
MicroRNAs as a Suitable Biomarker to Detect the Effects of Long-Term Exposures to Nanomaterials. Studies on TiO2NP and MWCNT

Abstract
:1. Introduction
2. Materials and Methods
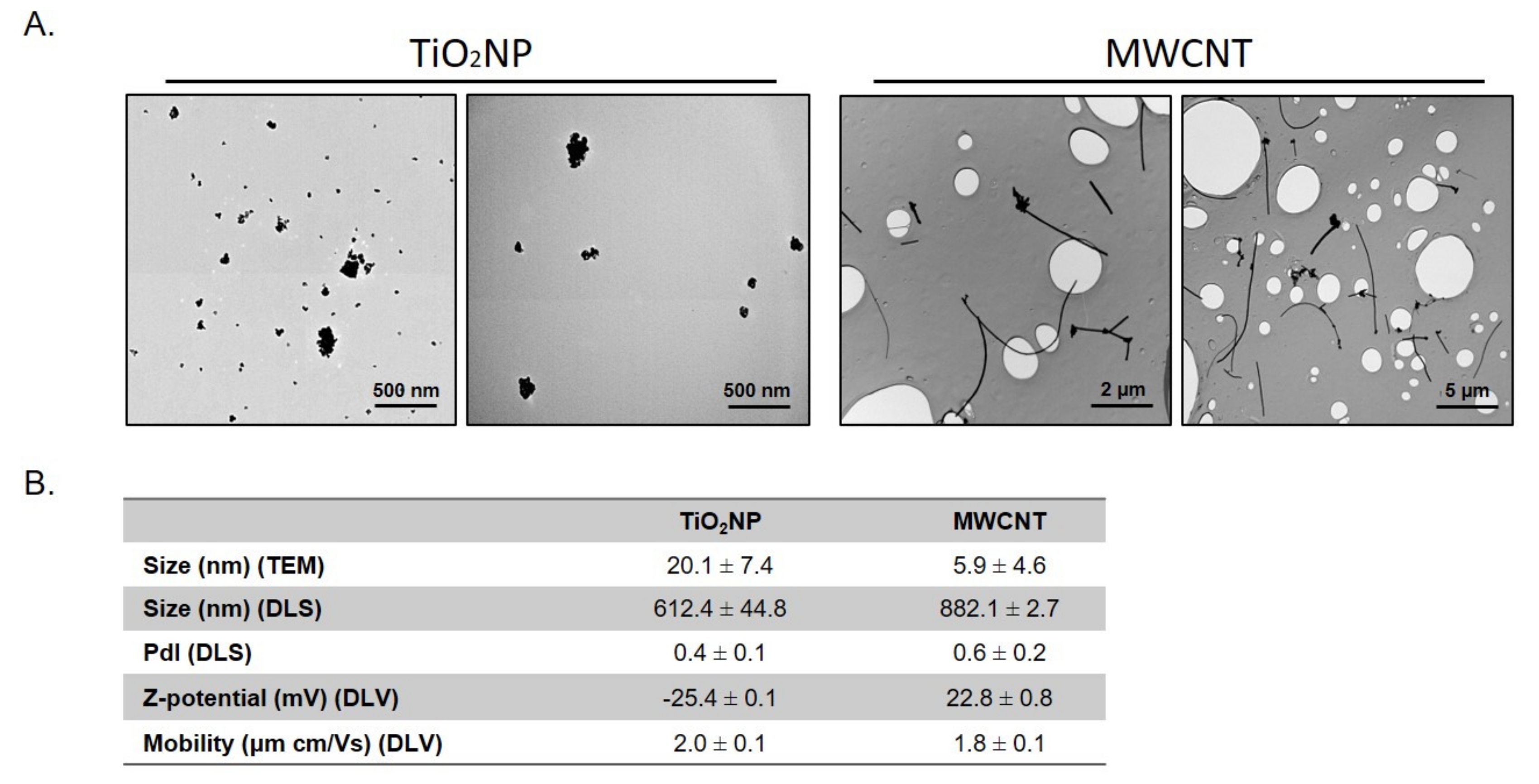
2.1. Nanomaterials Characterization
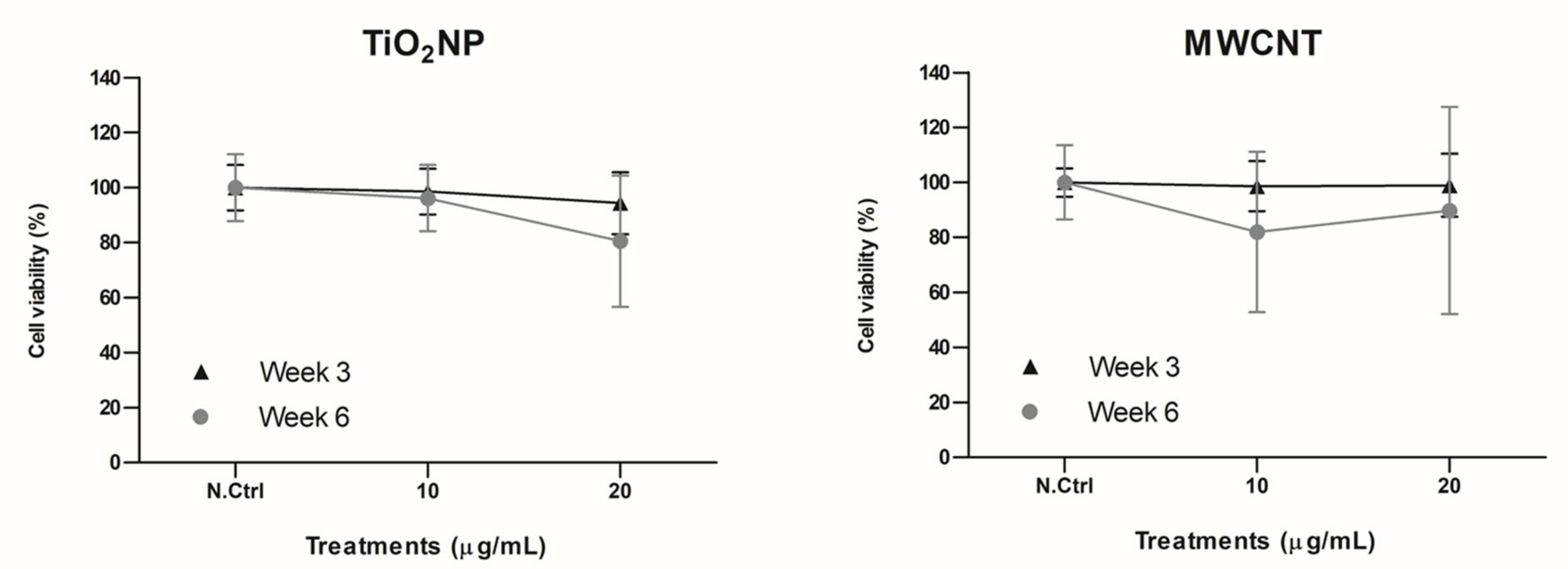
2.2. Cell Culture and Exposure Conditions
2.3. RNA Extraction and Retrotranscription Experiments
2.4. Real-Time RT-PCR
2.5. Pathway-Enrichment Analysis
2.6. Validation of the Selected Battery of MicroRNAs as Biomarkers of Long-Term Effects of NMs
3. Results
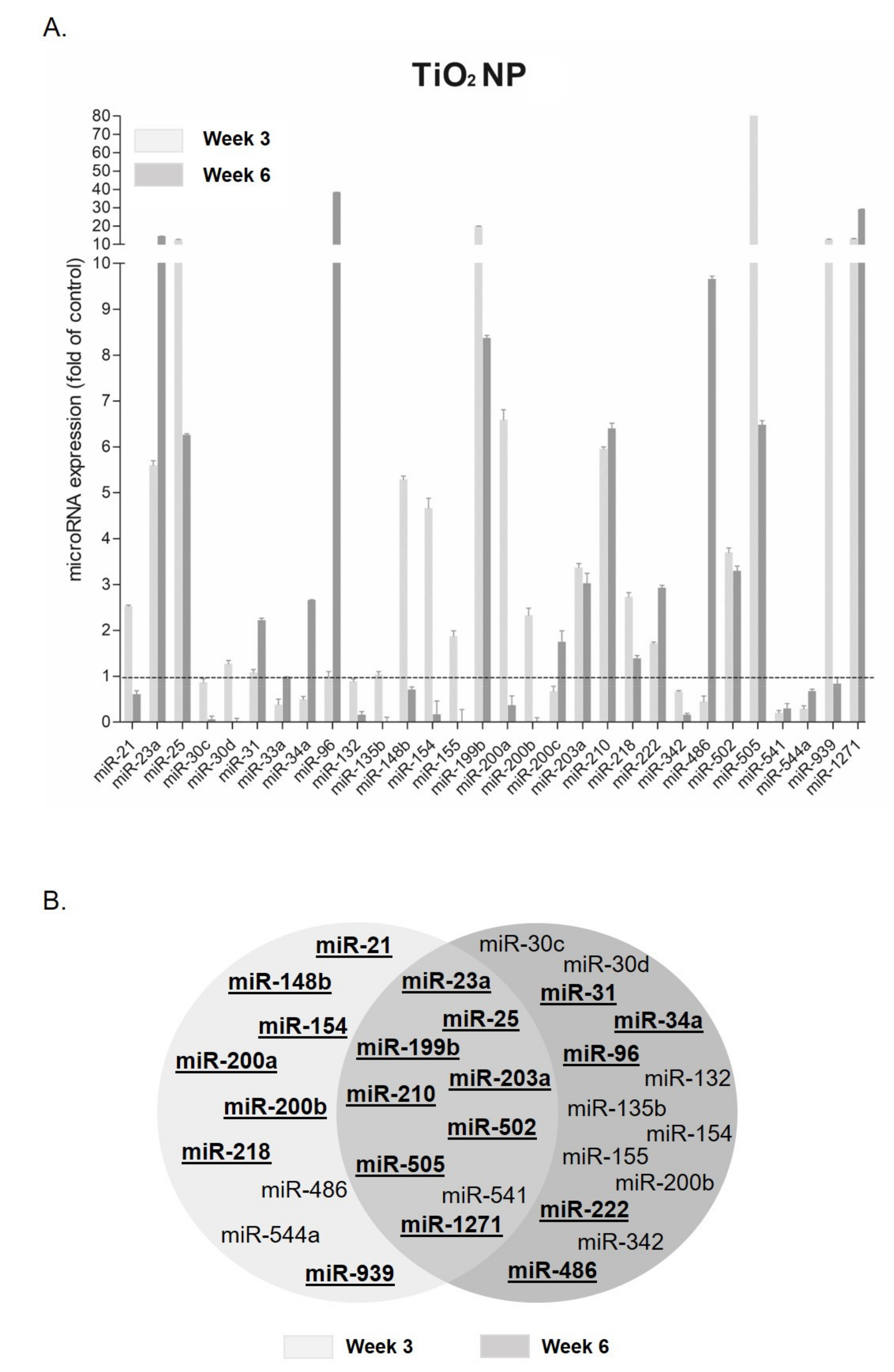
3.1. MicroRNA Expression Changes after TiO2 NP Exposure
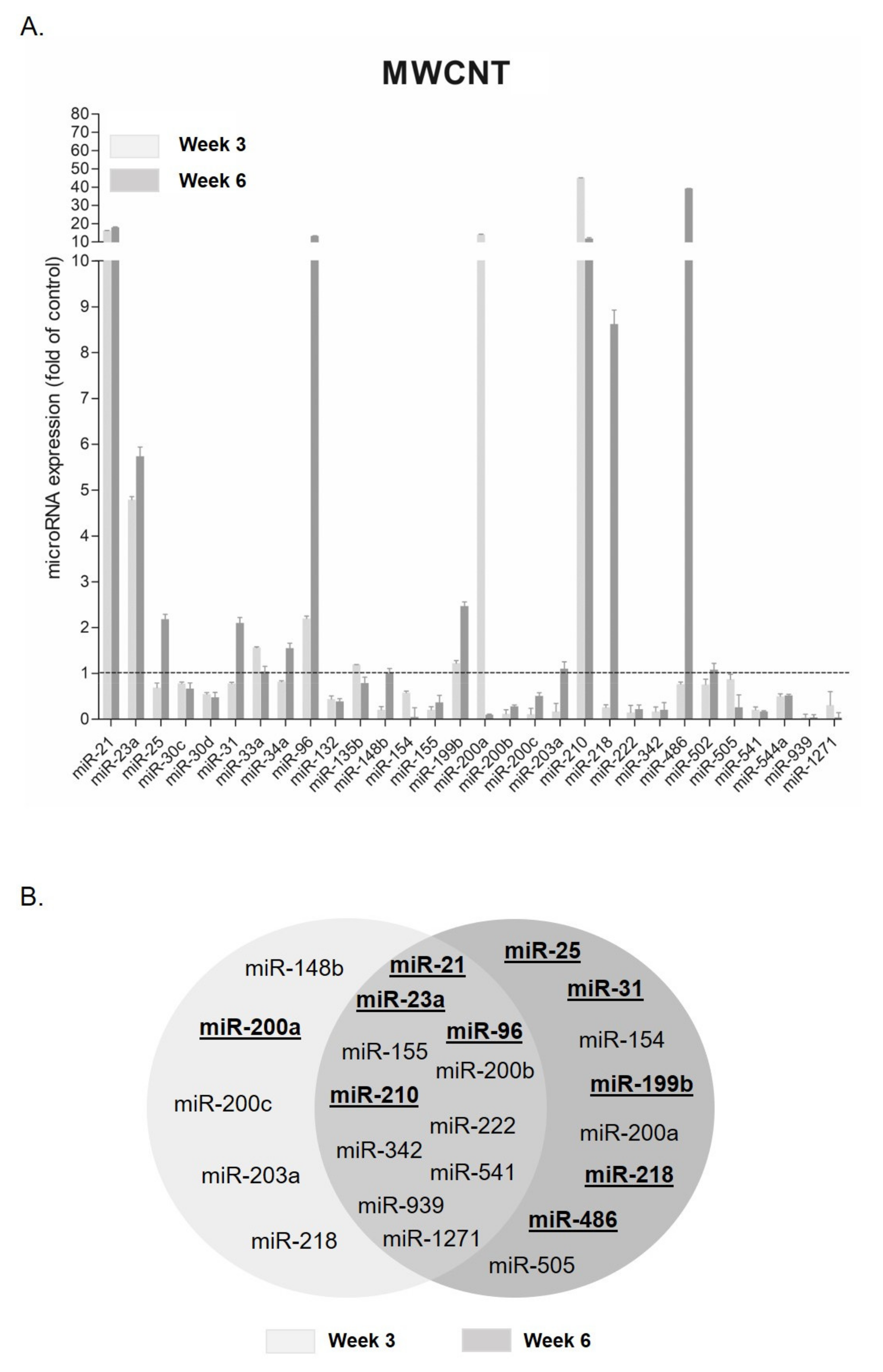
3.2. MicroRNA Expression Changes after MWCNT Exposure
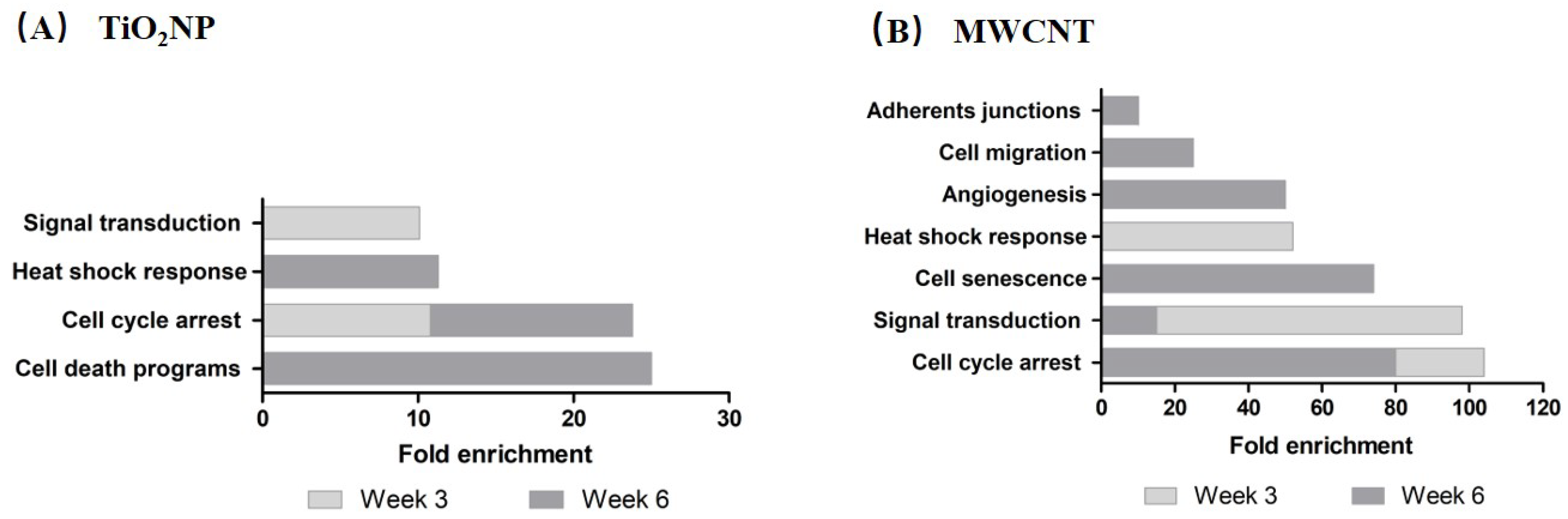
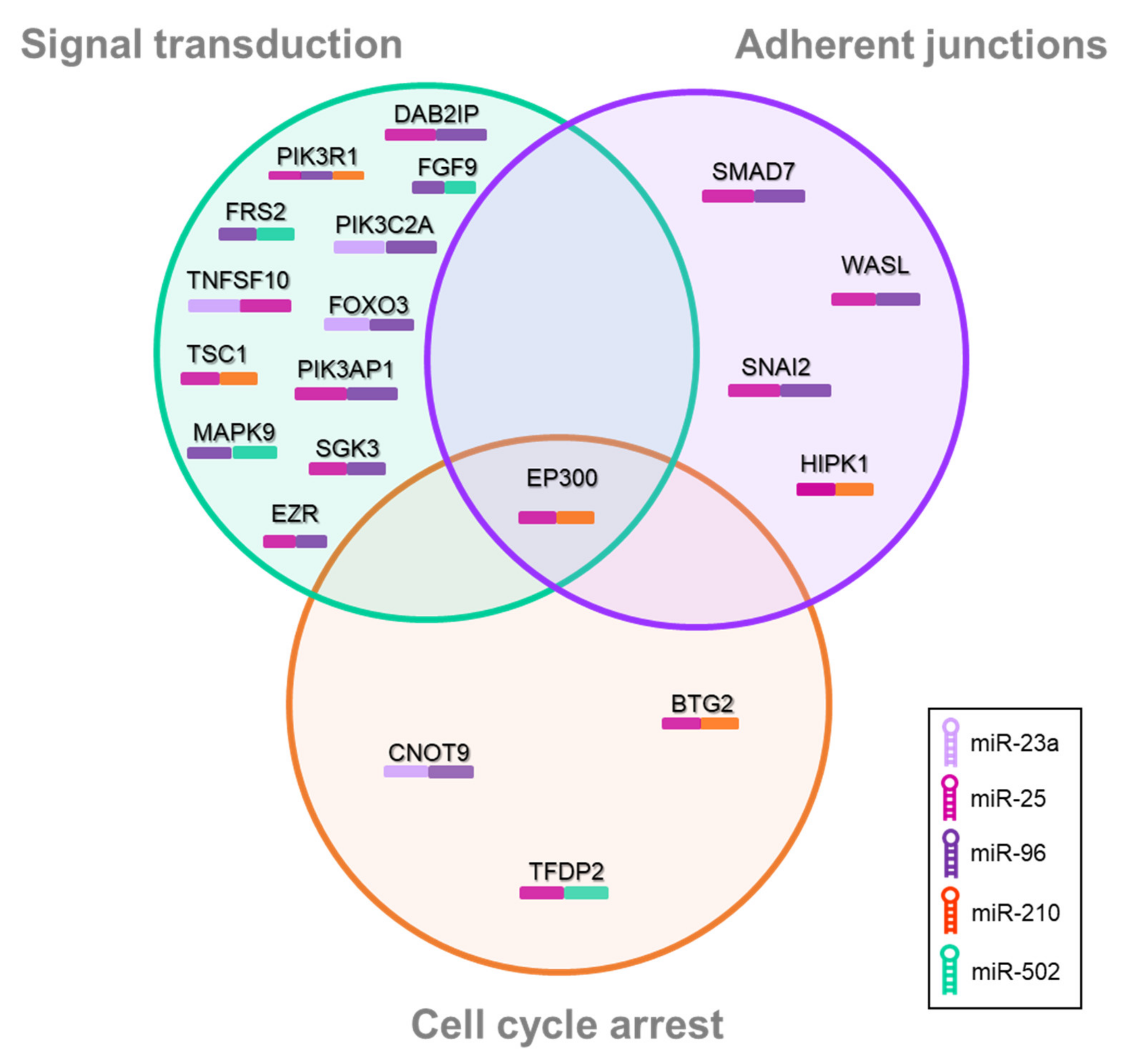
3.3. Pathway-Enrichment Analysis
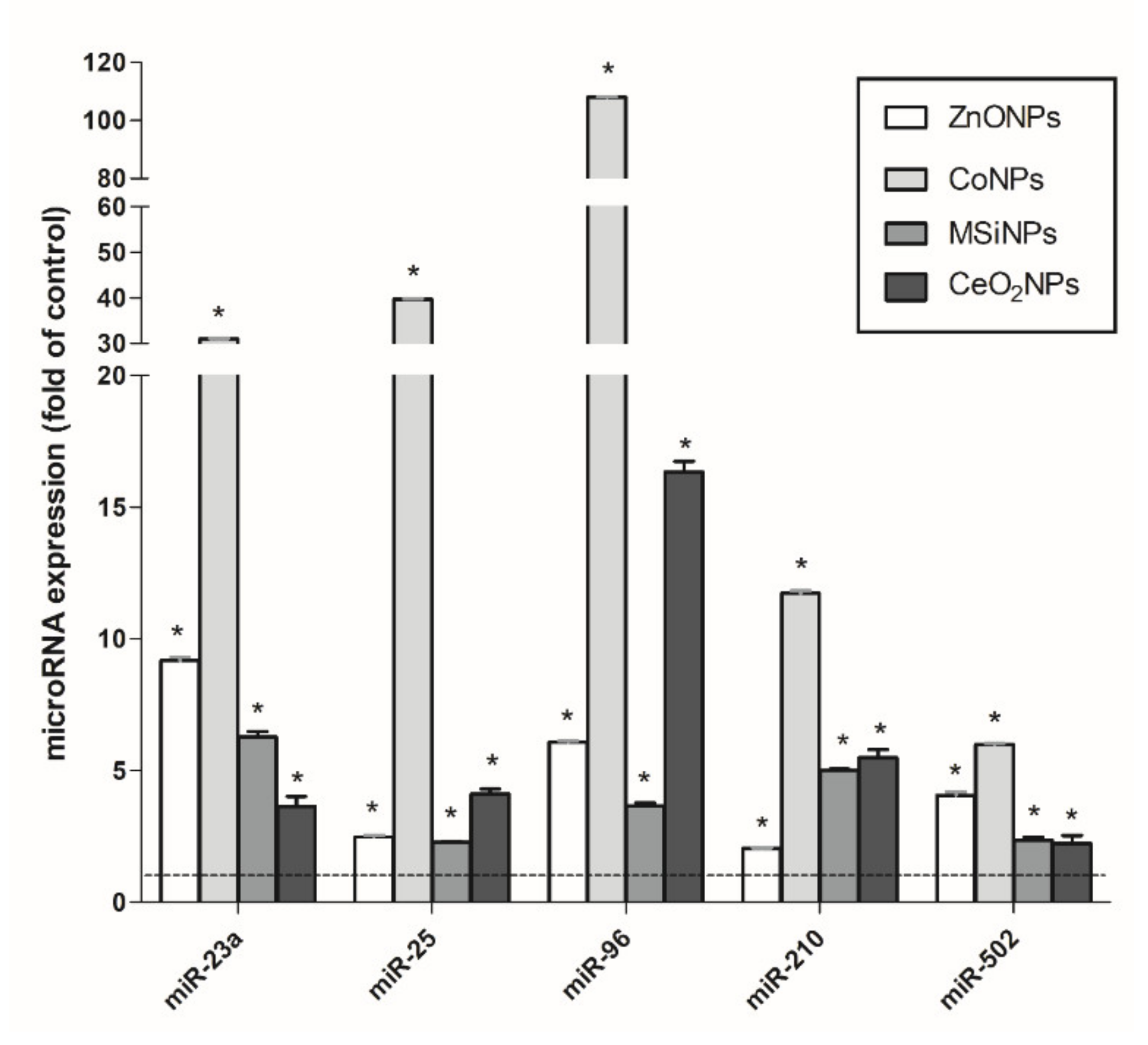
