
Nanostructured Modifications of Titanium Surfaces Improve Vascular Regenerative Properties of Exosomes Derived from Mesenchymal Stem Cells: Preliminary In Vitro Results


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fabrication of Nanostructured Surfaces
2.2. Characterization of Nanostructured Surfaces
2.3. Culture of MSC onto TNs
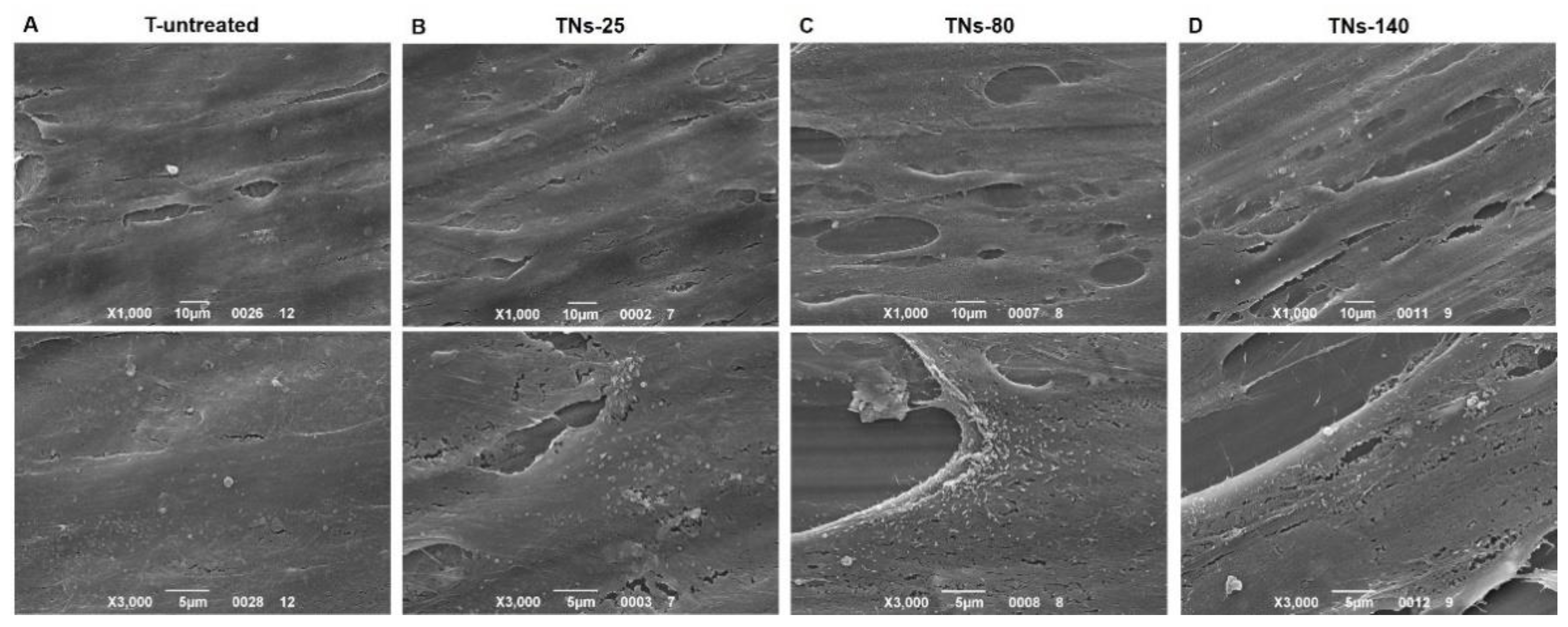
2.4. SEM
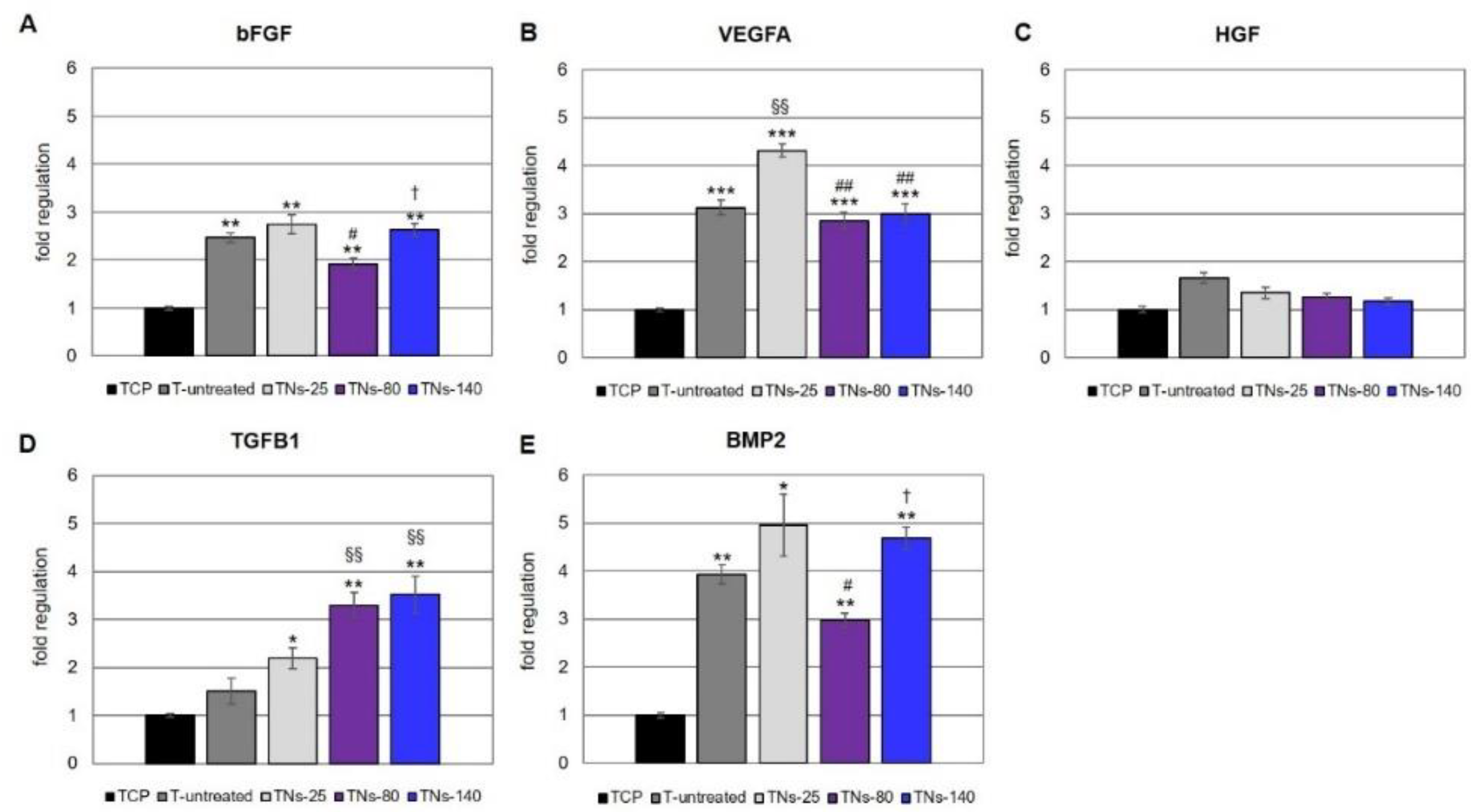
