
Cells on Hydrogels with Micron-Scaled Stiffness Patterns Demonstrate Local Stiffness Sensing

 ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fabrication of Stiffness-Patterned Polyacrylamide Hydrogel
2.2. Mechanical and Geometrical Characterisation of the Stiffness Patterns
2.3. Functionalization of the Stiffness-Patterned Hydrogel
2.4. Characterization of the Surface Chemistry
2.5. Cell Culture and Immunostaining
2.6. Calculations of Cellular Stresses
2.6.1. Deformation Field
2.6.2. Traction Force Calculation
2.7. TFM Analysis
3. Statistical Analysis
4. Results
4.1. Micron-Scaled Rigidity Patterns with kPa/µm Gradients
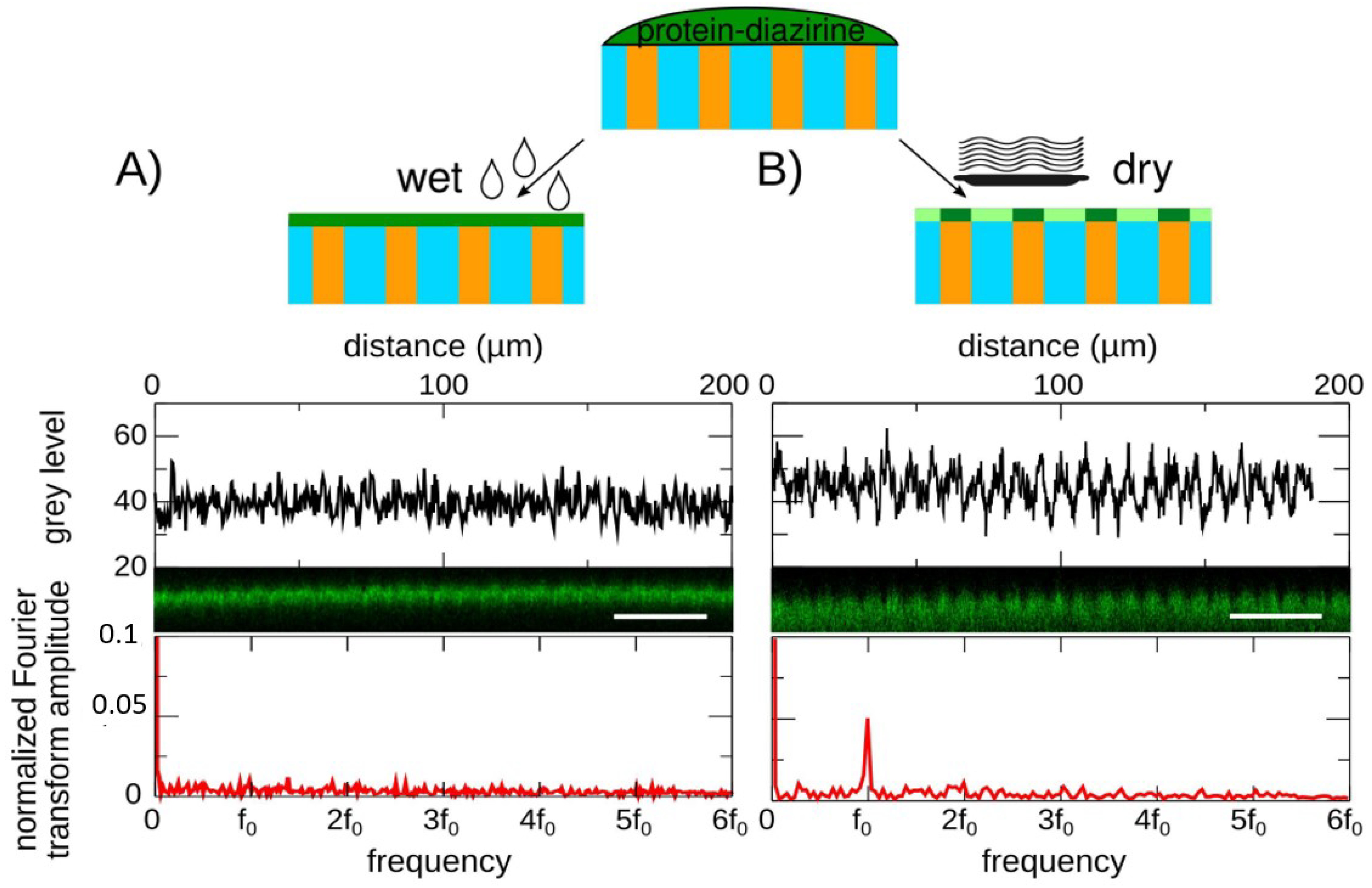
4.2. Tuning of the Uniformity of the Surface Coating by the Hydration Rate of the Hydrogel
4.3. Localization of the Adhesion Proteins on the Stiff Patterns on Uniform Surface Coating
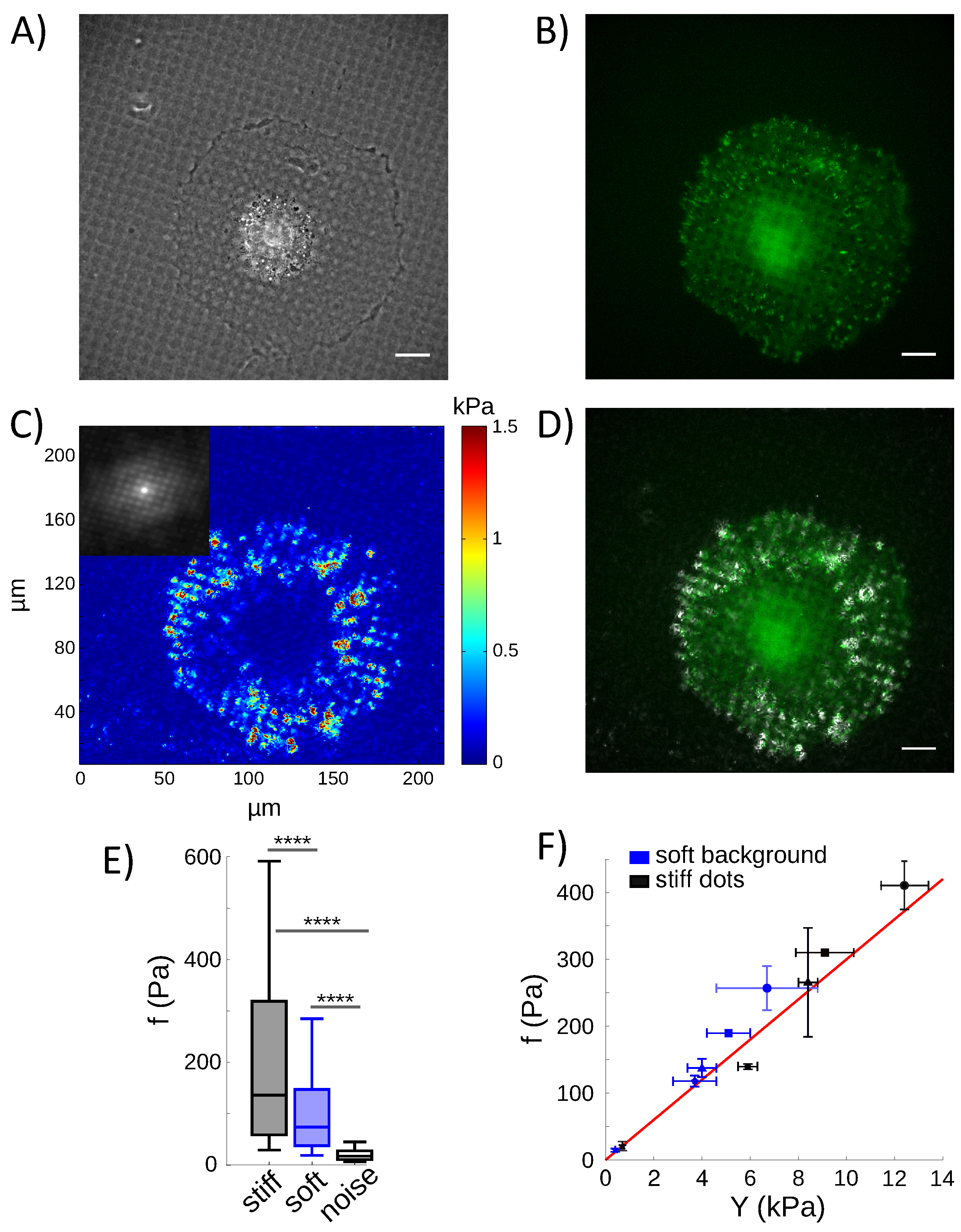
4.4. Calculation of Cell Traction Stresses on Rigidity-Patterned Substrates
4.5. Cells Pull on the Rigidity Patterns Proportional to the Local Stiffness
5. Discussion
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TFM | traction force microscopy |
| KLT | Kanade–Lucas–Tomasi |
| PDMS | poly (dimethyl siloxane) |
Appendix A. Discrimination of the Stiff and Soft Regions for the Analysis of the Cellular Stresses

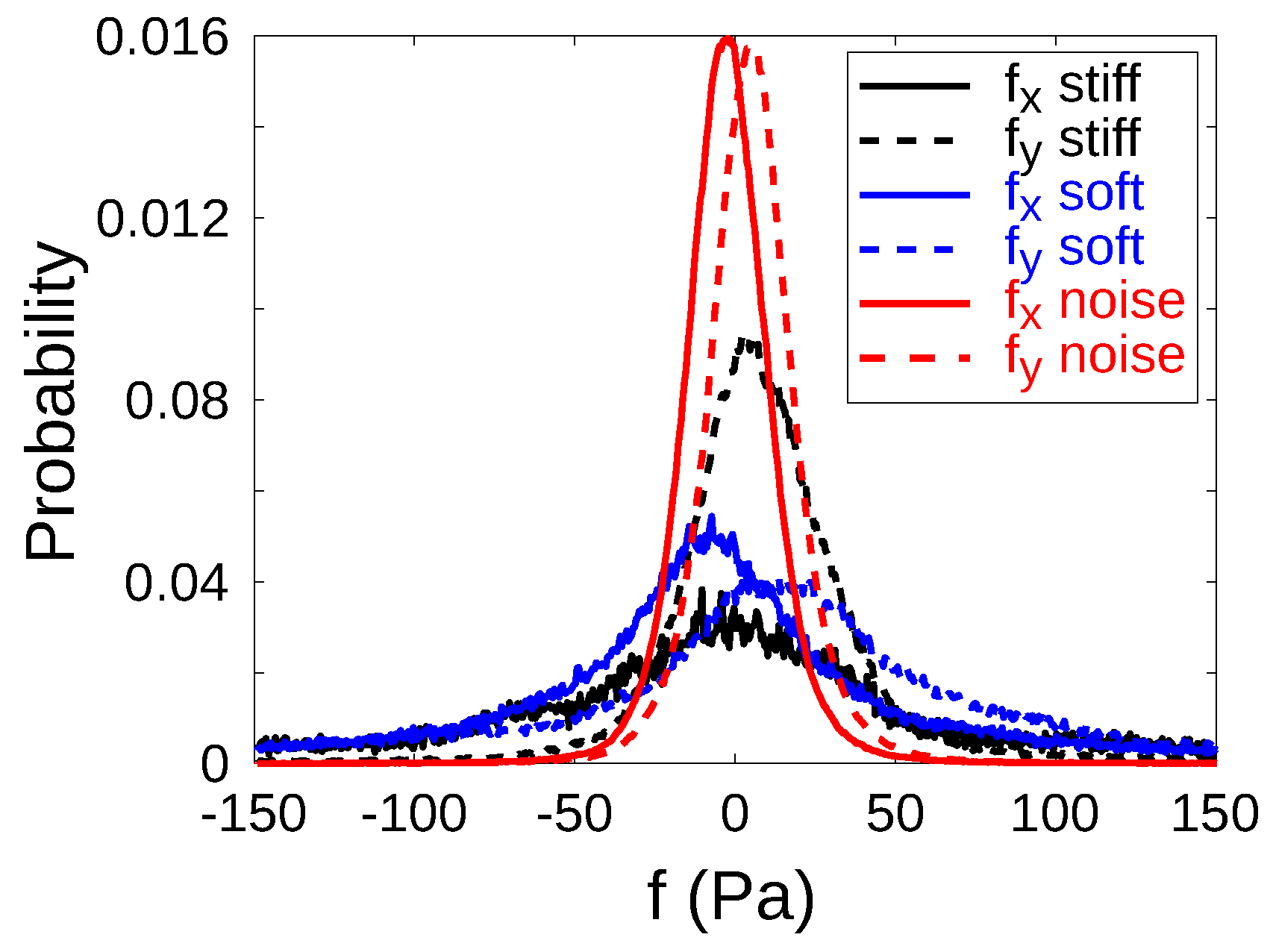
Appendix B. Filtering Cellular Stresses: Gaussian Noise

References
- Macri-Pellizzeri, L.; De-Juan-Pardo, E.M.; Prosper, F.; Pelacho, B. Role of substrate biomechanics in controlling (stem) cell fate: Implications in regenerative medicine. J. Tissue Eng. Regener. Med. 2018, 12, 1012–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perestrelo, T.; Correia, M.; Ramalho-Santos, J.; Wirtz, D. Metabolic and Mechanical Cues Regulating Pluripotent Stem Cell Fate. Trends Cell Biol. 2018, 28, 1014–1029. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.E. From mechanobiology to developmentally inspired engineering. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2018, 373. [Google Scholar] [CrossRef] [Green Version]