3.4. Validation of a Set of MicroRNAs as Informative Biomarkers of Long-Term Effects of NMs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Competing Financial Interests
References
- Bimová, P.; Barbieriková, Z.; Grenčíková, A.; Šípoš, R.; Škulcová, A.B.; Krivjanská, A.; Mackuľak, T. Environmental risk of nanomaterials and nanoparticles and EPR technique as an effective tool to study them-a review. Environ. Sci. Pollut. Res. Int. 2021, 28, 22203–22220. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Chong, Y.; Ge, C. Understanding the nano-bio interactions and the corresponding biological responses. Front. Chem. 2020, 8, 446. [Google Scholar] [CrossRef] [PubMed]
- Freixa, A.; Acuña, V.; Sanchís, J.; Farré, M.; Barceló, D.; Sabater, S. Ecotoxicological effects of carbon based nanomaterials in aquatic organisms. Sci. Total Environ. 2018, 619–620, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Annangi, B.; Bach, J.; Vales, G.; Rubio, L.; Marcos, R.; Hernández, A. Long-term exposures to low doses of cobalt nanoparticles induce cell transformation enhanced by oxidative damage. Nanotoxicology 2015, 9, 138–147. [Google Scholar] [CrossRef]
- Annangi, B.; Rubio, L.; Alaraby, M.; Bach, J.; Marcos, R.; Hernández, A. Acute and long-term in vitro effects of zinc oxide nanoparticles. Arch. Toxicol. 2016, 90, 2201–2213. [Google Scholar] [CrossRef]
- Vila, L.; Marcos, R.; Hernández, A. Long-term effects of silver nanoparticles in Caco-2 cells. Nanotoxicology 2017, 11, 771–780. [Google Scholar] [CrossRef]
- Barthel, H.; Darne, C.; Gaté, L.; Visvikis, A.; Seidel, C. Continuous long-term exposure to low concentrations of MWCNTs induces an epithelial-mesenchymal transition in BEAS-2B cells. Nanomaterials 2021, 11, 1742. [Google Scholar] [CrossRef]
