
The Effect of Nanomaterials on DNA Methylation: A Review

Abstract
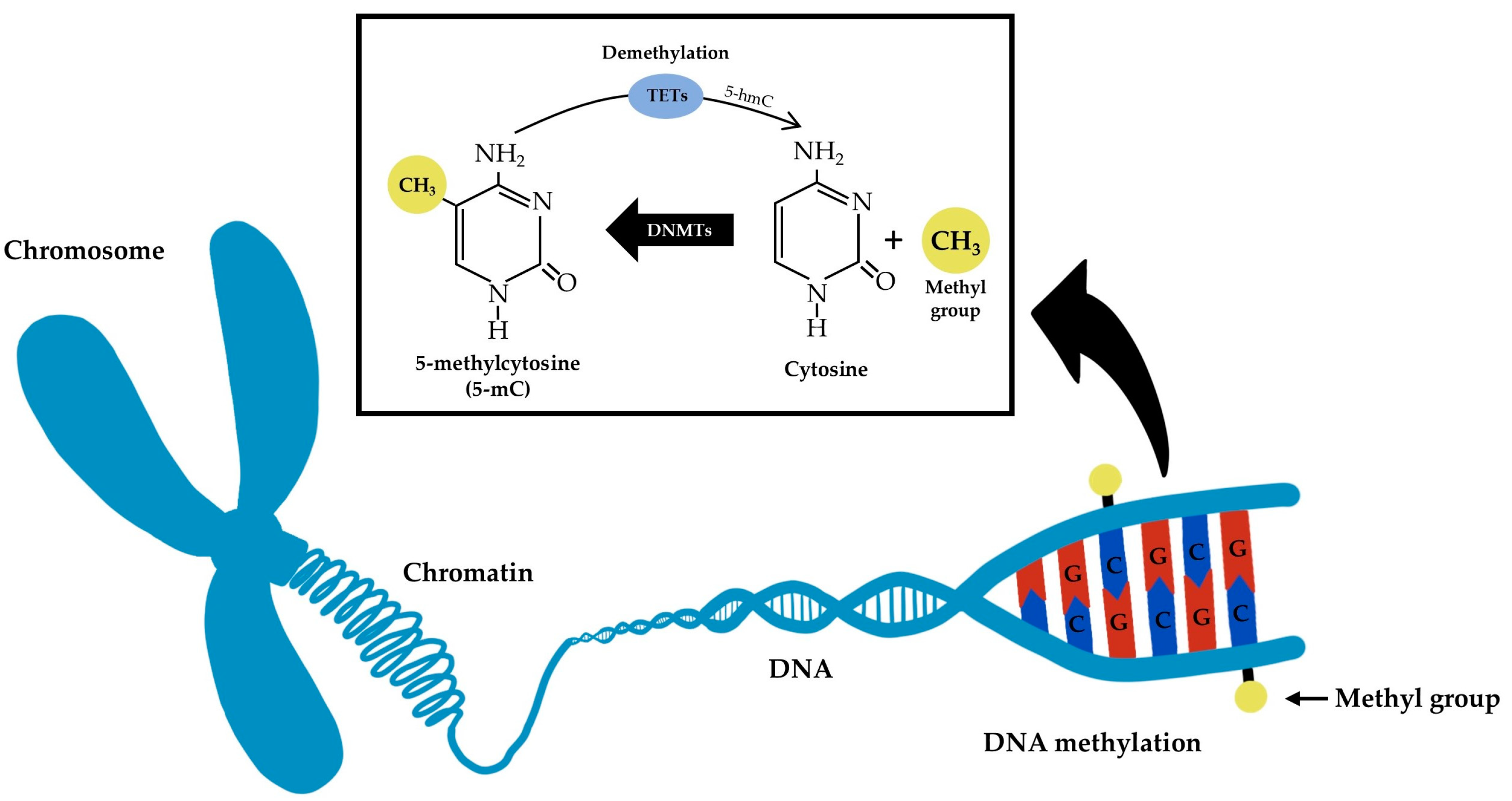
:1. Introduction
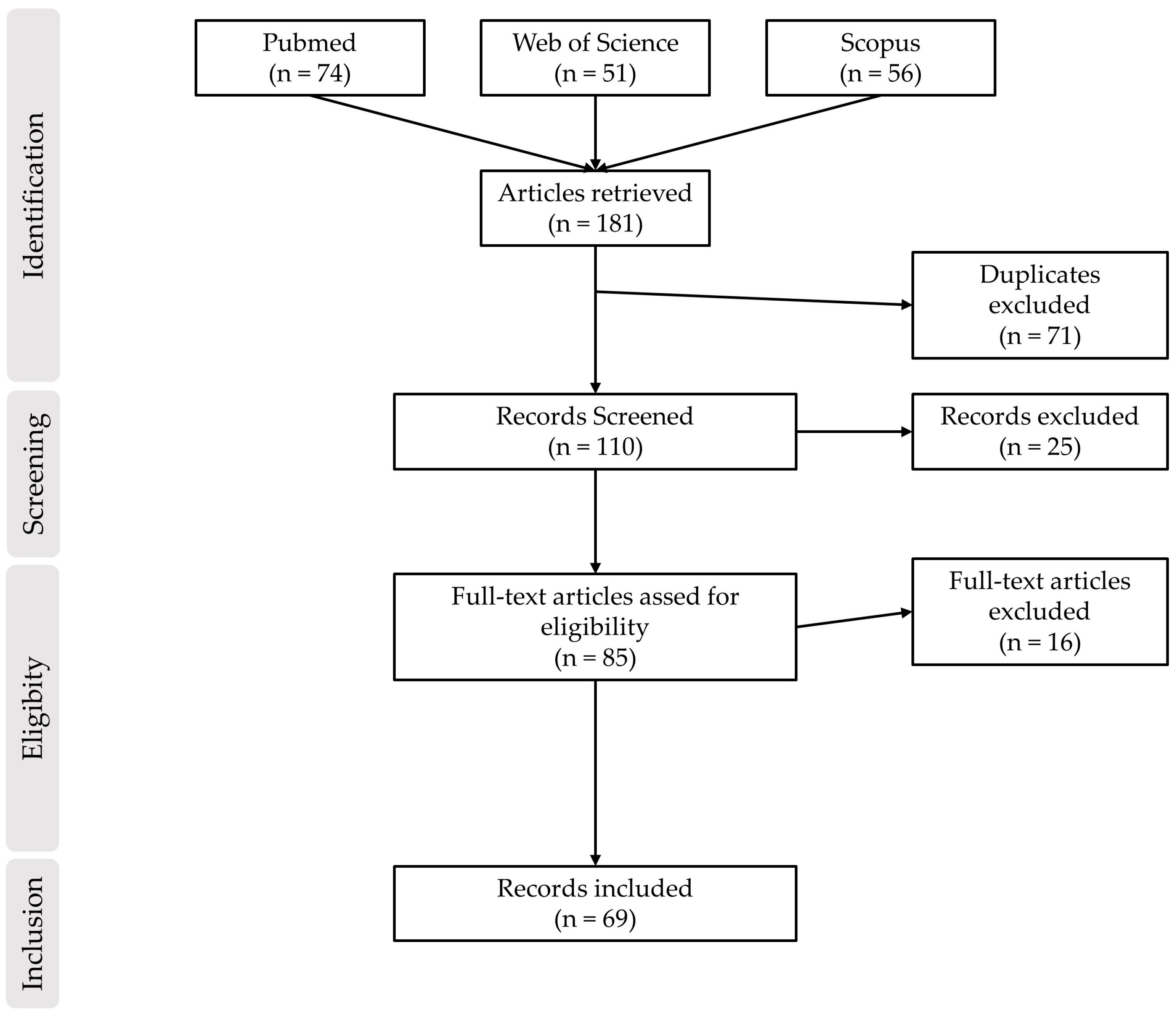
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Portela, A.; Esteller, M. Epigenetic Modifications and Human Disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. In Epigenetics in Allergy and Autoimmunity; Chang, C., Lu, Q., Eds.; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2020; pp. 3–55. ISBN 9789811534492. [Google Scholar]
- Baccarelli, A.; Bollati, V. Epigenetics and Environmental Chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, E.M.; Fry, R.C. Environmental Influences on the Epigenome: Exposure- Associated DNA Methylation in Human Populations. Annu. Rev. Public. Health 2018, 39, 309–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salata, O. Applications of Nanoparticles in Biology and Medicine. J. Nanobiotechnol. 2004, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Shafiq, M.; Anjum, S.; Hano, C.; Anjum, I.; Abbasi, B.H. An Overview of the Applications of Nanomaterials and Nanodevices in the Food Industry. Foods 2020, 9, 148. [Google Scholar] [CrossRef] [Green Version]
- Akgöl, S.; Ulucan-Karnak, F.; Kuru, C.İ.; Kuşat, K. The Usage of Composite Nanomaterials in Biomedical Engineering Applications. Biotechnol. Bioeng. 2021, 118, 2906–2922. [Google Scholar] [CrossRef]
- Siddique, S.; Chow, J.C.L. Application of Nanomaterials in Biomedical Imaging and Cancer Therapy. Nanomaterials 2020, 10, 1700. [Google Scholar] [CrossRef]
- Saleh, T.A. Nanomaterials: Classification, Properties, and Environmental Toxicities. Environ. Technol. Innov. 2020, 20, 101067. [Google Scholar] [CrossRef]
- European Commission. Commission Recommendation of 10.6.2022 on the Definition of Nanomaterial; European Commission: Brussels, Belgium, 2018. [Google Scholar]
- ISO/TR 11360:2010; Nanotechnologies-Methodology for the Classification and Categorization of Nanomaterials. International Organization for Standardization: Geneva, Switzerland, 2010.
