Applications. Science 2013, 339, 535–539. [Google Scholar] [CrossRef]
- Mueller, N.C.; Nowack, B. Exposure Modeling of Engineered Nanoparticles in the Environment. Environ. Sci. Technol. 2008, 42, 4447–4453. [Google Scholar] [CrossRef]
- Petersen, E.J.; Zhang, L.; Mattison, N.T.; O’Carroll, D.M.; Whelton, A.J.; Uddin, N.; Nguyen, T.; Huang, Q.; Henry, T.B.; Holbrook, R.D.; et al. Potential Release Pathways, Environmental Fate, and Ecological Risks of Carbon Nanotubes. Environ. Sci. Technol. 2011, 45, 9837–9856. [Google Scholar] [CrossRef]
- Selck, H.; Handy, R.D.; Fernandes, T.F.; Klaine, S.J.; Petersen, E.J. Nanomaterials in the Aquatic Environment: A European Union-United States Perspective on the Status of Ecotoxicity Testing, Research Priorities, and Challenges Ahead. Environ. Toxicol. Chem. 2016, 35, 1055–1067. [Google Scholar] [CrossRef] [PubMed]
- Freixa, A.; Acuña, V.; Sanchís, J.; Farré, M.; Barceló, D.; Sabater, S. Ecotoxicological Effects of Carbon Based Nanomaterials in Aquatic Organisms. Sci. Total Environ. 2018, 619–620, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Petersen, E.J.; Huang, Q. Phase Distribution of 14C-Labeled Multiwalled Carbon Nanotubes in Aqueous Systems Containing Model Solids: Peat. Environ. Sci. Technol. 2011, 45, 1356–1362. [Google Scholar] [CrossRef] [PubMed]
- Bergami, E.; Pugnalini, S.; Vannuccini, M.L.; Manfra, L.; Faleri, C.; Savorelli, F.; Dawson, K.A.; Corsi, I. Long-Term Toxicity of Surface-Charged Polystyrene Nanoplastics to Marine Planktonic Species Dunaliella Tertiolecta and Artemia Franciscana. Aquat. Toxicol. 2017, 189, 159–169. [Google Scholar] [CrossRef]
- Caixeta, M.B.; Araújo, P.S.; Rodrigues, C.C.; Gonçalves, B.B.; Araújo, O.A.; Bevilaqua, G.B.; Malafaia, G.; Silva, L.D.; Rocha, T.L. Risk Assessment of Iron Oxide Nanoparticles in an Aquatic Ecosystem: A Case Study on Biomphalaria Glabrata. J. Hazard. Mater. 2021, 401, 123398. [Google Scholar] [CrossRef]
- Estrela, F.N.; Batista Guimarães, A.T.; Silva, F.G.; Marinho da Luz, T.; Silva, A.M.; Pereira, P.S.; Malafaia, G. Effects of Polystyrene Nanoplastics on Ctenopharyngodon Idella (Grass Carp) after Individual and Combined Exposure with Zinc Oxide Nanoparticles. J. Hazard. Mater. 2021, 403, 123879. [Google Scholar] [CrossRef]
- Klaper, R.; Crago, J.; Barr, J.; Arndt, D.; Setyowati, K.; Chen, J. Toxicity Biomarker Expression in Daphnids Exposed to Manufactured Nanoparticles: Changes in Toxicity with Functionalization. Environ. Pollut. 2009, 157, 1152–1156. [Google Scholar] [CrossRef]
- De Marchi, L.; Oliva, M.; Freitas, R.; Neto, V.; Figueira, E.; Chiellini, F.; Morelli, A.; Soares, A.M.V.M.; Pretti, C. Toxicity Evaluation of Carboxylated Carbon Nanotubes to the Reef-Forming Tubeworm Ficopomatus Enigmaticus (Fauvel, 1923). Mar. Environ. Res. 2019, 143, 1–9. [Google Scholar] [CrossRef]
- De Marchi, L.; Neto, V.; Pretti, C.; Figueira, E.; Chiellini, F.; Morelli, A.; Soares, A.M.V.M.; Freitas, R. The Influence of Salinity on the Effects of Multi-Walled Carbon Nanotubes on Polychaetes. Sci. Rep. 2018, 8, 8571. [Google Scholar] [CrossRef]