- Xi, W.; Saw, T.B.; Delacour, D.; Lim, C.T.; Ladoux, B. Material approaches to active tissue mechanics. Nat. Rev. Mat. 2018, 4, 23–44. [Google Scholar] [CrossRef]
- Smith, L.R.; Cho, S.; Discher, D.E. Stem Cell Differentiation is Regulated by Extracellular Matrix Mechanics. Physiology 2018, 33, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Kandow, C.E.; Georges, P.C.; Janmey, P.A.; Beningo, K.A. Polyacrylamide Hydrogels for Cell Mechanics: Steps Toward Optimization and Alternative Uses. In Cell Mechanics; Wang, Y.L., Discher, D.E., Eds.; Academic Press: New York, NY, USA, 2007; Volume 83, pp. 29–46. [Google Scholar]
- Levental, I.; Georges, P.C.; Janmey, P.A. Soft biological materials and their impact on cell function. Soft Matter 2007, 3, 299–306. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Paszek, M.J.; Zahir, N.; Johnson, K.R.; Lakins, J.N.; Rozenberg, G.I.; Gefen, A.; Reinhart-King, C.A.; Margulies, S.S.; Dembo, M.; Boettiger, D.; et al. Tensional homeostasis and the malignant phenotype. Cancer Cell 2005, 8, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Verhulsel, M.; Vignes, M.; Descroix, S.; Malaquin, L.; Vignjevic, D.M.; Viovy, J.L. A review of microfabrication and hydrogel engineering for micro-organs on chips. Biomaterials 2014, 35, 1816–1832. [Google Scholar] [CrossRef]
- Correia Carreira, S.; Begum, R.; Perriman, A.W. 3D bioprinting: The emergence of programmable biodesign. Adv. Healthc. Mater. 2020, 9, 1900554. [Google Scholar] [CrossRef]
- Bertassoni, L.E. Bioprinting of Complex Multicellular Organs with Advanced Functionality—Recent Progress and Challenges Ahead. Adv. Mater. 2022, 34, e2101321. [Google Scholar] [CrossRef]