- Li, X.; Liu, W.; Sun, L.; Aifantis, K.E.; Yu, B.; Fan, Y.; Feng, Q.; Cui, F.; Watari, F. Effects of Physicochemical Properties of Nanomaterials on Their Toxicity. J. Biomed. Mater. Res. Part A 2015, 103, 2499–2507. [Google Scholar] [CrossRef]
- Gatoo, M.A.; Naseem, S.; Arfat, M.Y.; Mahmood Dar, A.; Qasim, K.; Zubair, S. Physicochemical Properties of Nanomaterials: Implication in Associated Toxic Manifestations. Biomed. Res. Int. 2014, 2014, 498420. [Google Scholar] [CrossRef] [Green Version]
- Landsiedel, R.; Sauer, U.G.; Ma-Hock, L.; Schnekenburger, J.; Wiemann, M. Pulmonary Toxicity of Nanomaterials: A Critical Comparison of Published in vitro Assays and in vivo Inhalation or Instillation Studies. Nanomedicine 2014, 9, 2557–2585. [Google Scholar] [CrossRef] [PubMed]
- Baan, R.; Straif, K.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Cogliano, V. WHO International Agency for Research on Cancer Monograph Working Group Carcinogenicity of Carbon Black, Titanium Dioxide, and Talc. Lancet Oncol. 2006, 7, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Han, J.W.; Eppakayala, V.; Jeyaraj, M.; Kim, J.-H. Cytotoxicity of Biologically Synthesized Silver Nanoparticles in MDA-MB-231 Human Breast Cancer Cells. Biomed. Res. Int. 2013, 2013, 535796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.-K.; Gurunathan, S.; Kang, M.-H.; Han, J.W.; Das, J.; Choi, Y.-J.; Kwon, D.-N.; Cho, S.-G.; Park, C.; Seo, H.G.; et al. Hypoxia-Mediated Autophagic Flux Inhibits Silver Nanoparticle-Triggered Apoptosis in Human Lung Cancer Cells. Sci. Rep. 2016, 6, 21688. [Google Scholar] [CrossRef] [Green Version]
- Grosse, Y.; Loomis, D.; Guyton, K.Z.; Lauby-Secretan, B.; Ghissassi, F.E.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Scoccianti, C.; Mattock, H.; et al. Carcinogenicity of Fluoro-Edenite, Silicon Carbide Fibres and Whiskers, and Carbon Nanotubes. Lancet Oncol. 2014, 15, 1427–1428. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.; Marks, L.; May, G.H.W.; Wilson, J.B. The Genetic Basis of Disease. Essays Biochem. 2018, 62, 643–723. [Google Scholar] [CrossRef] [Green Version]
- Engvall, E.; Perlmann, P. Enzyme-Linked Immunosorbent Assay, Elisa: III. Quantitation of Specific Antibodies by Enzyme-Labeled Anti-Immunoglobulin in Antigen-Coated Tubes. J. Immunol. 1972, 109, 129–135. [Google Scholar] [CrossRef]
- Kuo, K.C.; McCune, R.A.; Gehrke, C.W.; Midgett, R.; Ehrlich, M. Quantitative Reversed-Phase High Performance Liquid Chromatographic Determination of Major and Modified Deoxyribonucleosides in DNA. Nucleic Acids Res. 1980, 8, 4763–4776. [Google Scholar] [CrossRef] [Green Version]
- Jemal, M. High-Throughput Quantitative Bioanalysis by LC/MS/MS. Biomed. Chromatogr. 2000, 14, 422–429. [Google Scholar] [CrossRef]
- Yong, W.-S.; Hsu, F.-M.; Chen, P.-Y. Profiling Genome-Wide DNA Methylation. Epigenetics Chromatin 2016, 9, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blohm, D.H.; Guiseppi-Elie, A. New Developments in Microarray Technology. Curr. Opin. Biotechnol. 2001, 12, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Metzker, M.L. Sequencing Technologies—The next Generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.G.; Graff, J.R.; Myöhänen, S.; Nelkin, B.D.; Baylin, S.B. Methylation-Specific PCR: A Novel PCR Assay for Methylation Status of CpG Islands. Proc. Natl. Acad. Sci. USA 1996, 93, 9821–9826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, S.-H.; Wu, W.-T.; Liao, H.-Y.; Chen, C.-Y.; Tsai, C.-Y.; Jung, W.-T.; Lee, H.-L. Global DNA Methylation and Oxidative Stress Biomarkers in Workers Exposed to Metal Oxide Nanoparticles. J. Hazard. Mater. 2017, 331, 329–335. [Google Scholar] [CrossRef]
- Blanco, J.; Lafuente, D.; Gómez, M.; García, T.; Domingo, J.L.; Sánchez, D.J. Polyvinyl Pyrrolidone-Coated Silver Nanoparticles in a Human Lung Cancer Cells: Time- and Dose-Dependent Influence over P53 and Caspase-3 Protein Expression and Epigenetic Effects. Arch. Toxicol. 2017, 91, 651–666. [Google Scholar] [CrossRef]
- Maki, A.; Lin, Y.; Aoyama, M.; Sato, K.; Gao, J.-Q.; Tsujino, H.; Nagano, K.; Higashisaka, K.; Tsutsumi, Y. Silver Nanoparticles Induce DNA Hypomethylation through Proteasome-Mediated Degradation of DNA Methyltransferase 1. Biol. Pharm. Bull. 2020, 43, 1924–1930. [Google Scholar] [CrossRef]
- Gliga, A.R.; Di Bucchianico, S.; Lindvall, J.; Fadeel, B.; Karlsson, H.L. RNA-Sequencing Reveals Long-Term Effects of Silver Nanoparticles on Human Lung Cells. Sci. Rep. 2018, 8, 6668. [Google Scholar] [CrossRef] [Green Version]
- Thai, S.-F.; Jones, C.P.; Robinette, B.L.; Ren, H.; Vallanat, B.; Fisher, A.A.; Kitchin, K.T. Effects of Silver Nanoparticles and Silver Nitrate on MRNA and MicroRNA Expression in Human Hepatocellular Carcinoma Cells (HepG2). J. Nanosci. Nanotechnol. 2021, 21, 5414–5428. [Google Scholar] [CrossRef]
- Mytych, J.; Zebrowski, J.; Lewinska, A.; Wnuk, M. Prolonged Effects of Silver Nanoparticles on P53/P21 Pathway-Mediated Proliferation, DNA Damage Response, and Methylation Parameters in HT22 Hippocampal Neuronal Cells. Mol. Neurobiol. 2017, 54, 1285–1300. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Sheng, F.; Wang, X.; Zhang, Z.; Qi, S.; Chen, L. Early Epigenetic Responses in the Genomic DNA Methylation Fingerprints in Cells in Response to Sublethal Exposure of Silver Nanoparticles. Front. Bioeng. Biotechnol. 2022, 10, 927036. [Google Scholar] [CrossRef] [PubMed]
- González-Palomo, A.K.; Saldaña-Villanueva, K.; Cortés-García, J.D.; Fernández-Macias, J.C.; Méndez-Rodríguez, K.B.; Pérez Maldonado, I.N. Effect of Silver Nanoparticles (AgNPs) Exposure on MicroRNA Expression and Global DNA Methylation in Endothelial Cells EA.Hy926. Environ. Toxicol. Pharmacol. 2021, 81, 103543. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Qasim, M.; Park, C.; Yoo, H.; Choi, D.Y.; Song, H.; Park, C.; Kim, J.-H.; Hong, K. Cytotoxicity and Transcriptomic Analysis of Silver Nanoparticles in Mouse Embryonic Fibroblast Cells. Int. J. Mol. Sci. 2018, 19, 3618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzóska, K.; Grądzka, I.; Kruszewski, M. Silver, Gold, and Iron Oxide Nanoparticles Alter MiRNA Expression but Do Not Affect DNA Methylation in HepG2 Cells. Materials 2019, 12, 1038. [Google Scholar] [CrossRef] [Green Version]
- Gambelunghe, A.; Giovagnoli, S.; Di Michele, A.; Boncompagni, S.; Dell’Omo, M.; Leopold, K.; Iavicoli, I.; Talesa, V.N.; Antognelli, C. Redox-Sensitive Glyoxalase 1 Up-Regulation Is Crucial for Protecting Human Lung Cells from Gold Nanoparticles Toxicity. Antioxidants 2020, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.-T.; Dheen, S.T.; Yip, W.-C.G.; Ong, C.-N.; Bay, B.-H.; Lanry Yung, L.-Y. The Induction of Epigenetic Regulation of PROS1 Gene in Lung Fibroblasts by Gold Nanoparticles and Implications for Potential Lung Injury. Biomaterials 2011, 32, 7609–7615. [Google Scholar] [CrossRef] [PubMed]
- Sooklert, K.; Nilyai, S.; Rojanathanes, R.; Jindatip, D.; Sae-Liang, N.; Kitkumthorn, N.; Mutirangura, A.; Sereemaspun, A. N-Acetylcysteine Reverses the Decrease of DNA Methylation Status Caused by Engineered Gold, Silicon, and Chitosan Nanoparticles. Int. J. Nanomed. 2019, 14, 4573–4587. [Google Scholar] [CrossRef] [Green Version]