- Bergami, E.; Bocci, E.; Vannuccini, M.L.; Monopoli, M.; Salvati, A.; Dawson, K.A.; Corsi, I. Nano-Sized Polystyrene Affects Feeding, Behavior and Physiology of Brine Shrimp Artemia Franciscana Larvae. Ecotoxicol. Environ. Saf. 2016, 123, 18–25. [Google Scholar] [CrossRef]
- De Marchi, L.; Neto, V.; Pretti, C.; Figueira, E.; Chiellini, F.; Morelli, A.; Soares, A.M.V.M.; Freitas, R. Effects of Multi-Walled Carbon Nanotube Materials on Ruditapes Philippinarum under Climate Change: The Case of Salinity Shifts. Aquat. Toxicol. 2018, 199, 199–211. [Google Scholar] [CrossRef]
- Velzeboer, I.; Kupryianchyk, D.; Peeters, E.T.H.M.; Koelmans, A.A. Community Effects of Carbon Nanotubes in Aquatic Sediments. Environ. Int. 2011, 37, 1126–1130. [Google Scholar] [CrossRef]
- Piscopo, R.; Coppola, F.; Almeida, Â.; De Marchi, L.; Russo, T.; Esteves, V.I.; Soares, A.M.V.M.; Pretti, C.; Chiellini, F.; Polese, G.; et al. Effects of Temperature on Caffeine and Carbon Nanotubes Co-Exposure in Ruditapes Philippinarum. Chemosphere 2021, 271, 129775. [Google Scholar] [CrossRef]
- Naasz, S.; Altenburger, R.; Kühnel, D. Environmental Mixtures of Nanomaterials and Chemicals: The Trojan-Horse Phenomenon and Its Relevance for Ecotoxicity. Sci. Total Environ. 2018, 635, 1170–1181. [Google Scholar] [CrossRef]
- Hartmann, N.B.; Baun, A. The Nano Cocktail: Ecotoxicological Effects of Engineered Nanoparticles in Chemical Mixtures. Integr. Environ. Assess. Manag. 2010, 6, 311–313. [Google Scholar] [CrossRef]
- Cravo, A.; Pereira, C.; Gomes, T.; Cardoso, C.; Serafim, A.; Almeida, C.; Rocha, T.; Lopes, B.; Company, R.; Medeiros, A.; et al. A Multibiomarker Approach in the Clam Ruditapes Decussatus to Assess the Impact of Pollution in the Ria Formosa Lagoon, South Coast of Portugal. Mar. Environ. Res. 2012, 75, 23–34. [Google Scholar] [CrossRef]
- Flassch, J.; Leborgne, Y. Introduction in Europe, from 1972 to 1980, of the Japanese Manila Clam (Tapes Philippinarum) and the Effects on Aquaculture Production and Natural Settlement. ICES Mar. Sci. Symp. 1992, 194, 92–96. [Google Scholar]
- Anacleto, P.; Maulvault, A.L.; Barrento, S.; Mendes, R.; Nunes, M.L.; Rosa, R.; Marques, A. Physiological Responses to Depuration and Transport of Native and Exotic Clams at Different Temperatures. Aquaculture 2013, 408–409, 136–146. [Google Scholar] [CrossRef]
- Bidegain, G.; Bárcena, J.F.; García, A.; Juanes, J.A. Predicting Coexistence and Predominance Patterns between the Introduced Manila Clam (Ruditapes Philippinarum) and the European Native Clam (Ruditapes Decussatus). Estuar. Coast. Shelf Sci. 2015, 152, 162–172. [Google Scholar] [CrossRef]
- Velez, C.; Leandro, S.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. Biochemical Performance of Native and Introduced Clam Species Living in Sympatry: The Role of Elements Accumulation and Partitioning. Mar. Environ. Res. 2015, 109, 81–94. [Google Scholar] [CrossRef]