- Doyle, A.D.; Carvajal, N.; Jin, A.; Matsumoto, K.; Yamada, K.M. Local 3D matrix microenvironment regulates cell migration through spatiotemporal dynamics of contractility-dependent adhesions. Nat. Commun. 2015, 6, 8720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuFort, C.C.; Paszek, M.J.; Weaver, V.M. Balancing forces: Architectural control of mechanotransduction. Nat. Rev. Mol. Cell Biol. 2011, 12, 308–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.W.; Mooney, D.J. Improving Stem Cell Therapeutics with Mechanobiology. Cell Stem Cell 2016, 18, 16–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.; Bouchonville, N.; Gaude, C.; Gay, E.; Ratel, D.; Nicolas, A. The Micro-Mechanical Signature of Pituitary Adenomas: New Perspectives for the Diagnosis and Surgery. Adv. NanoBiomed. Res. 2021, 1, 2000085. [Google Scholar] [CrossRef]
- Bershadsky, A.D.; Balaban, N.Q.; Geiger, B. Adhesion-dependent cell mechanosensitivity. Annu. Rev. Cell Dev. Biol. 2003, 19, 677–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plodinec, M.; Loparic, M.; Monnier, C.A.; Obermann, E.C.; Zanetti-Dallenbach, R.; Oertle, P.; Hyotyla, J.T.; Aebi, U.; Bentires-Alj, M.; Lim, R.Y.H.; et al. The nanomechanical signature of breast cancer. Nat. Nanotechnol. 2012, 7, 757–765. [Google Scholar] [CrossRef]
- Bouchonville, N.; Meyer, M.; Gaude, C.; Gay, E.; Ratel, D.; Nicolas, A. AFM mapping of the elastic properties of brain tissue reveals kPa/µm gradients of rigidity. Soft Matter 2016, 12, 6232–6239. [Google Scholar] [CrossRef]
- Wong, S.; Guo, W.H.; Hoffecker, I.; Wang, Y.L. Preparation of a micropatterned rigid-soft composite substrate for probing cellular rigidity sensing. Meth. Cell Biol. 2014, 121, 3–15. [Google Scholar] [CrossRef]