- Senut, M.-C.; Zhang, Y.; Liu, F.; Sen, A.; Ruden, D.M.; Mao, G. Size-Dependent Toxicity of Gold Nanoparticles on Human Embryonic Stem Cells and Their Neural Derivatives. Small 2016, 12, 631–646. [Google Scholar] [CrossRef] [Green Version]
- Biola-Clier, M.; Beal, D.; Caillat, S.; Libert, S.; Armand, L.; Herlin-Boime, N.; Sauvaigo, S.; Douki, T.; Carriere, M. Comparison of the DNA Damage Response in BEAS-2B and A549 Cells Exposed to Titanium Dioxide Nanoparticles. Mutagenesis 2017, 32, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Bai, W.; Chen, Y.; Gao, A. Cross Talk between Poly(ADP-Ribose) Polymerase 1 Methylation and Oxidative Stress Involved in the Toxic Effect of Anatase Titanium Dioxide Nanoparticles. Int. J. Nanomed. 2015, 10, 5561–5569. [Google Scholar] [CrossRef] [Green Version]
- Stoccoro, A.; Di Bucchianico, S.; Coppedè, F.; Ponti, J.; Uboldi, C.; Blosi, M.; Delpivo, C.; Ortelli, S.; Costa, A.L.; Migliore, L. Multiple Endpoints to Evaluate Pristine and Remediated Titanium Dioxide Nanoparticles Genotoxicity in Lung Epithelial A549 Cells. Toxicol. Lett. 2017, 276, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Guo, Y.; Wu, S.; Lv, Z.; Zhang, Q.; Ke, Y. Titanium Dioxide Nanoparticles Induce Size-Dependent Cytotoxicity and Genomic DNA Hypomethylation in Human Respiratory Cells. RSC Adv. 2017, 7, 23560–23572. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, M.; Öner, D.; Duca, R.-C.; Cokic, S.M.; Seys, S.; Kerkhofs, S.; Van Landuyt, K.; Hoet, P.; Godderis, L. Cyto-Genotoxic and DNA Methylation Changes Induced by Different Crystal Phases of TiO2-Np in Bronchial Epithelial (16-HBE) Cells. Mutat. Res. 2017, 796, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sierra, M.I.; Rubio, L.; Bayón, G.F.; Cobo, I.; Menendez, P.; Morales, P.; Mangas, C.; Urdinguio, R.G.; Lopez, V.; Valdes, A.; et al. DNA Methylation Changes in Human Lung Epithelia Cells Exposed to Multi-Walled Carbon Nanotubes. Nanotoxicology 2017, 11, 857–870. [Google Scholar] [CrossRef]
- Pogribna, M.; Koonce, N.A.; Mathew, A.; Word, B.; Patri, A.K.; Lyn-Cook, B.; Hammons, G. Effect of Titanium Dioxide Nanoparticles on DNA Methylation in Multiple Human Cell Lines. Nanotoxicology 2020, 14, 534–553. [Google Scholar] [CrossRef]
- Malakootian, M.; Nasiri, A.; Osornio-Vargas, A.R.; Faraji, M. Effect of Titanium Dioxide Nanoparticles on DNA Methylation of Human Peripheral Blood Mononuclear Cells. Toxicol. Res. 2021, 10, 1045–1051. [Google Scholar] [CrossRef]
- Lu, X.; Miousse, I.R.; Pirela, S.V.; Melnyk, S.; Koturbash, I.; Demokritou, P. Short-Term Exposure to Engineered Nanomaterials Affects Cellular Epigenome. Nanotoxicology 2016, 10, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Patil, N.A.; Gade, W.N.; Deobagkar, D.D. Epigenetic Modulation upon Exposure of Lung Fibroblasts to TiO2 and ZnO Nanoparticles: Alterations in DNA Methylation. Int. J. Nanomed. 2016, 11, 4509–4519. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.R.; Ordaz, J.; Lo, C.-L.; Damayanti, N.P.; Zhou, F.; Irudayaraj, J. From the Cover: Zinc Oxide Nanoparticles-Induced Reactive Oxygen Species Promotes Multimodal Cyto- and Epigenetic Toxicity. Toxicol. Sci. 2017, 156, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Thai, S.-F.; Jones, C.P.; Robinette, B.L.; Ren, H.; Vallant, B.; Fisher, A.; Kitchin, K.T. Effects of Copper Nanoparticles on MRNA and Small RNA Expression in Human Hepatocellular Carcinoma (HepG2) Cells. J. Nanosci. Nanotechnol. 2021, 21, 5083–5098. [Google Scholar] [CrossRef]
- Perreault, F.; Pedroso Melegari, S.; Henning da Costa, C.; de Oliveira Franco Rossetto, A.L.; Popovic, R.; Gerson Matias, W. Genotoxic Effects of Copper Oxide Nanoparticles in Neuro 2A Cell Cultures. Sci. Total Environ. 2012, 441, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Bonadio, R.S.; da Cunha, M.C.P.C.; Longo, J.P.F.; Azevedo, R.B.; PoÇas-Fonseca, M.J. Exposure to Maghemite Nanoparticles Induces Epigenetic Alterations in Human Submandibular Gland Cells. J. Nanosci. Nanotechnol. 2020, 20, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Uboldi, C.; Sanles Sobrido, M.; Bernard, E.; Tassistro, V.; Herlin-Boime, N.; Vrel, D.; Garcia-Argote, S.; Roche, S.; Magdinier, F.; Dinescu, G.; et al. In vitro Analysis of the Effects of ITER-Like Tungsten Nanoparticles: Cytotoxicity and Epigenotoxicity in BEAS-2B Cells. Nanomaterials 2019, 9, 1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tian, X.; Zhang, J.; Tan, L.; Ouyang, N.; Jia, B.; Chen, C.; Ge, C.; Li, J. Postchronic Single-Walled Carbon Nanotube Exposure Causes Irreversible Malignant Transformation of Human Bronchial Epithelial Cells through DNA Methylation Changes. ACS Nano 2021, 15, 7094–7104. [Google Scholar] [CrossRef] [PubMed]
- Öner, D.; Ghosh, M.; Coorens, R.; Bové, H.; Moisse, M.; Lambrechts, D.; Ameloot, M.; Godderis, L.; Hoet, P.H.M. Induction and Recovery of CpG Site Specific Methylation Changes in Human Bronchial Cells after Long-Term Exposure to Carbon Nanotubes and Asbestos. Environ. Int. 2020, 137, 105530. [Google Scholar] [CrossRef]
- Öner, D.; Ghosh, M.; Bové, H.; Moisse, M.; Boeckx, B.; Duca, R.C.; Poels, K.; Luyts, K.; Putzeys, E.; Van Landuydt, K.; et al. Differences in MWCNT- and SWCNT-Induced DNA Methylation Alterations in Association with the Nuclear Deposition. Part. Fibre Toxicol. 2018, 15, 11. [Google Scholar] [CrossRef]
- Öner, D.; Moisse, M.; Ghosh, M.; Duca, R.C.; Poels, K.; Luyts, K.; Putzeys, E.; Cokic, S.M.; Van Landuyt, K.; Vanoirbeek, J.; et al. Epigenetic Effects of Carbon Nanotubes in Human Monocytic Cells. Mutagenesis 2017, 32, 181–191. [Google Scholar] [CrossRef]
- Saarimäki, L.A.; Kinaret, P.A.S.; Scala, G.; del Giudice, G.; Federico, A.; Serra, A.; Greco, D. Toxicogenomics Analysis of Dynamic Dose-Response in Macrophages Highlights Molecular Alterations Relevant for Multi-Walled Carbon Nanotube-Induced Lung Fibrosis. NanoImpact 2020, 20, 100274. [Google Scholar] [CrossRef]
- Chatterjee, N.; Yang, J.; Yoon, D.; Kim, S.; Joo, S.-W.; Choi, J. Differential Crosstalk between Global DNA Methylation and Metabolomics Associated with Cell Type Specific Stress Response by Pristine and Functionalized MWCNT. Biomaterials 2017, 115, 167–180. [Google Scholar] [CrossRef]
- Scala, G.; Kinaret, P.; Marwah, V.; Sund, J.; Fortino, V.; Greco, D. Multi-Omics Analysis of Ten Carbon Nanomaterials Effects Highlights Cell Type Specific Patterns of Molecular Regulation and Adaptation. NanoImpact 2018, 11, 99–108. [Google Scholar] [CrossRef]
- Li, J.; Tian, M.; Cui, L.; Dwyer, J.; Fullwood, N.J.; Shen, H.; Martin, F.L. Low-Dose Carbon-Based Nanoparticle-Induced Effects in A549 Lung Cells Determined by Biospectroscopy Are Associated with Increases in Genomic Methylation. Sci. Rep. 2016, 6, 20207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, R.F.; Soto Fernández, A.Y.; Bousquets Muñoz, P.; Sierra, M.I.; Tejedor, J.R.; Morales-Sánchez, P.; Valdés, A.F.; Santamaría, R.; Blanco, C.; Torrecillas, R.; et al. No Genome-Wide DNA Methylation Changes Found Associated with Medium-Term Reduced Graphene Oxide Exposure in Human Lung Epithelial Cells. Epigenetics 2020, 15, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, N.; Yang, J.; Choi, J. Differential Genotoxic and Epigenotoxic Effects of Graphene Family Nanomaterials (GFNs) in Human Bronchial Epithelial Cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2016, 798–799, 1–10. [Google Scholar] [CrossRef]