- Velez, C.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. The Impacts of As Accumulation under Different PH Levels: Comparing Ruditapes Decussatus and Ruditapes Philippinarum Biochemical Performance. Environ. Res. 2016, 151, 653–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez, C.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. Native and Introduced Clams Biochemical Responses to Salinity and PH Changes. Sci. Total Environ. 2016, 566–567, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Velez, C.; Freitas, R.; Antunes, S.C.; Soares, A.M.V.M.; Figueira, E. Clams Sensitivity towards As and Hg: A Comprehensive Assessment of Native and Exotic Species. Ecotoxicol. Environ. Saf. 2016, 125, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Jia, J.; Gong, X.; Luan, Z.; Fan, B. Aqueous Stability of Oxidized Carbon Nanotubes and the Precipitation by Salts. J. Hazard. Mater. 2009, 165, 1239–1242. [Google Scholar] [CrossRef]
- Ferguson, P.L.; Chandler, G.T.; Templeton, R.C.; DeMarco, A.; Scrivens, W.A.; Englehart, B.A. Influence of Sediment−Amendment with Single-Walled Carbon Nanotubes and Diesel Soot on Bioaccumulation of Hydrophobic Organic Contaminants by Benthic Invertebrates. Environ. Sci. Technol. 2008, 42, 3879–3885. [Google Scholar] [CrossRef]
- De Marchi, L.; Neto, V.; Pretti, C.; Figueira, E.; Chiellini, F.; Morelli, A.; Soares, A.M.V.M.; Freitas, R. Toxic Effects of Multi-Walled Carbon Nanotubes on Bivalves: Comparison between Functionalized and Nonfunctionalized Nanoparticles. Sci. Total Environ. 2018, 622–623, 1532–1542. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Q.; Zou, W.; Hu, X. Molecular Mechanisms of Developmental Toxicity Induced by Graphene Oxide at Predicted Environmental Concentrations. Environ. Sci. Technol. 2017, 51, 7861–7871. [Google Scholar] [CrossRef]
- Sturla Lompré, J.; Moleiro, P.; De Marchi, L.; Soares, A.M.V.M.; Pretti, C.; Chielini, F.; Pereira, E.; Freitas, R. Bioaccumulation and Ecotoxicological Responses of Clams Exposed to Terbium and Carbon Nanotubes: Comparison between Native (Ruditapes Decussatus) and Invasive (Ruditapes Philippinarum) Species. Sci. Total Environ. 2021, 784, 146914. [Google Scholar] [CrossRef]
- Henriques, B.; Teixeira, A.; Figueira, P.; Reis, A.T.; Almeida, J.; Vale, C.; Pereira, E. Simultaneous Removal of Trace Elements from Contaminated Waters by Living Ulva Lactuca. Sci. Total Environ. 2019, 652, 880–888. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.N.; Clarke, R.K. Permanova+ for Primer: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Freitas, R.; Coppola, F.; De Marchi, L.; Codella, V.; Pretti, C.; Chiellini, F.; Morelli, A.; Polese, G.; Soares, A.M.V.M.; Figueira, E. The Influence of Arsenic on the Toxicity of Carbon Nanoparticles in Bivalves. J. Hazard. Mater. 2018, 358, 484–493. [Google Scholar] [CrossRef]
- Anastopoulos, I.; Anagnostopoulos, V.A.; Bhatnagar, A.; Mitropoulos, A.C.; Kyzas, G.Z. A Review for Chromium Removal by Carbon Nanotubes. Chem. Ecol. 2017, 33, 572–588. [Google Scholar] [CrossRef]
- Atieh, M.A.; Bakather, O.Y.; Tawabini, B.S.; Bukhari, A.A.; Khaled, M.; Alharthi, M.; Fettouhi, M.; Abuilaiwi, F.A. Removal of Chromium (III) from Water by Using Modified and Nonmodified Carbon Nanotubes. J. Nanomater. 2010, 2010, 1–9. [Google Scholar] [CrossRef]
- Li, Y.H.; Ding, J.; Luan, Z.; Di, Z.; Zhu, Y.; Xu, C.; Wu, D.; Wei, B. Competitive Adsorption of Pb2+, Cu2+ and Cd 2+ Ions from Aqueous Solutions by Multiwalled Carbon Nanotubes. Carbon N. Y. 2003, 41, 2787–2792. [Google Scholar] [CrossRef]