- Gray, D.S.; Tien, J.; Chen, C.S. Repositioning of cells by mechanotaxis on surfaces with micropatterned Young’s modulus. J. Biomed. Mat. Res. Part A 2003, 66A, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Lieu, Z.Z.; Wolfenson, H.; Hameed, F.M.; Bershadsky, A.D.; Sheetz, M.P. Mechanosensing Controlled Directly by Tyrosine Kinases. Nano Lett. 2016, 16, 5951–5961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biggs, M.J.P.; Fernandez, M.; Thomas, D.; Cooper, R.; Palma, M.; Liao, J.; Fazio, T.; Dahlberg, C.; Wheadon, H.; Pallipurath, A.; et al. The Functional Response of Mesenchymal Stem Cells to Electron-Beam Patterned Elastomeric Surfaces Presenting Micrometer to Nanoscale Heterogeneous Rigidity. Adv. Mater. 2017, 29, 1702119. [Google Scholar] [CrossRef] [PubMed]
- Jagiello, A.; Hu, Q.; Castillo, U.; Botvinick, E. Patterned photocrosslinking to establish stiffness anisotropies in fibrous 3D hydrogels. Acta Biomater. 2021, in press. [Google Scholar] [CrossRef]
- Kechagia, J.Z.; Ivaska, J.; Roca-Cusachs, P. Integrins as biomechanical sensors of the microenvironment. Nat. Rev. Mol. Cell Biol. 2019, 20, 457–473. [Google Scholar] [CrossRef]
- Discher, D.E.; Janmey, P.; Wang, Y.L. Tissue cells feel and respond to the stiffness of their substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.E.; Odde, D.J. Traction dynamics of filopodia on compliant substrates. Science 2008, 322, 1687–1691. [Google Scholar] [CrossRef]
- Mitrossilis, D.; Fouchard, J.; Guiroy, A.; Desprat, N.; Rodriguez, N.; Fabry, B.; Asnacios, A. Single-cell response to stiffness exhibits muscle-like behavior. Proc. Natl. Acad. Sci. USA 2009, 106, 18243–18248. [Google Scholar] [CrossRef] [Green Version]
- Weng, S.; Shao, Y.; Chen, W.; Fu, J. Mechanosensitive subcellular rheostasis drives emergent single-cell mechanical homeostasis. Nat. Mater. 2016, 15, 961. [Google Scholar] [CrossRef] [Green Version]
- Northcott, J.M.; Dean, I.S.; Mouw, J.K.; Weaver, V.M. Feeling Stress: The Mechanics of Cancer Progression and Aggression. Front. Cell Dev. Biol. 2018, 6, 17. [Google Scholar] [CrossRef]