- Ku, T.; Hao, F.; Yang, X.; Rao, Z.; Liu, Q.S.; Sang, N.; Faiola, F.; Zhou, Q.; Jiang, G. Graphene Quantum Dots Disrupt Embryonic Stem Cell Differentiation by Interfering with the Methylation Level of Sox2. Environ. Sci. Technol. 2021, 55, 3144–3155. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Liu, J.; Li, X.; Zhou, G.; Zhang, Y.; Gao, L.; Zhao, Y.; Zhou, X. The Effect of SiNPs on DNA Methylation of Genome in Mouse Spermatocytes. Environ. Sci. Pollut. Res. Int. 2021, 28, 43684–43697. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Li, Q.; Jiang, L.; Guo, C.; Li, Y.; Yu, Y.; Li, Y.; Duan, J.; Sun, Z. DNA Hypermethylation of CREB3L1 and Bcl-2 Associated with the Mitochondrial-Mediated Apoptosis via PI3K/Akt Pathway in Human BEAS-2B Cells Exposure to Silica Nanoparticles. PLoS ONE 2016, 11, e0158475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, C.; Tao, G.; Yang, L.; Liu, J.; Liu, Q.; Li, W.; Zhuang, Z. Methylation of PARP-1 Promoter Involved in the Regulation of Nano-SiO2-Induced Decrease of PARP-1 MRNA Expression. Toxicol. Lett. 2012, 209, 264–269. [Google Scholar] [CrossRef]
- Gong, C.; Tao, G.; Yang, L.; Liu, J.; Liu, Q.; Zhuang, Z. SiO2 Nanoparticles Induce Global Genomic Hypomethylation in HaCaT Cells. Biochem. Biophys. Res. Commun. 2010, 397, 397–400. [Google Scholar] [CrossRef]
- Seidel, C.; Kirsch, A.; Fontana, C.; Visvikis, A.; Remy, A.; Gaté, L.; Darne, C.; Guichard, Y. Epigenetic Changes in the Early Stage of Silica-Induced Cell Transformation. Nanotoxicology 2017, 11, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Motawi, T.K.; El-Maraghy, S.A.; Sabry, D.; Nady, O.M.; Senousy, M.A. Cromolyn Chitosan Nanoparticles Reverse the DNA Methylation of RASSF1A and P16 Genes and Mitigate DNMT1 and METTL3 Expression in Breast Cancer Cell Line and Tumor Xenograft Model in Mice. Chem. Biol. Interact. 2022, 365, 110094. [Google Scholar] [CrossRef]
- Abbaszadeh, S.; Rashidipour, M.; Khosravi, P.; Shahryarhesami, S.; Ashrafi, B.; Kaviani, M.; Moradi Sarabi, M. Biocompatibility, Cytotoxicity, Antimicrobial and Epigenetic Effects of Novel Chitosan-Based Quercetin Nanohydrogel in Human Cancer Cells. Int. J. Nanomed. 2020, 15, 5963–5975. [Google Scholar] [CrossRef] [PubMed]
- Chamani, F.; Sadeghizadeh, M.; Masoumi, M.; Babashah, S. Evaluation of MiR-34 Family and DNA Methyltransferases 1, 3A, 3B Gene Expression Levels in Hepatocellular Carcinoma Following Treatment with Dendrosomal Nanocurcumin. Asian Pac. J. Cancer Prev. 2016, 17, 219–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirela, S.V.; Miousse, I.R.; Lu, X.; Castranova, V.; Thomas, T.; Qian, Y.; Bello, D.; Kobzik, L.; Koturbash, I.; Demokritou, P. Effects of Laser Printer-Emitted Engineered Nanoparticles on Cytotoxicity, Chemokine Expression, Reactive Oxygen Species, DNA Methylation, and DNA Damage: A Comprehensive in vitro Analysis in Human Small Airway Epithelial Cells, Macrophages, and Lymphoblasts. Environ. Health Perspect. 2016, 124, 210–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.-G.; Cai, H.-Q.; Wang, J.-L.; Mesalam, A.; Reza, A.M.M.T.; Li, L.; Chen, L.; Qian, C. Graphene Oxide–Silver Nanoparticle Nanocomposites Induce Oxidative Stress and Aberrant Methylation in Caprine Fetal Fibroblast Cells. Cells 2021, 10, 682. [Google Scholar] [CrossRef]
- Wen, L.; Li, M.; Lin, X.; Li, Y.; Song, H.; Chen, H. AgNPs Aggravated Hepatic Steatosis, Inflammation, Oxidative Stress, and Epigenetic Changes in Mice With NAFLD Induced by HFD. Front. Bioeng. Biotechnol. 2022, 10, 912178. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Park, J.-H.; Choi, Y.-J.; Kang, M.-H.; Gurunathan, S.; Kim, J.-H. Silver Nanoparticles Cause Complications in Pregnant Mice. Int. J. Nanomed. 2015, 10, 7057–7071. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.-H.; Guan, P.; Zhang, T.; Lu, C.; Li, G.; Liu, J.-X. Silver Nanoparticles Impair Zebrafish Skeletal and Cardiac Myofibrillogenesis and Sarcomere Formation. Aquat. Toxicol. 2018, 200, 102–113. [Google Scholar] [CrossRef]
- Tabish, A.M.; Poels, K.; Byun, H.-M.; Luyts, K.; Baccarelli, A.A.; Martens, J.; Kerkhofs, S.; Seys, S.; Hoet, P.; Godderis, L. Changes in DNA Methylation in Mouse Lungs after a Single Intra-Tracheal Administration of Nanomaterials. PLoS ONE 2017, 12, e0169886. [Google Scholar] [CrossRef] [Green Version]
- Kunovac, A.; Hathaway, Q.A.; Pinti, M.V.; Goldsmith, W.T.; Durr, A.J.; Fink, G.K.; Nurkiewicz, T.R.; Hollander, J.M. ROS Promote Epigenetic Remodeling and Cardiac Dysfunction in Offspring Following Maternal Engineered Nanomaterial (ENM) Exposure. Part. Fibre Toxicol. 2019, 16, 24. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Guo, Y.; Ye, H.; Huang, K.; Lv, Z.; Ke, Y. Different Effects of Titanium Dioxide Nanoparticles Instillation in Young and Adult Mice on DNA Methylation Related with Lung Inflammation and Fibrosis. Ecotoxicol. Environ. Saf. 2019, 176, 1–10. [Google Scholar] [CrossRef]
- Ognik, K.; Cholewińska, E.; Juśkiewicz, J.; Zduńczyk, Z.; Tutaj, K.; Szlązak, R. The Effect of Copper Nanoparticles and Copper (II) Salt on Redox Reactions and Epigenetic Changes in a Rat Model. J. Anim. Physiol. Anim. Nutr. 2019, 103, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.M.; Gosens, I.; Williams, A.; Farcal, L.; Pantano, D.; Brown, D.M.; Stone, V.; Cassee, F.R.; Halappanavar, S.; Fadeel, B. Transcriptional Profiling Reveals Gene Expression Changes Associated with Inflammation and Cell Proliferation Following Short-Term Inhalation Exposure to Copper Oxide Nanoparticles. J. Appl. Toxicol. 2018, 38, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Miousse, I.R.; Pirela, S.V.; Moore, J.K.; Melnyk, S.; Koturbash, I.; Demokritou, P. In vivo Epigenetic Effects Induced by Engineered Nanomaterials: A Case Study of Copper Oxide and Laser Printer-Emitted Engineered Nanoparticles. Nanotoxicology 2016, 10, 629–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bicho, R.C.; Faustino, A.M.R.; Rêma, A.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Confirmatory Assays for Transient Changes of Omics in Soil Invertebrates–Copper Materials in a Multigenerational Exposure. J. Hazard. Mater. 2021, 402, 123500. [Google Scholar] [CrossRef]
- Bicho, R.C.; Roelofs, D.; Mariën, J.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Epigenetic Effects of (Nano)Materials in Environmental Species–Cu Case Study in Enchytraeus Crypticus. Environ. Int. 2020, 136, 105447. [Google Scholar] [CrossRef]
- Zheng, J.; Wang, J.; Qin, X.; Li, K.; Gao, Q.; Yang, M.; Liu, H.; Li, S.; Chang, X.; Sun, Y. LncRNA HOTAIRM1 Involved in Nano NiO-Induced Pulmonary Fibrosis via Regulating PRKCB DNA Methylation-Mediated JNK/c-Jun Pathway. Toxicol. Sci. 2022, 190, 64–78. [Google Scholar] [CrossRef]