- Sierra, M.I.; Rubio, L.; Bayón, G.F.; Cobo, I.; Menendez, P.; Morales, P.; Mangas, C.; Urdinguio, R.G.; Lopez, V.; Valdes, A.; et al. DNA methylation changes in human lung epithelia cells exposed to multi-walled carbon nanotubes. Nanotoxicology 2017, 11, 857–870. [Google Scholar] [CrossRef]
- Smolkova, B.; Dusinska, M.; Gabelova, A. Nanomedicine and epigenome. Possible health risks. Food Chem. Toxicol. 2017, 109, 780–796. [Google Scholar] [CrossRef]
- Yu, J.; Loh, X.J.; Luo, Y.; Ge, S.; Fan, X.; Ruan, J. Insights into the epigenetic effects of nanomaterials on cells. Biomater. Sci. 2020, 8, 763–775. [Google Scholar] [CrossRef]
- Pogribna, M.; Hammons, G. Epigenetic effects of nanomaterials and nanoparticles. J. Nanobiotechnol. 2021, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; Chakraborty, A.; Sarkar, D.; Langthasa, M.; Rahman, M.; Bari, M.; Singha, R.K.S.; Malakar, A.K.; Chakraborty, S. Interplay between miRNAs and human diseases. J. Cell. Physiol. 2018, 233, 2007–2018. [Google Scholar] [CrossRef]