- Doss, B.L.; Pan, M.; Gupta, M.; Grenci, G.; Mège, R.M.; Lim, C.T.; Sheetz, M.P.; Voituriez, R.; Ladoux, B. Cell response to substrate rigidity is regulated by active and passive cytoskeletal stress. Proc. Natl. Acad. Sci. USA 2020, 117, 12817–12825. [Google Scholar] [CrossRef] [PubMed]
- Palva, S.; Joanne, P.; Migdal, C.; Lopez Soler, E.; Hovhannisyan, Y.; Nicolas, A.; Agbulut, O. Polyacrylamide hydrogels with rigidity-independent surface chemistry show limited long-term maintenance of pluripotency of human induced pluripotent stem cells on soft substrates. ACS Biomater. Sci. Eng. 2020, 6, 340–351. [Google Scholar] [CrossRef]
- Pargon, E.; Menguelti, K.; Martin, M.; Bazin, A.; Chaix-Pluchery, O.; Sourd, C.; Derrough, S.; Lill, T.; Joubert, O. Mechanisms involved in HBr and Ar cure plasma treatments applied to 193 nm photoresists. J. Appl. Phys. 2005, 105, 094902. [Google Scholar] [CrossRef]
- Zelsmann, M.; Truffier-Boutry, D.; Francone, A.; Alleaume, C.; Kurt, I.; Beaurain, A.; Pelissier, B.; Pepin-Donat, B.; Lombard, C.; Boussey, J. Double-anchoring fluorinated molecules for antiadhesion mold treatment in UV nanoimprint lithography. J. Vac. Sci. Technol. B 2009, 27, 2873–2876. [Google Scholar] [CrossRef]
- Frey, M.T.; Engler, A.; Discher, D.E.; Lee, J.; Wang, Y.L. Microscopic Methods for Measuring the Elasticity of Gel Substrates for Cell Culture: Microspheres, Microindenters, and Atomic Force Microscopy. In Cell Mechanics; Wang, Y., Discher, D.E., Eds.; Academic Press: New York, NY, USA, 2007; Volume 83, pp. 47–65. [Google Scholar]
- Barbacci, A.; Diener, J.; Heémon, P.; Adam, B.; Donès, N.; Reveret, L.; Moulia, B. A robust videogrametric method for the velocimetry of wind-induced motion in trees. Agric. For. Meteorol. 2014, 184, 220–229. [Google Scholar] [CrossRef]
- Diener, J. CRToolbox. Available online: https://sites.google.com/site/crtoolbox/home (accessed on 1 January 2022).