- Lu, C.; Liu, C.; Su, F. Sorption Kinetics, Thermodynamics and Competition of Ni2+ from Aqueous Solutions onto Surface Oxidized Carbon Nanotubes. Desalination 2009, 249, 18–23. [Google Scholar] [CrossRef]
- Durier, G.; Nadalini, J.B.; Saint-Louis, R.; Genard, B.; Comeau, L.A.; Tremblay, R. Sensitivity to Oil Dispersants: Effects on the Valve Movements of the Blue Mussel Mytilus Edulis and the Giant Scallop Placopecten Magellanicus, in Sub-Arctic Conditions. Aquat. Toxicol. 2021, 234, 105797. [Google Scholar] [CrossRef]
- Britto, R.S.; Nascimento, J.P.; Serode, T.; Santos, A.P.; Soares, A.M.V.M.; Figueira, E.; Furtado, C.; Lima-Ventura, J.; Monserrat, J.M.; Freitas, R. The Effects of Co-Exposure of Graphene Oxide and Copper under Different PH Conditions in Manila Clam Ruditapes Philippinarum. Environ. Sci. Pollut. Res. 2020, 27, 30945–30956. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy Homeostasis as an Integrative Tool for Assessing Limits of Environmental Stress Tolerance in Aquatic Invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef]
- Gomes, T.; Pereira, C.G.; Cardoso, C.; Pinheiro, J.P.; Cancio, I.; Bebianno, M.J. Accumulation and Toxicity of Copper Oxide Nanoparticles in the Digestive Gland of Mytilus Galloprovincialis. Aquat. Toxicol. 2012, 118–119, 72–79. [Google Scholar] [CrossRef]
- Regoli, F.; Giuliani, M.E. Oxidative Pathways of Chemical Toxicity and Oxidative Stress Biomarkers in Marine Organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef]
- Matozzo, V.; Giacomazzo, M.; Finos, L.; Marin, M.G.; Bargelloni, L.; Milan, M. Can Ecological History Influence Immunomarker Responses and Antioxidant Enzyme Activities in Bivalves That Have Been Experimentally Exposed to Contaminants? A New Subject for Discussion in “Eco-Immunology” Studies. Fish Shellfish Immunol. 2013, 35, 126–135. [Google Scholar] [CrossRef]
- Munari, M.; Matozzo, V.; Gagné, F.; Chemello, G.; Riedl, V.; Finos, L.; Pastore, P.; Badocco, D.; Marin, M.G. Does Exposure to Reduced PH and Diclofenac Induce Oxidative Stress in Marine Bivalves? A Comparative Study with the Mussel Mytilus Galloprovincialis and the Clam Ruditapes Philippinarum. Environ. Pollut. 2018, 240, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Klimova, Y.S.; Chuiko, G.M.; Gapeeva, M.V.; Pesnya, D.S.; Ivanova, E.I. The Use of Oxidative Stress Parameters of Bivalve Mollusks Dreissena Polymorpha (Pallas, 1771) as Biomarkers for Ecotoxicological Assessment of Environment. Inl. Water Biol. 2019, 12, 88–95. [Google Scholar] [CrossRef]
- Coppola, F.; Bessa, A.; Henriques, B.; Russo, T.; Soares, A.M.V.M.; Figueira, E.; Marques, P.A.A.P.; Polese, G.; Di Cosmo, A.; Pereira, E.; et al. Oxidative Stress, Metabolic and Histopathological Alterations in Mussels Exposed to Remediated Seawater by GO-PEI after Contamination with Mercury. Comp. Biochem. Physiol.-Part A Mol. Integr. Physiol. 2020, 243, 110674. [Google Scholar] [CrossRef] [PubMed]
- Santonocito, M.; Salerno, B.; Trombini, C.; Tonini, F.; Pintado-Herrera, M.G.; Martínez-Rodríguez, G.; Blasco, J.; Lara-Martín, P.A.; Hampel, M. Stress under the Sun: Effects of Exposure to Low Concentrations of UV-Filter 4- Methylbenzylidene Camphor (4-MBC) in a Marine Bivalve Filter Feeder, the Manila Clam Ruditapes Philippinarum. Aquat. Toxicol. 2020, 221, 105418. [Google Scholar] [CrossRef] [PubMed]