- Huang, Q.; Liu, Y.; Dong, S. Emerging roles of long non-coding RNAs in the toxicology of environmental chemicals. J. Appl. Toxicol. 2018, 38, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Chekhun, V.F. MicroRNAs are a key factor in the globalization of tumor-host relationships. Exp. Oncol. 2019, 41, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, S.; Barguilla, I.; Marcos, R.; Hernández, A. Ex vivo long-term in vitro exposures to nanoceria, alone or in combination with cigarette-smoke condensate, induce transforming and epigenetic cancer-like features. Nanomedicine 2021, 16, 293–305. [Google Scholar] [CrossRef]
- George, G.P.; Mittal, R.D. MicroRNAs: Potential biomarkers in cancer. Ind. J. Clin. Biochem. 2010, 25, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vales, G.; Rubio, L.; Marcos, R. Long-term exposures to low doses of titanium dioxide nanoparticles induce cell transformation, but not genotoxic damage in BEAS-2B cells. Nanotoxicology 2015, 9, 568–578. [Google Scholar] [CrossRef]
- Vales, G.; Rubio, L.; Marcos, R. Genotoxic and cell-transformation effects of multi-walled carbon nanotubes (MWCNT) following in vitro sub-chronic exposures. J. Hazard. Mater. 2016, 306, 193–202. [Google Scholar] [CrossRef]
- Shabbir, S.; Kulyar, M.F.; Bhutta, Z.A.; Boruah, P.; Asif, M. Toxicological consequences of titanium dioxide nanoparticles (TiO2NPs) and their jeopardy to human population. BioNanoScience 2021, 26, 621–632. [Google Scholar] [CrossRef]
- Saleemi, M.A.; Hosseini, F.M.; Yong, P.V.C.; Chinna, K.; Palanisamy, N.K.; Wong, E.H. Toxicity of carbon nanotubes: Molecular mechanisms, signaling cascades, and remedies in biomedical applications. Chem. Res. Toxicol. 2021, 34, 24–46. [Google Scholar] [CrossRef] [PubMed]
- Nanogenotox. 2011. Available online: https://www.anses.fr/en/system/files/nanogenotox_deliverable_5.pdf (accessed on 2 November 2021).