- Lucas, B.D.; Kanade, T. An Iterative Image Registration Technique with an Application to Stereo Vision. In Proceedings of the 7th International Joint Conference on Artificial Intelligence, Vancouver, BC, Canada, 24–28 August 1981; pp. 674–679. [Google Scholar]
- Holenstein, C.N.; Silvan, U.; Snedeker, J.G. High-resolution traction force microscopy on small focal adhesions-improved accuracy through optimal marker distribution and optical flow tracking. Sci. Rep. 2017, 7, 41633. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.P.; Tolic-Nørrelykke, I.M.; Fabry, B.; Fredberg, J.J. Traction fields, moments, and strain energy that cells exert on their surroundings. Am. J. Physiol. Cell Physiol. 2002, 282, C595–C605. [Google Scholar] [CrossRef] [Green Version]
- Sabass, B.; Gardel, M.L.; Waterman, C.M.; Schwarz, U.S. High resolution traction force microscopy based on experimental and computational advances. Biophys. J. 2008, 94, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Landau, L.; Lifchitz, E. Theory of Elasticity; Springer: Berlin/Heidelberg, Germany, 1967. [Google Scholar]
- Decker, C. Polymérisation Sous Rayonnement. Techniques de L’ingénieur. Traité Plastiques et Composites. 2000. Available online: https://lhc-div-mms.web.cern.ch/tests/MAG/docum/Radiation_resistance/Literature/am3043-polymerisation-sous-rayonnement.pdf (accessed on 7 May 2012).
- Reinhart-King, C.; Dembo, M.; Hammer, D. Endothelial Cell Traction Forces on RGD-Derivatized Polyacrylamide Substrata. Langmuir 2003, 19, 1573–1579. [Google Scholar] [CrossRef]
- Yang, C.; DelRio, F.W.; Ma, H.; Killaars, A.R.; Basta, L.P.; Kyburz, K.A.; Anseth, K.S. Spatially patterned matrix elasticity directs stem cell fate. Proc. Natl. Acad. Sci. USA 2016, 113, E4439–E4445. [Google Scholar] [CrossRef] [Green Version]
- Zaidel-Bar, R.; Itzkovitz, S.; Ma’ayan, A.; Iyengar, R.; Geiger, B. Functional atlas of the integrin adhesome. Nat. Cell Biol. 2007, 9, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Ghassemi, S.; Meacci, G.; Liu, S.; Gondarenko, A.A.; Mathur, A.; Roca-Cusachs, P.; Sheetz, M.P.; Hone, J. Cells test substrate rigidity by local contractions on submicrometer pillars. Proc. Natl. Acad. Sci. USA 2012, 109, 5328–5333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasapera, A.; Plotnikov, S.; Fischer, R.; Case, L.; Egelhoff, T.; Waterman, C. Rac1-Dependent Phosphorylation and Focal Adhesion Recruitment of Myosin IIA Regulates Migration and Mechanosensing. Curr. Biol. 2015, 25, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfenson, H.; Meacci, G.; Liu, S.; Stachowiak, M.R.; Iskratsch, T.; Ghassemi, S.; Roca-Cusachs, P.; O’Shaughnessy, B.; Hone, J.; Sheetz, M.P. Tropomyosin controls sarcomere-like contractions for rigidity sensing and suppressing growth on soft matrices. Nat. Cell Biol. 2016, 18, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizwan, M.; Peh, G.S.L.; Ang, H.P.; Lwin, N.C.; Adnan, K.; Mehta, J.S.; Tan, W.S.; Yim, E.K.F. Sequentially-crosslinked bioactive hydrogels as nano-patterned substrates with customizable stiffness and degradation for corneal tissue engineering applications. Biomaterials 2017, 120, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Tse, J.R.; Engler, A.J. Preparation of hydrogel substrates with tunable mechanical properties. Curr. Protoc. Cell Biol. 2010, 47. [Google Scholar] [CrossRef]
- Wong, J.; Velasco, A.; Rajagopalan, P.; Pham, Q. Directed Movement of Vascular Smooth Muscle Cells on Gradient-Compliant Hydrogels. Langmuir 2003, 19, 1908–1913. [Google Scholar] [CrossRef]
- Tomba, C.; Migdal, C.; Fuard, D.; Villard, C.; Nicolas, A. Poly-L-lysine/laminin surface coating reverses glail cell mechanosensitivity on stiffness-patterned hydrogels. submitted.