- De Marchi, L.; Neto, V.; Pretti, C.; Figueira, E.; Chiellini, F.; Morelli, A.; Soares, A.M.V.M.; Freitas, R. The Impacts of Seawater Acidification on: Ruditapes Philippinarum Sensitivity to Carbon Nanoparticles. Environ. Sci. Nano 2017, 4, 1692–1704. [Google Scholar] [CrossRef]
- Mesquita, A.F.; Marques, S.M.; Marques, J.C.; Gonçalves, F.J.M.; Gonçalves, A.M.M. Copper Sulphate Impact on the Antioxidant Defence System of the Marine Bivalves Cerastoderma Edule and Scrobicularia Plana. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Völker, C.; Kämpken, I.; Boedicker, C.; Oehlmann, J.; Oetken, M. Toxicity of Silver Nanoparticles and Ionic Silver: Comparison of Adverse Effects and Potential Toxicity Mechanisms in the Freshwater Clam Sphaerium Corneum. Nanotoxicology 2015, 9, 677–685. [Google Scholar] [CrossRef]
- Cid, A.; Picado, A.; Correia, J.B.; Chaves, R.; Silva, H.; Caldeira, J.; de Matos, A.P.A.; Diniz, M.S. Oxidative Stress and Histological Changes Following Exposure to Diamond Nanoparticles in the Freshwater Asian Clam Corbicula Fluminea (Müller, 1774). J. Hazard. Mater. 2015, 284, 27–34. [Google Scholar] [CrossRef]
- Minetto, D.; Volpi Ghirardini, A.; Libralato, G. Saltwater Ecotoxicology of Ag, Au, CuO, TiO2, ZnO and C60 Engineered Nanoparticles: An Overview. Environ. Int. 2016, 92–93, 189–201. [Google Scholar] [CrossRef]
- Lionetto, M.G.; Caricato, R.; Calisi, A.; Giordano, M.E.; Schettino, T. Acetylcholinesterase as a Biomarker in Environmental and Occupational Medicine: New Insights and Future Perspectives. Biomed. Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Bocquené, G.; Galgani, F. Biological Effects of Contaminants: Cholinesterase Inhibition by Organophosphate and Carbamate Compounds. ICES Tech. Mar. Environ. Sci. 1998, 22, 1–19. [Google Scholar]
- Cunha, I.; Mangas-Ramirez, E.; Guilhermino, L. Effects of Copper and Cadmium on Cholinesterase and Glutathione S-Transferase Activities of Two Marine Gastropods (Monodonta Lineata and Nucella Lapillus). Comp. Biochem. Physiol.-C Toxicol. Pharmacol. 2007, 145, 648–657. [Google Scholar] [CrossRef]
- Turja, R.; Höher, N.; Snoeijs, P.; Baršiene, J.; Butrimavičiene, L.; Kuznetsova, T.; Kholodkevich, S.V.; Devier, M.H.; Budzinski, H.; Lehtonen, K.K. A Multibiomarker Approach to the Assessment of Pollution Impacts in Two Baltic Sea Coastal Areas in Sweden Using Caged Mussels (Mytilus Trossulus). Sci. Total Environ. 2014, 473–474, 398–409. [Google Scholar] [CrossRef]
- Mennillo, E.; Casu, V.; Tardelli, F.; De Marchi, L.; Freitas, R.; Pretti, C. Suitability of Cholinesterase of Polychaete Diopatra Neapolitana as Biomarker of Exposure to Pesticides: In Vitro Characterization. Comp. Biochem. Physiol. Part-C Toxicol. Pharmacol. 2017, 191, 152–159. [Google Scholar] [CrossRef]
- Moncaleano-Niño, A.M.; Luna-Acosta, A.; Gómez-Cubillos, M.C.; Villamil, L.; Ahrens, M.J. Cholinesterase Activity in the Cup Oyster Saccostrea Sp. Exposed to Chlorpyrifos, Imidacloprid, Cadmium and Copper. Ecotoxicol. Environ. Saf. 2018, 151, 242–254. [Google Scholar] [CrossRef]