- Shoeb, M.; Kodali, V.K.; Farris, B.Y.; Bishop, L.M.; Meighan, T.G.; Salmen, R.; Eye, T.; Friend, S.; Schwegler-Berry, D.; Roberts, J.R.; et al. Oxidative Stress, DNA Methylation, and Telomere Length Changes in Peripheral Blood Mononuclear Cells after Pulmonary Exposure to Metal-Rich Welding Nanoparticles. NanoImpact 2017, 5, 61–69. [Google Scholar] [CrossRef]
- Gorrochategui, E.; Li, J.; Fullwood, N.J.; Ying, G.-G.; Tian, M.; Cui, L.; Shen, H.; Lacorte, S.; Tauler, R.; Martin, F.L. Diet-Sourced Carbon-Based Nanoparticles Induce Lipid Alterations in Tissues of Zebrafish (Danio Rerio) with Genomic Hypermethylation Changes in Brain. Mutagenesis 2017, 32, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Lin, W.; Lin, B.; Wu, K.; Fan, H.; Yu, Y. Persistent DNA Methylation Changes in Zebrafish Following Graphene Quantum Dots Exposure in Surface Chemistry-Dependent Manner. Ecotoxicol. Environ. Saf. 2019, 169, 370–375. [Google Scholar] [CrossRef]
- Li, M.; Zhu, J.; Fang, H.; Wang, M.; Wang, Q.; Zhou, B. Coexposure to Environmental Concentrations of Cis-Bifenthrin and Graphene Oxide: Adverse Effects on the Nervous System during Metamorphic Development of Xenopus Laevis. J. Hazard. Mater. 2020, 381, 120995. [Google Scholar] [CrossRef]
- Zhou, W.; Tian, D.; He, J.; Yan, X.; Zhao, J.; Yuan, X.; Peng, S. Prolonged Exposure to Carbon Nanoparticles Induced Methylome Remodeling and Gene Expression in Zebrafish Heart. J. Appl. Toxicol. 2019, 39, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Li, J.; Lou, H.; Cao, Z.; Fan, X. Genome-Wide DNA Methylation Alterations and Potential Risk Induced by Subacute and Subchronic Exposure to Food-Grade Nanosilica in Mice. ACS Nano 2021, 15, 8225–8243. [Google Scholar] [CrossRef] [PubMed]
- Rossnerova, A.; Honkova, K.; Chvojkova, I.; Pelclova, D.; Zdimal, V.; Hubacek, J.A.; Lischkova, L.; Vlckova, S.; Ondracek, J.; Dvorackova, S.; et al. Individual DNA Methylation Pattern Shifts in Nanoparticles-Exposed Workers Analyzed in Four Consecutive Years. Int. J. Mol. Sci. 2021, 22, 7834. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-W.; Bae, S.-H.; Jeong, J.-W.; Kim, S.-H.; Kim, K.-W. Hypoxia-Inducible Factor (HIF-1)Alpha: Its Protein Stability and Biological Functions. Exp. Mol. Med. 2004, 36, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ventura, C.; Torres, V.; Vieira, L.; Gomes, B.; Rodrigues, A.S.; Rueff, J.; Penque, D.; Silva, M.J. New “Omics” Approaches as Tools to Explore Mechanistic Nanotoxicology. In Nanotoxicology in Safety Assessment of Nanomaterials; Louro, H., Silva, M.J., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2022; pp. 179–194. ISBN 978-3-030-88071-2. [Google Scholar]
- Schneider, K.; Schwarz, M.; Burkholder, I.; Kopp-Schneider, A.; Edler, L.; Kinsner-Ovaskainen, A.; Hartung, T.; Hoffmann, S. “ToxRTool”, a new tool to assess the reliability of toxicological data. Toxicol. Lett. 2009, 189, 138–144. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Nanomaterials, Concentration-Range | Cell Model | Methodology | Main Conclusions | References |
|---|---|---|---|---|
| Metal-based nanomaterials | ||||
| AgNPs, 10–200 µg/mL for 72 h. | A549 cells | Global DNA methylation (ELISA) | High concentrations of AgNPs for 72 h induced higher levels of global DNA methylation. | Blanco et al., 2017 [29] |
| AgNPs with a diameter of 10, 50 or 100 nm (nAg10, nAg50 and nAg100, respectively), 10 μg/mL for 24 h. | A549 cells | Global DNA methylation (immunofluorescence staining); DNMT1 and 3B expression (Western blot) | All AgNPs decreased DNA methylation, decreased DNMT1 and increased DNMT3B. nAg10 may induce DNA hypomethylation through a proteasome-mediated degradation of DNMT1. | Maki et al., 2020 [30] |
| AgNPs, 1 µg/mL. | BEAS-2B | Genome-wide methylation (array) | Only 1 differentially methylated promoter of a poorly characterized gene, 6 differentially methylated CpG sites and 5 differentially methylated tiling regions located in intergenic regions with regulatory function. | Gliga et al., 2018 [31] |
| AgNPs capped with glutathione, NA. | HepG2 cells | mRNA and microRNA expression (RNAseq) | DNA methylation was an affected pathway identified in the bioinformatics pathway analyses. | Thai et al., 2021 [32] |
| AgNPs, 5 μg/mL for 48 h, with or without 96 h without treatment. | HT22 cells | Global DNA methylation (ELISA), DNMTs expression (ELISA) | Increase in 5-mC, DNMT1, DNMT2, DNMT3a and DNMT3b levels. The upregulation of DNMT2 may be a part of cellular stress response to AgNPs. | Mytych et al., 2017 [33] |
| AgNPs, 1–20 μg/mL for 24 h. | HEK293T cells | Genome-wide methylation (MeDIP-Seq) | A total of 12 up-regulated and hypomethylated genes and 22 down-regulated and hypermethylated genes, which were primarily involved in lipid and ion metabolism. | Chen et al., 2022 [34] |
| AgNPs, 0.1–100 μM for 3, 6, 12 and 24 h. | EA.hy926 cells | Global DNA methylation (flow cytometry) | Increased global DNA methylation. | González-Palomo et al., 2021 [35] |
| AgNPs, 5–25 μg/mL for 24 h. | NIH3T3 cells | Transcriptomics (RNAseq) | Alterations in epigenetic-related processes including nucleosome assembly and DNA methylation. | Gurunathan et al., 2018 [36] |
| AgNPs (10 µg/mL), AuNPs (10 µg/mL) and superparamagnetic iron oxide nanoparticles (5 µg/mL) for 24 h. | HepG2 cells | Promoter methylation of genes related to inflammatory response and apoptosis | No differences in the methylation for any type of nanoparticle. | Brzóska et al., 2019 [37] |
| AuNPs, 0.8 and 1.6 µg/cm2 for 3, 24 and 48 h. | BEAS-2B and A549 cells | Global DNA methylation (ELISA) | LPS-challenged cells underwent an increase in global DNA 5-mC and a decrease in global 5-hmC, possibly associated with a carcinogenic-like transformation process. | Gambelunghe et al., 2020 [38] |
| AuNPs, 1 nM for 48 or 72 h. | MRC-5 cells | Methylation of the PROS1 gene | No changes in PROS1 methylation. | Ng et al., 2011 [39] |
| AuNPs, 10, 50 and 100 μg/mL for 72 h. | HEK293 and HaCaT cells | Global DNA methylation (Immunocytochemical staining) | No differences in methylation in HEK293 cells and HaCaT cells treated with AuNPs. | Sooklert et al., 2019 [40] |
| AuNPs of 1.5, 4 and 14 nm core sizes, 10 μg/mL for 24 h. | hESCs | Genome-wide methylation (array) | Thiolate-capped 4 nm AuNPs at 10 g/mL caused a dramatic decrease in global DNA methylation and an increase in global DNA hydroxymethylation in only 24 h. | Senut et al., 2016 [41] |
| TiO2, 1–100 μg/mL for 4, 24 and 48 h. | A549 cells | Global DNA methylation (HPLC-MS/MS) and methylation profile of 20 DNA repair gene promoters (qPCR) | No change in the overall DNA methylation. Exposure to 100 µg/mL TiO2 for 4 or 24 h led to a moderate increase in APE1, POLD3, MRE11A and PMS2 methylation. | Biola-Clier et al., 2017 [42] |
| TiO2, 6.25–100 μg/mL for 24 h. | A549 cells | PARP-1 methylation | Increased levels of PARP-1 methylation. | Bai et al., 2015 [43] |
| TiO2 uncoated and coated with silica or citrate, 40 μg/cm2 for 48 and 72 h. | A549 cells | LINE-1 methylation (ELISA) | No effect on LINE-1 methylation after 48 h. After 72 h, a reduction in global DNA methylation levels was induced by all nanoparticles. | Stoccoro et al., 2017 [44] |