- Tian, M.; Li, Y.; Liu, W.; Jin, L.; Jiang, X.; Wang, X.; Ding, Z.; Peng, Y.; Zhou, J.; Fan, J.; et al. The nanomechanical signature of liver cancer tissues and its molecular origin. Nanoscale 2015, 7, 12998–13010. [Google Scholar] [CrossRef]
- Ciasca, G.; Sassun, T.E.; Minelli, E.; Antonelli, M.; Papi, M.; Santoro, A.; Giangaspero, F.; Delfini, R.; De Spirito, M. Nano-mechanical signature of brain tumours. Nanoscale 2016, 8, 19629–19643. [Google Scholar] [CrossRef]
- Feng, J.; Song, Q.; Zhang, B.; Wu, Y.; Wang, T.; Jiang, L. Large-Scale, Long-Range-Ordered Patterning of Nanocrystals via Capillary-Bridge Manipulation. Adv. Mater. 2017, 29. [Google Scholar] [CrossRef]
- Versaevel, M.; Grevesse, T.; Riaz, M.; Lantoine, J.; Gabriele, S. Micropatterning hydroxy-PAAm hydrogels and Sylgard 184 silicone elastomers with tunable elastic moduli. Methods Cell Biol. 2014, 121, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Azioune, A.; Carpi, N.; Tseng, Q.; Théry, M.; Piel, M. Protein micropatterns a direct printing protocol using deep UVs. Methods Cell Biol. 2010, 97, 133–146. [Google Scholar] [CrossRef]
- Tseng, Q.; Wang, I.; Duchemin-Pelletier, E.; Azioune, A.; Carpi, N.; Gao, J.; Filhol, O.; Piel, M.; Théry, M.; Balland, M. A new micropatterning method of soft substrates reveals that different tumorigenic signals can promote or reduce cell contraction levels. Lab. Chip 2011, 11, 2231–2240. [Google Scholar] [CrossRef]
- Murrell, M.; Kamm, R.; Matsudaira, P. Substrate viscosity enhances correlation in epithelial sheet movement. Biophys. J. 2011, 101, 297–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colin-York, H.; Eggeling, C.; Fritzsche, M. Dissection of mechanical force in living cells by super-resolved traction force microscopy. Nat. Protoc. 2017, 12, 783. [Google Scholar] [CrossRef]
- Califano, J.P.; Reinhart-King, C.A. Substrate Stiffness and Cell Area Predict Cellular Traction Stresses in Single Cells and Cells in Contact. Cell. Mol. Bioeng. 2010, 3, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izquierdo-Alvarez, A.; Vargas, D.A.; Jorge-Penas, A.; Subramani, R.; Vaeyens, M.M.; Van Oosterwyck, H. Spatiotemporal Analyses of Cellular Tractions Describe Subcellular Effect of Substrate Stiffness and Coating. Ann. Biomed. Eng. 2019, 47, 624–637. [Google Scholar] [CrossRef]
- Stricker, J.; Sabass, B.; Schwarz, U.S.; Gardel, M.L. Optimization of traction force microscopy for micron-sized focal adhesions. J. Phys. Condens. Matter 2010, 22, 194104. [Google Scholar] [CrossRef] [Green Version]
- Zamir, E.; Katz, B.Z.; Aota, S.; Yamada, K.M.; Geiger, B.; Kam, Z. Molecular diversity of cell-matrix adhesions. J. Cell Sci. 1999, 112, 1655–1669. [Google Scholar] [CrossRef]
- Tee, S.Y.; Fu, J.; Chen, C.S.; Janmey, P.A. Cell shape and substrate rigidity both regulate cell stiffness. Biophys. J. 2011, 100, L25–L27. [Google Scholar] [CrossRef] [Green Version]
- Oakes, P.W.; Banerjee, S.; Marchetti, M.C.; Gardel, M.L. Geometry regulates traction stresses in adherent cells. Biophys. J. 2014, 107, 825–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotnikov, S.V.; Pasapera, A.M.; Sabass, B.; Waterman, C.M. Force fluctuations within focal adhesions mediate ECM-rigidity sensing to guide directed cell migration. Cell 2012, 151, 1513–1527. [Google Scholar] [CrossRef] [Green Version]