- Balcells, I.; Cirera, S.; Busk, P.K. Specific and sensitive quantitative RT-PCR of miRNAs with DNA primers. BMC Biotechnol. 2011, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Busk, P.K. A tool for design of primers for microRNA-specific quantitative RT-qPCR. BMC Bioinform. 2014, 15, 29. [Google Scholar] [CrossRef] [Green Version]
- Khanna, P.; Ong, C.; Bay, B.; Baeg, G. Nanotoxicity: An interplay of oxidative stress, inflammation and cell death. Nanomaterials 2015, 5, 1163–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramanathan, A. Toxicity of nanoparticles challenges and opportunities. Appl. Microsc. 2019, 49, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gliga, A.R.; Bucchianico, S.D.; Åkerlund, E.; Karlsson, H.L. Transcriptome profiling and toxicity following long-term, low dose exposure of human lung cells to Ni and NiO nanoparticles-comparison with NiCl2. Nanomaterials 2020, 10, 649. [Google Scholar] [CrossRef] [Green Version]
- Sierra, M.I.; Valdés, A.; Fernandez, A.F.; Torrecillas, R.; Fraga, M.F. The effect of exposure to nanoparticles and nanomaterials on the mammalian epigenome. Int. J. Nanomed. 2016, 11, 6297–6306. [Google Scholar] [CrossRef] [Green Version]
- Markou, A.; Zavridou, M.; Lianidou, E.S. miRNA-21 as a novel therapeutic target in lung cancer. Lung Cancer 2016, 7, 19–27. [Google Scholar] [PubMed] [Green Version]
- Zhao, M.Y.; Wang, L.M.; Liu, J.; Huang, X.; Liu, J.; Zhang, Y.F. MiR-21 suppresses anoikis through targeting PDCD4 and PTEN in human esophageal adenocarcinoma. Curr. Med. Sci. 2018, 38, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Bica-Pop, C.; Cojocneanu-Petric, R.; Magdo, L.; Raduly, L.; Gulei, D.; Berindan-Neagoe, I. Overview upon miR-21 in lung cancer: Focus on NSCLC. Cell. Mol. Life Sci. 2018, 75, 3539–3551. [Google Scholar] [CrossRef]
- Cao, M.; Seike, M.; Soeno, C.; Mizutani, H.; Kitamura, K.; Minegishi, Y.; Noro, R.; Yoshimura, A.; Cai, L.; Gemma, A. MiR-23a regulates TGF-β-induced epithelial-mesenchymal transition by targeting E-cadherin in lung cancer cells. Int. J. Oncol. 2012, 41, 869–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, T.Y.; Lee, M.S.; Mun, J.Y.; Ihm, C.; Kim, S.A. Exosome cargo reflects TGF-β1-mediated epithelial-to-mesenchymal transition (EMT) status in A549 human lung adenocarcinoma cells. Biochem. Biophys. Res. Commun. 2016, 478, 643–648. [Google Scholar] [CrossRef]
- Karacosta, L.G.; Anchang, B.; Ignatiadis, N.; Kimmey, S.C.; Benson, J.A.; Shrager, J.B.; Tibshirani, R.; Bendall, S.C.; Plevritis, S.K. Mapping lung cancer epithelial-mesenchymal transition states and trajectories with single-cell resolution. Nat. Commun. 2019, 10, 5587. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, P.; Chen, T.; Gao, G.; Chen, X.; Du, Y.; Gao, F.; Zhang, G. Expression of microRNA-96 and its potential functions by targeting FOXO3 in non-small cell lung cancer. Tumor Biol. 2015, 36, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Schaaij-Visser, T.B.; de Wit, M.; Lam, S.W.; Jiménez, C.R. The cancer secretome, current status and opportunities in the lung, breast and colorectal cancer context. Biochim. Biophys. Acta 2013, 1834, 2242–2258. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Li, X.; Dong, H.; Suo, L.; Zhang, J.; Zhang, L.; Zhang, J. miR2103p regulates the proliferation and apoptosis of nonsmall cell lung cancer cells by targeting SIN3A. Exp. Ther. Med. 2019, 18, 2565–2573. [Google Scholar] [PubMed]
- Guo, H.; Li, Q.; Li, W.; Zheng, T.; Zhao, S.; Liu, Z. miR-96 downregulates RECK to promote growth and motility of non-small cell lung cancer cells. Mol. Cell. Biochem. 2014, 390, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Sun, X. miR-25 promotes invasion of human non-small cell lung cancer via CDH1. Bioengineered 2019, 10, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Zhou, J.; Mei, S.; Wu, D.; Mu, Z.; Chen, B.; Xie, Y.; Ye, Y.; Liu, J. Circulating exosomal microRNA-96 promotes cell proliferation, migration and drug resistance by targeting LMO7. J. Cell. Mol. Med. 2017, 21, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or tumor suppressor? The duplicity of microRNAs in cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadeh-Fanalou, S.; Khosravi, M.; Alian, F.; Rokhsartalb-Azar, S.; Nazarizadeh, A.; Karimi-Dehkordi, M.; Mohammadi, F. Dual role of microRNA-1297 in the suppression and progression of human malignancies. Biomed. Pharmacother. 2021, 141, 111863. [Google Scholar] [CrossRef]