| TiO2, 0.1–100 μg/mL−1 for 48 h. | 16HBE and A549 cells | Global DNA methylation (ELISA) and DNMT3B and TET expression (qPCR) | Only the anatase-type TiO2 decreased global DNA methylation and altered expression levels of methylation-related genes and proteins. | Ma et al., 2017 [45] |
| TiO2, 3.25, 12.5 and 25 μg/mL for 3 and 24 h. | 16HBE cells | Global DNA methylation (LC-MS/MS) | Increased global DNA methylation at 24 h for anatase and rutile TiO2, and the mixture of both. | Ghosh et al., 2017 [46] |
| TiO2, 10 and 20 mg/mL for up to 4 weeks. | BEAS-2B cells | Genome-wide methylation | In total, 755 CpG sites were identified with only minor consistent effects of hypomethylation. | Sierra et al., 2017 [47] |
| TiO2, 100 μg/mL for 24 or 72 h. | Caco-2, HepG2, NL20 and A-431 cells | Global DNA methylation (ELISA), gene-specific methylation (array) and expression of DNMTs, MBD2 and UHRF1 | Decrease in global DNA methylation in Caco-2, HepG2 and A-431 cells. Across the four cell lines, eight genes (CDKN1A, DNAJC15, GADD45A, GDF15, INSIG1, SCARA3, TP53 and BNIP3) with promotors methylated after exposure. Altered expression of DNMT1, DNMT3A, DNMT3B, MBD2 and UHRF1, which was cell-type-dependent. | Pogribna et al., 2020 [48] |
| TiO2, 25–100 μg/mL for 24 h. | PBMCs | Global DNA methylation (ELISA) | DNA hypomethylation. | Malakootian et al., 2021 [49] |
| TiO2 and CuONPs, 0.5 and 30 µg/mL for 24 h. | THP-1, RAW264.7 and SAEC cells | Global DNA methylation (LC-MS/MS); LINE1 and Alu/SINE element methylation (methylation-sensitive qPCR) and expression of DNA methylation machinery (qPCR) | Modest alterations in methylation of LINE-1 and Alu/SINE, and decreased expression of DNA methylation machinery in a cell-, dose- and nanomaterial-dependent manner. | Lu et al., 2016 [50] |
| TiO2 and ZnO, 0.125–8 μg/mL for 24, 48 and 72 h. | MRC-5 cells | Global DNA methylation (ELISA), DNMTs activity (ELISA) and expression (qPCR) | Decrease in global DNA methylation and DNA methyltransferase activity. Direct correlation between nanoparticle concentration, global DNA methylation and expression of DNMT1, 3A and 3B. | Patil et al., 2016 [51] |
| ZnO, 25 and 50 μg/mL for 48 h. | HEK-293 cells | Global and locus-specific DNA-methylation at LINE-1, D4Z4 and NBL2, and at the promoter of selected ROS-responsive genes (AOX1, HMOX1, NCF2, SOD3). Global DNA methylation, DNMTs and TET activity | Global reduction in 5-methylcytosine and increase in 5-hydroxymethylcytosine. Significant increase in the expression of TETs, but not in the expression of DNMTs. | Choudhury et al., 2017 [52] |
| CuNPs, NA. | HepG2 cells | mRNA and microRNA expression (RNAseq) | mRNA–microRNA interaction revealed altered DNA methylation. Altered expression of DNMTs. | Thai et al., 2021 [53] |
| CuONPs, 6.25 to 400 mg L−1 for 24 h. | N2A cells | Global DNA methylation (HPLC) | Changes in the m5dC/dC ratio were less than 1%, which may indicate that CuONPs do not alter DNA methylation in vitro. | Perreault et al., 2012 [54] |
| Maghemite nanoparticles covered with citric acid at 0.5 and 3.0 mg FemL−1 for 24 or 48 h. | HSG cells | Global DNA methylation (ELISA) | Altered global DNA methylation with reduced expression of genes related to epigenetic reprogramming. | Bonadio et al., 2020 [55] |
| Pristine plasma and laser ITER-like tungsten nanoparticles, 1–5 µg/mL for 24 h. | BEAS-2B cells | DNA methylation of Alu, LINE and Satellite 2 and 3 (Sat 2 and Sat 3) repeats (bisulfite sequencing) | No significant changes in DNA methylation. | Uboldi et al., 2019 [56] |
| Carbon-based nanomaterials | ||||
| SWCNTs, 10 µg/mL with 30- and 60-day recovery periods. | BEAS-2B cells | Genome-wide methylation (array); gene-specific methylation (methylation-specific PCR) | DNMT3A and DNMT1 up-regulation after 30–60 days of recovery. A total of 457 hypermethylated and 367 hypomethylated gene promoters. Hypermethylation of PIM2 gene and hypomethylation of ABCA2 and CRYBG3 genes in the 60-day recovery period group. | Wang et al., 2021 [57] |
| MWCNTs or SWCNTs, 0.25 µg/mL for four weeks and recovery period of two weeks. | 16HBE cells | Genome-wide methylation (array) | MWCNTs induced a single hypomethylation at a CpG site on a gene promoter. Exposure to SWCNTs induced hypermethylation at CpG sites which may involve ‘transcription factor activity’ and ‘sequence-specific DNA binding’ gene ontologies. After the recovery period, no change in DNA methylation for MWCNTs, and hypermethylation and hypomethylation for SWCNTs. HPCAL1, PRSS3, KLK3, KLF3 genes were hypermethylated at different time points in SWCNT-exposed cells. | Öner et al., 2020 [58] |
| MWCNTs and SWCNTs, 25 and 100 μg/mL for 24 h. | 16HBE cells | Global DNA methylation and hydroxymethylation (LC-MS/MS); whole-genome methylation (array) | MWCNTs hypomethylated 2398 gene promoters; after exposure to SWCNTs, 589 CpG sites (located on 501 genes) were either hypo- (N = 493) or hypermethylated (N = 96). Differentially methylated and expressed genes induced changes (MWCNTs > SWCNTs) at different cellular pathways, such as p53 signaling, DNA damage repair and cell cycle. SWCNT exposure showed hypermethylation on SKI, GTSP1, SHROOM2 and NF1 genes. | Öner et al., 2018 [59] |
| MWCNTs and SWCNTs, 25 and 100 μg/mL for 3 and 24 h. | THP-1 cells | Global DNA methylation (LC-MS/MS), genome-wide CpG site-specific methylation (array) | No global DNA 5-mC or 5-hmC changes. MWCNTs hypomethylated 3340 promoter regions (2398 genes), with no differential methylation at individual CpG sites. SWCNTs hypomethylated 5 gene promoters (AKAP8L, FOXK2, EIF4E, snoU13 and RP11-223 l10.1); 493 hypomethylated and 96 hypermethylated single CpG sites, located on 501 different genes. | Öner et al., 2017 [60] |
| MWCNTs, 5, 10 and 15 μg/mL for 24, 48 and 72 h. | THP-1 cells | Genome-wide methylation (array) | Increasing dose-dependent trend of differentially methylated promoters at 24 h and a dose-dependent decrease in hypomethylated promoters at 48 h. | Saarimäki et al., 2020 [61] |
| MWCNTs, 10 and 20 mg/mL for up to 4 weeks. | BEAS-2B cells | Genome-wide methylation | 755 CpG sites were identified with only minor consistent effects of hypomethylation. | Sierra et al., 2017 [47] |
| Functionalized MWCNTs (hydroxylation (8.37 and 6.34 mg/L); carboxylation (37.99 and 4.44 mg/L) and pristine (2.92 and 2.17 mg/L)) | BEAS-2B and HepG2 cells | Global DNA methylation (ELISA) | DNMT3B-dependent hypo-methylation in BEAS-2B cells and hyper-methylation in HepG2 cells in a functionalization-dependent manner. | Chatterjee et al., 2017 [62] |
| Carbon black, fullerene, graphite nanofibers, SWCNTs and MWCNTs, 0.1–500 μg/mL for 48 h. | A549, BEAS-2B and THP-1 cells | Genome-wide methylation (array) | Molecular alterations are highly dependent on the cell type and geometrical properties of the carbon nanomaterials. | Scala et al., 2018 [63] |
| Fullerene, long or short MWCNTs or SWCNTs, 0.1 mg/L. | A549 cells | Global DNA methylation (HPLC-MS), DNMTs expression (qPCR) | Increased global DNA methylation. Down-regulating tendency in DNMT transcription, except for C60, but only significant for DNMT3b after SWCNT treatment. | Li et al., 2016 [64] |