- Sarangi, B.R.; Gupta, M.; Doss, B.L.; Tissot, N.; Lam, F.; Mège, R.M.; Borghi, N.; Ladoux, B. Coordination between Intra- and Extracellular Forces Regulates Focal Adhesion Dynamics. Nano Lett. 2017, 17, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas, A.; Geiger, B.; Safran, S.A. Cell mechanosensitivity controls the anisotropy of focal adhesions. Proc. Natl. Acad. Sci. USA 2004, 101, 12520–12525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shemesh, T.; Geiger, B.; Bershadsky, A.D.; Kozlov, M. Focal adhesions as mechanosensors: A physical mechanism. Proc. Natl. Acad. Sci. USA 2005, 102, 12383–12388. [Google Scholar] [CrossRef] [Green Version]
- Olberding, J.E.; Thouless, M.D.; Arruda, E.M.; Garikipati, K. The non-equilibrium thermodynamics and kinetics of focal adhesion dynamics. PLoS ONE 2010, 5, e12043. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.; Lin, M.; Huang, G.; Li, Y.; Ji, B.; Genin, G.M.; Deshpande, V.S.; Lu, T.J.; Xu, F. Cellular Mechanosensing of the Biophysical Microenvironment: A Review of Mathematical Models of Biophysical Regulation of Cell Responses. Phys. Life Rev. 2017, 22–23, 88–119. [Google Scholar] [CrossRef]
- Fouchard, J.; Mitrossilis, D.; Asnacios, A. Acto-myosin based response to stiffness and rigidity sensing. Cell Adh. Migr. 2010, 5, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Trichet, L.; Digabel, J.L.; Hawkins, R.J.; Vedula, S.R.K.; Gupta, M.; Ribrault, C.; Hersen, P.; Voituriez, R.; Ladoux, B. Evidence of a large-scale mechanosensing mechanism for cellular adaptation to substrate stiffness. Proc. Natl. Acad. Sci. USA 2012, 109, 6933–6938. [Google Scholar] [CrossRef] [Green Version]
- Gupta, M.; Sarangi, B.R.; Deschamps, J.; Nematbakhsh, Y.; Callan-Jones, A.; Margadant, F.; Mège, R.M.; Lim, C.T.; Voituriez, R.; Ladoux, B. Adaptive rheology and ordering of cell cytoskeleton govern matrix rigidity sensing. Nat. Commun. 2015, 6, 7525. [Google Scholar] [CrossRef]
- Tan, J.L.; Tien, J.; Pirone, D.M.; Gray, D.S.; Bhadriraju, K.; Chen, C.S. Cells lying on a bed of microneedles: An approach to isolate mechanical force. Proc. Natl. Acad. Sci. USA 2003, 100, 1484–1489. [Google Scholar] [CrossRef] [Green Version]
- Saez, A.; Buguin, A.; Silberzan, P.; Ladoux, B. Is the mechanical activity of epithelial cells controlled by deformations or forces? Biophys. J. 2005, 89, L52–L54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prager-Khoutorsky, M.; Lichtenstein, A.; Krishnan, R.; Rajendran, K.; Mayo, A.; Kam, Z.; Geiger, B.; Bershadsky, A.D. Fibroblast polarization is a matrix-rigidity-dependent process controlled by focal adhesion mechanosensing. Nat. Cell Biol. 2011, 13, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Janković, B.; Adnađević, B.; Jovanović, J. Isothermal kinetics of dehydration of equilibrium swollen poly (acrylic acid) hydrogel. J. Therm. Anal. Calorim. 2008, 92, 821–827. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mgharbel, A.; Migdal, C.; Bouchonville, N.; Dupenloup, P.; Fuard, D.; Lopez-Soler, E.; Tomba, C.; Courçon, M.; Gulino-Debrac, D.; Delanoë-Ayari, H.; et al. Cells on Hydrogels with Micron-Scaled Stiffness Patterns Demonstrate Local Stiffness Sensing. Nanomaterials 2022, 12, 648. https://doi.org/10.3390/nano12040648
Mgharbel A, Migdal C, Bouchonville N, Dupenloup P, Fuard D, Lopez-Soler E, Tomba C, Courçon M, Gulino-Debrac D, Delanoë-Ayari H, et al. Cells on Hydrogels with Micron-Scaled Stiffness Patterns Demonstrate Local Stiffness Sensing. Nanomaterials. 2022; 12(4):648. https://doi.org/10.3390/nano12040648
Chicago/Turabian StyleMgharbel, Abbas, Camille Migdal, Nicolas Bouchonville, Paul Dupenloup, David Fuard, Eline Lopez-Soler, Caterina Tomba, Marie Courçon, Danielle Gulino-Debrac, Héléne Delanoë-Ayari, and et al. 2022. "Cells on Hydrogels with Micron-Scaled Stiffness Patterns Demonstrate Local Stiffness Sensing" Nanomaterials 12, no. 4: 648. https://doi.org/10.3390/nano12040648