- Bainy, D.A.C.; Helena, M.; Medeiros, G.D.; Mascio, P.D.; Almeida, E.A. De In Vivo Effects of Metals on the Acetylcholinesterase Activity of the Perna Perna Mussel’s Digestive Gland. Biotemas 2006, 19, 35–39. [Google Scholar]
- Rajkumar, J.S.I. Arsenic Induced Biochemical Changes in Perna Viridis as Potential Biomarkers in Metal Pollution. Asian J. Biochem. Pharm. Res. 2013, 3, 179–186. [Google Scholar]
- Liu, X.; Zhang, L.; You, L.; Cong, M.; Zhao, J.; Wu, H.; Li, C.; Liu, D.; Yu, J. Toxicological Responses to Acute Mercury Exposure for Three Species of Manila Clam Ruditapes Philippinarum by NMR-Based Metabolomics. Environ. Toxicol. Pharmacol. 2011, 31, 323–332. [Google Scholar] [CrossRef]
- Romani, R.; Antognelli, C.; Baldracchini, F.; De Santis, A.; Isani, G.; Giovannini, E.; Rosi, G. Increased Acetylcholinesterase Activities in Specimens of Sparus Auratus Exposed to Sublethal Copper Concentrations. Chem. Biol. Interact. 2003, 145, 321–329. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T14 | ||||
| CNTs | CNTs + Cr | |||
| Size (nm) | PDI | Size (nm) | PDI | |
| R. decussatus | I.d. | - | I.d. | - |
| 1215.4 | 0.49 | I.d. | - | |
| I.d. | - | 7387.2 | 3.10 | |
| R. philippinarum | I.d. | - | I.d. | - |
| I.d. | - | I.d. | - | |
| 4201.9 | 1.88 | 2279.8 | 1.02 | |
| T21 | ||||
| CNTs | CNTs + Cr | |||
| Size (nm) | PDI | Size (nm) | PDI | |
| R. decussatus | 9688.5 | 3.99 | I.d. | - |
| I.d. | - | 2243.6 | 0.85 | |
| I.d. | - | 4664.0 | 2.00 | |
| R. philippinarum | I.d. | - | I.d. | - |
| I.d. | - | 4844.0 | 1.99 | |
| 1237.5 | 0.75 | 5296.3 | 2.05 | |
| T28 | ||||
| CNTs | CNTs + Cr | |||
| Size (nm) | PDI | Size (nm) | PDI | |
| R. decussatus | I.d. | - | 2222.0 | 1.24 |
| I.d. | - | I.d. | - | |
| 4343.1 | 1.76 | I.d. | - | |
| R. philippinarum | 808.4 | 0.45 | 7200.7 | 2.84 |
| I.d. | - | 1096.5 | 0.43 | |
| 954.7 | 1.42 | 2209.0 | 0.85 | |
| Cr (mg/L) | ||
|---|---|---|
| R. decussatus | R. philippinarum | |
| CTL | <LOQ | <LOQ |
| Spiked | 3.21 ± 0.37 | 3.15 ± 0.53 |
| Cr (μg/g DW) | ||
|---|---|---|
| R. decussatus | R. philippinarum | |
| CTL | 1.36 ± 0.58 A | 1.17 ± 0.36 a |
| CNTs | 1.26 ± 0.19 A | 0.98 ± 0.06 a |
| Cr | 115.55 ± 15.80 B* | 48.13 ± 20.85 b* |
| CNTs + Cr | 118.51 ± 9.77 B* | 71.6 ± 17.12 b* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sturla Lompré, J.; De Marchi, L.; Pinto, J.; Soares, A.M.V.M.; Pretti, C.; Chielini, F.; Pereira, E.; Freitas, R. Effects of Carbon Nanoparticles and Chromium Combined Exposure in Native (Ruditapes decussatus) and Invasive (Ruditapes philippinarum) Clams. Nanomaterials 2023, 13, 690. https://doi.org/10.3390/nano13040690
Sturla Lompré J, De Marchi L, Pinto J, Soares AMVM, Pretti C, Chielini F, Pereira E, Freitas R. Effects of Carbon Nanoparticles and Chromium Combined Exposure in Native (Ruditapes decussatus) and Invasive (Ruditapes philippinarum) Clams. Nanomaterials. 2023; 13(4):690. https://doi.org/10.3390/nano13040690
Chicago/Turabian StyleSturla Lompré, Julieta, Lucia De Marchi, João Pinto, Amadeu M. V. M. Soares, Carlo Pretti, Federica Chielini, Eduarda Pereira, and Rosa Freitas. 2023. "Effects of Carbon Nanoparticles and Chromium Combined Exposure in Native (Ruditapes decussatus) and Invasive (Ruditapes philippinarum) Clams" Nanomaterials 13, no. 4: 690. https://doi.org/10.3390/nano13040690