- Li, M.; Huo, X.; Davuljigari, C.B.; Dai, Q.; Xu, X. MicroRNAs and their role in environmental chemical carcinogenesis. Environ. Geochem. Health 2019, 41, 225–247. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, M.; Cristaldi, A.; Oliveri, C.G. Oncogenic role of miRNA in environmental exposure to plasticizers: A systematic review. J. Pers. Med. 2021, 11, 500. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FORWARD | REVERSE | |
|---|---|---|
| >hsa-miR-21-5p | TCAGTAGCTTATCAGACTGATG | CGTCCAGTTTTTTTTTTTTTTTCAAC |
| >hsa-miR-23a-5p | CATCACATTGCCAGGGAT | CGTCCAGTTTTTTTTTTTTTTTGGAA |
| >hsa-miR-25-3p | CATTGCACTTGTCTCGGT | GGTCCAGTTTTTTTTTTTTTTTCAG |
| >hsa-miR-30c-5p | GCAGTGTAAACATCCTACACTCT | TCCAGTTTTTTTTTTTTTTTGCTGA |
| >hsa-miR-30d-5p | AGTGTAAACATCCCCGACT | GGTCCAGTTTTTTTTTTTTTTTCTTC |
| >hsa-miR-31-5p | GCAGAGGCAAGATGCTG | GTCCAGTTTTTTTTTTTTTTTAGCTATG |
| >hsa-miR-33a-5p | CGCAGGTGCATTGTAGT | GTCCAGTTTTTTTTTTTTTTTGCAAT |
| >hsa-miR-34a-5p | GCAGTGGCAGTGTCTTAG | GGTCCAGTTTTTTTTTTTTTTTACAAC |
| >hsa-miR-96-5p | CAGTTTGGCACTAGCACA | GGTCCAGTTTTTTTTTTTTTTTAGCA |
| >hsa-miR-124-5p | GCAGCGTGTTCACAGC | TCCAGTTTTTTTTTTTTTTTATCAAGGT |
| >hsa-miR-126-5p | CGCAGCATTATTACTTTTGGT | CCAGTTTTTTTTTTTTTTTCGCGT |
| >hsa-miR-132-5p | ACCGTGGCTTTCGATTG | GGTCCAGTTTTTTTTTTTTTTTAGTAAC |
| >hsa-miR-135b-5p | GCAGTATGGCTTTTCATTCCT | GGTCCAGTTTTTTTTTTTTTTTCACA |
| >hsa-miR-148b-3p | GCAGTCAGTGCATCACAGA | GGTCCAGTTTTTTTTTTTTTTTACAAAG |
| >hsa-miR-154-5p | GCAGTAGGTTATCCGTGTTG | GTCCAGTTTTTTTTTTTTTTTCGAAG |
| >hsa-miR-155-5p | CGCAGTTAATGCTAATCGTGATAG | AGGTCCAGTTTTTTTTTTTTTTTACC |
| >hsa-miR-199b-5p | CAGCCCAGTGTTTAGACTATC | GTCCAGTTTTTTTTTTTTTTTGAACAG |
| >hsa-miR-200a | AGCATCTTACCGGACAGT | CCAGTTTTTTTTTTTTTTTCCAGCA |
| >hsa-miR-200b-5p | GCATCTTACTGGGCAGCA | GGTCCAGTTTTTTTTTTTTTTTCCAA |
| >hsa-miR-200c | CGTCTTACCCAGCAGTGT | GGTCCAGTTTTTTTTTTTTTTTCCA |
| >hsa-miR-203a-3p | CAGGTGAAATGTTTAGGACCA | GGTCCAGTTTTTTTTTTTTTTTCTAGT |
| >hsa-miR-210-5p | TGCCCACCGCACA | GGTCCAGTTTTTTTTTTTTTTTCAGT |
| >hsa-miR-218-5p | CGCAGTTGTGCTTGATCT | TCCAGTTTTTTTTTTTTTTTACATGGT |
| >hsa-miR-222-3p | GCAGAGCTACATCTGGCT | CCAGTTTTTTTTTTTTTTTACCCAGT |
| >hsa-miR-224-5p | GCAGCAAGTCACTAGTGGT | TCCAGTTTTTTTTTTTTTTTAACGGA |
| >hsa-miR-342-3p | GTCTCACACAGAAATCGCA | GGTCCAGTTTTTTTTTTTTTTTACG |
| >hsa-miR-486-3p | GGGGCAGCTCAGTACA | GGTCCAGTTTTTTTTTTTTTTTATCCT |
| >hsa-miR-502-3p | AATGCACCTGGGCAAG | GGTCCAGTTTTTTTTTTTTTTTGAATC |
| >hsa-miR-505-3p | CGTCAACACTTGCTGGT | GGTCCAGTTTTTTTTTTTTTTTAGGA |
| >hsa-miR-541-3p | GTGGTGGGCACAGAATC | CCAGTTTTTTTTTTTTTTTAGTCCAG |
| hsa-miR-544a | GCAGATTCTGCATTTTTAGCAAG | GGTCCAGTTTTTTTTTTTTTTTGAAC |
| >hsa-miR-939-3p | CCTGGGCCTCTGCTC | GGTCCAGTTTTTTTTTTTTTTTCTG |
| >hsa-miR-1271-3p | TGCCTGCTATGTGCCA | TCCAGTTTTTTTTTTTTTTTGCCT |
| >U6 small nuclear RNA | CTCGCTTCGGCAGCACA | AACGCTTCACGAATTTGCGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballesteros, S.; Vales, G.; Velázquez, A.; Pastor, S.; Alaraby, M.; Marcos, R.; Hernández, A. MicroRNAs as a Suitable Biomarker to Detect the Effects of Long-Term Exposures to Nanomaterials. Studies on TiO2NP and MWCNT. Nanomaterials 2021, 11, 3458. https://doi.org/10.3390/nano11123458
Ballesteros S, Vales G, Velázquez A, Pastor S, Alaraby M, Marcos R, Hernández A. MicroRNAs as a Suitable Biomarker to Detect the Effects of Long-Term Exposures to Nanomaterials. Studies on TiO2NP and MWCNT. Nanomaterials. 2021; 11(12):3458. https://doi.org/10.3390/nano11123458
Chicago/Turabian StyleBallesteros, Sandra, Gerard Vales, Antonia Velázquez, Susana Pastor, Mohamed Alaraby, Ricard Marcos, and Alba Hernández. 2021. "MicroRNAs as a Suitable Biomarker to Detect the Effects of Long-Term Exposures to Nanomaterials. Studies on TiO2NP and MWCNT" Nanomaterials 11, no. 12: 3458. https://doi.org/10.3390/nano11123458