| GONPs, 1 and 10 µg/mL for 15 and 30 days. | BEAS-2B cells | LINE-1, D4Z4 and NBL2, SATα and AluYb8 methylation (bisulfite pyrosequencing); genome-wide DNA methylation (array) | No genome-wide or global DNA methylation changes associated with either condition. | Pérez et al., 2020 [65] |
| Pristine, carboxylated and aminated graphene, graphene nanoplatelets, SLGO and FLGO, 10 and 50 mg/L for 24 h. | BEAS-2B cells | Global DNA methylation (ELISA) | Increased global DNA methylation after exposure to SLGO/FLGO and decreased global DNA methylation after exposure to the remaining nanoparticles. | Chatterjee et al., 2016 [66] |
| GQD, 50 μg/mL. | mESCs | DNMT1, DNMT3A, DNMT3B, TET1, TET2 and TET3 gene expression (qPCR); methylation of Sox2 and Oct4 promoter regions (bisulfite treatment and NGS) | GQD-induced inhibition in CpG methylation of Sox2 through altering methyltransferase and demethyltransferase expression. | Ku et al., 2021 [67] |
| Silica nanoparticles | ||||
| SiNPs, 20 μg/mL for 24 h. | GC-2 cells | MeDIP-seq | Extensive methylation changes, with a total of 428 hyper-methylated genes and 398 hypo-methylated genes, probably involved with abnormal transcription and translation, mitochondrial damage and cell apoptosis. | Sang et al., 2021 [68] |
| SiNPs, 3.125–100 μg/mL for 24 h. | BEAS-2B cells | Genome-wide methylation (array) | Of the 25 significant altered processes, the apoptosis-related PI3K/Akt pathway involved 32 differentially methylated gene promoters, in which CREB3L1 and Bcl-2 were hypermethylated, in association with the downregulation of their mRNA levels. | Zou et al., 2016 [69] |
| SiNPs, 10, 50 and 100 μg/mL for 72 h. | HEK293 and HaCaT cells | Global DNA methylation (Immunocytochemical staining) | No differences in methylation in HEK293 cells, but HaCaT cells exposed to 10 µg/mL SiNPs had lower levels of methylation. | Sooklert et al., 2019 [40] |
| SiO2NPs, 2.5, 5 and 10 μg/mL for 24 h. | HaCaT cells | Promoter methylation of PARP-1 (methylation-specific PCR and bisulfite sequencing) | Decrease in PARP-1 mRNA and protein levels and a simultaneous increase in PARP-1 methylation. | Gong et al., 2012 [70] |
| SiNPs, 2–10 µg/mL for 48 h. | HaCaT cells | Global DNA methylation (flow cytometry); DNMTs expression (qPCR and Western blot) | Decreased levels of DNMT1, DNMT3A and MBD2 in a dose-dependent manner at mRNA and protein levels. Global DNA methylation decreased with dose. | Gong et al., 2010 [71] |
| SiNPs, 2 and 5 μg/cm2 for 6 days. | Bhas 42 cells | Global DNA methylation (ELISA) and DNMTs expression (Western blot) | SiNPs treatment did not affect DNMT3A and DNMT3B expression or DNA methylation. | Seidel et al., 2017 [72] |
| Other nanomaterials | ||||
| CSNPs, 10, 50 and 100 μg/mL for 72 h. | HEK293 and HaCaT cells | Global DNA methylation (Immunocytochemical staining) | No differences in methylation in HEK293 cells, but HaCaT cells exposed to 100 µg/mL CSNPs had lower levels of methylation. | Sooklert et al., 2019 [40] |
| Cromolyn CSNPs and CSNPs, 62.5, 125, 250, 500 μg/mL for 48 h. | MCF-7 cells | Global DNA methylation (DNMT1 and METTL3 expression (qPCR); methylation of RASSF1A and p16 genes (methylation-specific PCR) | Reduction in DNMT1 expression, reversed hypermethylation pattern of RASSF1A and p16 genes and lower expression of METTL3. Cromolyn chitosan nanoparticles may act by inhibiting ERK1/2 phosphorylation/DNMT1/DNA methylation, possibly impacting RNA methylation machinery via METTL3 expression. | Motawi et al., 2022 [73] |
| ChiNH/Q, 10–1000 μg/mL for 48 h. | HepG2 cells | Global DNA methylation (ELISA) and expression of DNMTs (qPCR) | Reduced expression levels of DNMT1/3A/3B and increased levels of 5-mC. | Abbaszadeh et al., 2020 [74] |
| Dendrosomal nano curcumin, 0–45 μM for 24 h and 0–38 μM for 48 h. | HepG2 and Huh7 cells | DNMT1, DNMT3A and 3B expression (semi-quantitative and qPCR) | Downregulation of DNMT1, DNMT3A and DNMT3B expression in both cell lines. | Chamani et al., 2016 [75] |
| PEPs, 0.5–100 μg/mL for 8 h or more. | SAECs, THP-1 and TK6 cells | Methylation and expression of transposable elements (TEs) (qPCR), LINE-1 copy number analysis (qPCR), expression of DNMT1, DNMT3A, DNMT3b, UHRF1 and TET1, TET2, TET3 (qPCR) | Dysfunction of the DNA methylation and demethylation machinery associated with the loss of DNA methylation and the reactivation of TEs. | Pirela et al., 2016 [76] |
| PEPs and mild steel welding fumes, 0.5 and 30 µg/mL for 24 h. | THP-1, RAW264.7 and SAEC cells | Global DNA methylation (LC-MS/MS); LINE1 and Alu/SINE element methylation (methylation-sensitive qPCR) and expression of DNA methylation machinery (qPCR) | Modest alterations in methylation of LINE-1 and Alu/SINE, and decreased expression of DNA methylation machinery in a cell-, dose- and nanomaterial-dependent manner. | Lu et al., 2016 [50] |
| GO-AgNPs, 4 and 8 μg/mL for 24 h. | CFFCs cells | Global DNA methylation, DNMTs expression (qPCR) | DNA hypomethylation and decreased expression of DNMT3A. | Yuan et al., 2021 [77] |
| Nanomaterials, Exposure Conditions | Animal Model | Methodology | Main Conclusions | References |
|---|---|---|---|---|
| Metal-based nanomaterials | ||||
| AgNPs, 0.5, 2.5 and 12.5 mg/kg BW for 7 days. | C57BL/6J mice | Global DNA methylation (ELISA) | Decreased global DNA methylation and DNA hydroxymethylation in the livers of mice with high-fat-diet-induced non-alcoholic fatty liver disease (NAFLD), contributing to NAFLD development and progression. | Wen et al., 2022 [78] |
| AgNPs, 1.0 mg/kg for 17.5 days. | ICR mice | Methylation of Zac1 and Igf2r genes (bisulfite sequencing) | AgNP exposure significantly altered the methylation levels of Zac1. | Zhang et al., 2015 [79] |
| AgNPs, 0.4 mg/L. | Zebrafish embryos | Gene-specific methylation | Myogenic loci-specific DNA methylation might result in muscle dysfunctions in treated embryos. | Xu et al., 2018 [80] |
| AuNPs of 5, 60 and 250 nm, 2.5 mg/kg and 0.25 mg/kg for 48 h. | BALB/c mice | Global DNA methylation and hydroxymethylation (LC-MS), gene-specific methylation of 17 genes (bisulfite pyrosequencing) | AuNP exposure had no effect on 5-mC and 5-hmC levels in mouse lungs. AuNP 60 nm induced CpG hypermethylation in Atm, Cdk and Gsr genes and hypomethylation in Gpx; Gsr and Trp53 showed changes in methylation between low- and high-dose AuNP, 60 and 250 nm, respectively, and AuNP had size effects on methylation for Trp53. | Tabish et al., 2017 [81] |
| TiO2, 12 mg/mL for 6 h/day for 6 non-consecutive days. | FVB/NJ pregnant dams and fetal pups | Global DNA methylation (ELISA), Hif1α activity (ELISA), DNMT activity (colorimetric assay) | DNA methylation was significantly increased in fetal pups following maternal exposure, along with increased Hif1α activity and DNMT1 protein expression. | Kunovac et al., 2019 [82] |
| TiO2 of 25 nm, 80 mg/mL for 30 days. | NIH mice | Global DNA methylation (ELISA), promoter methylation of IFN-gama, TNF-alfa, Thy-1 | Decreased global DNA methylation and hydroxymethylation in the lung tissue only in the young group. Altered methylation of TNF-alfa and Thy-1 promoters with a role in inflammation and fibrosis. | Ma et al., 2019 [83] |
| CuNPs, 6.5 and 3.25 mg/kg for 4 weeks. | Wistar rats | Global DNA methylation (ELISA) | Lowering the level of copper nanoparticles in the diet increased DNA methylation. | Ognik et al., 2019 [84] |
| CuONPs, 3.3 mg m−3 and 13.2 mg m−3 for 6 h. | Wistar Unilever outbred rats | Methylation in inflammation-related genes (PCR array coupled with DNA restriction kit) | No aberrant DNA methylation of inflammation-associated genes. | Costa et al., 2018 [85] |
| CuONPs, 2.5 mg/kg body weight. | BALB/c mice | Global DNA methylation (LC-MS/MS), methylation at the LINE-1 and SINE B1 elements (methylation-sensitive qPCR), DNMT1/3A/3B expression (qPCR) | CuONPs increased the 5-mC and 5-hmC levels in lung tissue. CuONPs reduced the expressions of DNMT1, 3a and 3b in the lung tissue, but not in alveolar macrophages. The expression of TET1 decreased in both alveolar macrophages and lung tissue after exposure to CuO. | Lu et al., 2016 [86] |
| CuONPs, 0–500–1400 mg Cu/kg soil (DW). | Enchytraeus crypticus (soil invertebrate) | Global DNA methylation (immunostaining) | Differences in 5-mC between E. crypticus generations after exposure. | Bicho et al., 2021 [87] |
| CuONPs, 500–1400 mg Cu/kg soil for 32 days. | Enchytraeus crypticus | Global DNA methylation (LC-MS), gene-specific DNA methylation (qPCR and MS-HRM) and bisulfite sequencing | Multigenerational long-term exposure to CuO NMs induced changes in epigenetic markers. However, global DNA methylation and gene-specific methylation did not confirm the epigenetic effect. | Bicho et al., 2020 [88] |
| Nickel oxide nanoparticles, NA. | Mice with pulmonary fibrosis | Genome-wide methylation (whole-genome bisulfite sequencing) and transcriptomics | Hypomethylation in lung fibrotic tissue. mRNA transcriptome data found 93 DNA methylation genes with transcriptional significance. | Zheng et al., 2022 [89] |
| Metal-rich welding nanoparticles, 2.0 mg/rat for 30 days. | Sprague-Dawley rats | Global DNA methylation (ELISA) | No significant differences were observed when comparing DNA methylation between the welding fume and control groups at any of the time points. | Shoeb et al., 2017 [90] |
| Carbon-based nanoparticles | ||||
| SWCNTs and MWCNTs, 2.5 mg/kg and 0.25 mg/kg for 48 h. | BALB/c mice | Global DNA methylation and hydroxymethylation (LC-MS), gene-specific methylation of 17 genes (bisulfite pyrosequencing) | SWCNT and MWCNT exposure had no effect on 5-mC and 5-hmC levels in mouse lungs. SWCNT exposure induced promoter hypomethylation in Atm. | Tabish et al., 2017 [81] |
| SWCNTs, MWCNTs and fullerene, 0.1 mg/L for 21 days. | Zebrafish | Global DNA methylation (LC-MS) | Increased global genomic methylation, most profound in female zebrafish brain tissues, after exposure to short MWCNTs or SWCNTs. | Gorrochategui et al., 2017 [91] |
| GQD, 2, 10 and 50 mg/L for 7 days. | Zebrafish | Global DNA methylation (LC-MS/MS) | Global DNA hypermethylation in various tissues in a dose-dependent manner. The global DNA methylation of reduced and aminated GQD exposure increased in intestines even at low concentrations. Fourteen days after exposure, the effects had ceased. DNA methylation in the livers of fish from exposure groups was higher, even after exposure had ceased, indicating a more complex mechanism of DNA methylation deregulation. | Hu et al., 2019 [92] |
| GONPs with cis-bifenthrin (cis-BF) (0.06 and 0.3 μg/L) or cis-BF alone (0.1 mg/L) for 21 days. | Xenopus laevis (tadpoles) | Global DNA methylation (ELISA) | Reduced levels of genomic DNA methylation were observed in the co-exposure groups. | Li et al., 2020 [93] |
| Black carbon, 10 and 30 μg/mL for 60 days. | Zebrafish | Global DNA methylation (ELISA), promoter methylation (touch-down PCR of bisulfite-treated DNA) | Increased global genome methylation in a dose-dependent manner, upregulation of the mRNA content of DNMT3B and TET2 in heart tissue and dose-dependent decreases in the mRNA expression of DNMT1, DNMT3A and TET1. Increased unmethylated CpG dinucleotide sites at lepb, cd248b and il11a promoters. | Zhou et al., 2019 [94] |
| Other nanoparticles | ||||
| Food-grade precipitated silica (S200) and fumed silica Aerosil 200F (A200F), 225, 1000 and 5000 mg/kg for 28 and 84 days. | BALB/c mice | Global DNA methylation (ELISA), methylation levels of LINE-1 and SINEB1 (pyrosequencing), genome-wide methylation (NGS) | Changes in whole-genome methylation in peripheral mouse leukocytes and liver after 28 days. After 84 days of high-dose continuous exposure, differential methylation was mainly found in introns, intergenic regions and promoters. | Lu et al., 2021 [95] |
| PEPs, 2.5 mg/kg body weight. | BALB/c mice | Global DNA methylation (LC-MS/MS), methylation at the LINE-1 and SINE B1 elements (methylation-sensitive qPCR), DNMT1/3A/3B expression (qPCR) | PEPs increased the 5-mC and 5-hmC levels in lung tissue. Hypermethylation of the LINE-1 element was observed in mouse lung tissue after exposure to PEPs. PEPs increased the expression of DNMT1 in the alveolar macrophages and down-regulated DNMT3a expression in the alveolar macrophages and lung tissue; the expression of TET1 decreased in both alveolar macrophages and lung tissue after exposure. | Lu et al., 2016 [86] |
| CSNPs, 5 mg cromolyn/kg twice a week for 2 weeks. | Swiss albino mice injected with Ehrlich ascites carcinoma cells subcutaneously | DNMT1 and METTL3 expression (qPCR) | Cromolyn CSNPs lessened the tumor volume and halted DNMT1 and METTL3 expression in Ehrlich carcinoma mice. | Motawi et al., 2022 [73] |
| Nanomaterial | Population | Methodology | Main Conclusions | References |
|---|---|---|---|---|
| PM < 25–100 nm | 14 nanocomposite research workers (10 exposed and 4 controls) from 2016 to 2019. | Genome-wide methylation (microarray) | Shift in individual DNA methylation patterns in the blood of all the exposed and control subjects, between 2016 and 2019. Differences seem to be consistently greater in the NP-exposed subjects compared with the controls. The selected 14 most differently methylated CG loci were relatively stable in the chronically exposed subjects. | Rossnerova et al., 2021 [96] |
| TiO2, SiNPs and indium tin oxide (ITO) | 172 workers from 14 nanomaterial manufacturing and/or handling factories (130 exposed and 43 controls). | Global DNA methylation (HPLC) | Global DNA methylation in white blood cells was decreased in ITO-exposed workers compared with controls. | Liou et al., 2017 [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valente, A.; Vieira, L.; Silva, M.J.; Ventura, C. The Effect of Nanomaterials on DNA Methylation: A Review. Nanomaterials 2023, 13, 1880. https://doi.org/10.3390/nano13121880
Valente A, Vieira L, Silva MJ, Ventura C. The Effect of Nanomaterials on DNA Methylation: A Review. Nanomaterials. 2023; 13(12):1880. https://doi.org/10.3390/nano13121880
Chicago/Turabian StyleValente, Ana, Luís Vieira, Maria João Silva, and Célia Ventura. 2023. "The Effect of Nanomaterials on DNA Methylation: A Review" Nanomaterials 13, no. 12: 1880. https://doi.org/10.3390/nano13121880