2.5. RNA Isolation from hMSCs, First-Strand cDNA Synthesis, and Real-Time PCR
2.6. Exosomes Isolation from Cells Grown onto TNs
2.7. Characterization of exo-hMSCs
2.8. Exosomes Labeling and Internalization by Endothelial Cells
2.9. Wound Healing Migration Assay
2.10. RNA Isolation from HUVECs, First-Strand cDNA Synthesis, and Real-Time PCR
2.11. Isolation of Total RNA from exo-hMSCs and miScript® miRNA PCR Arrays
2.12. Statistical Analysis
3. Results
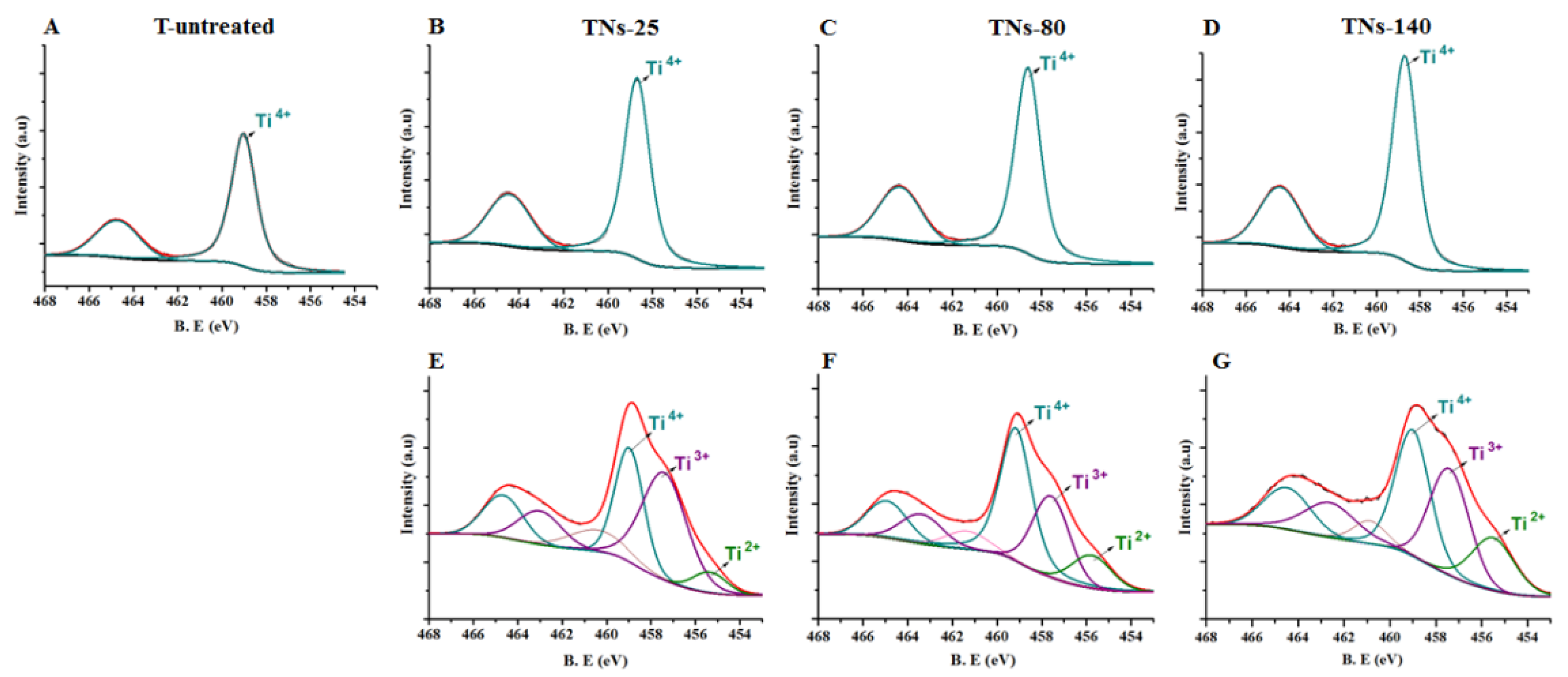
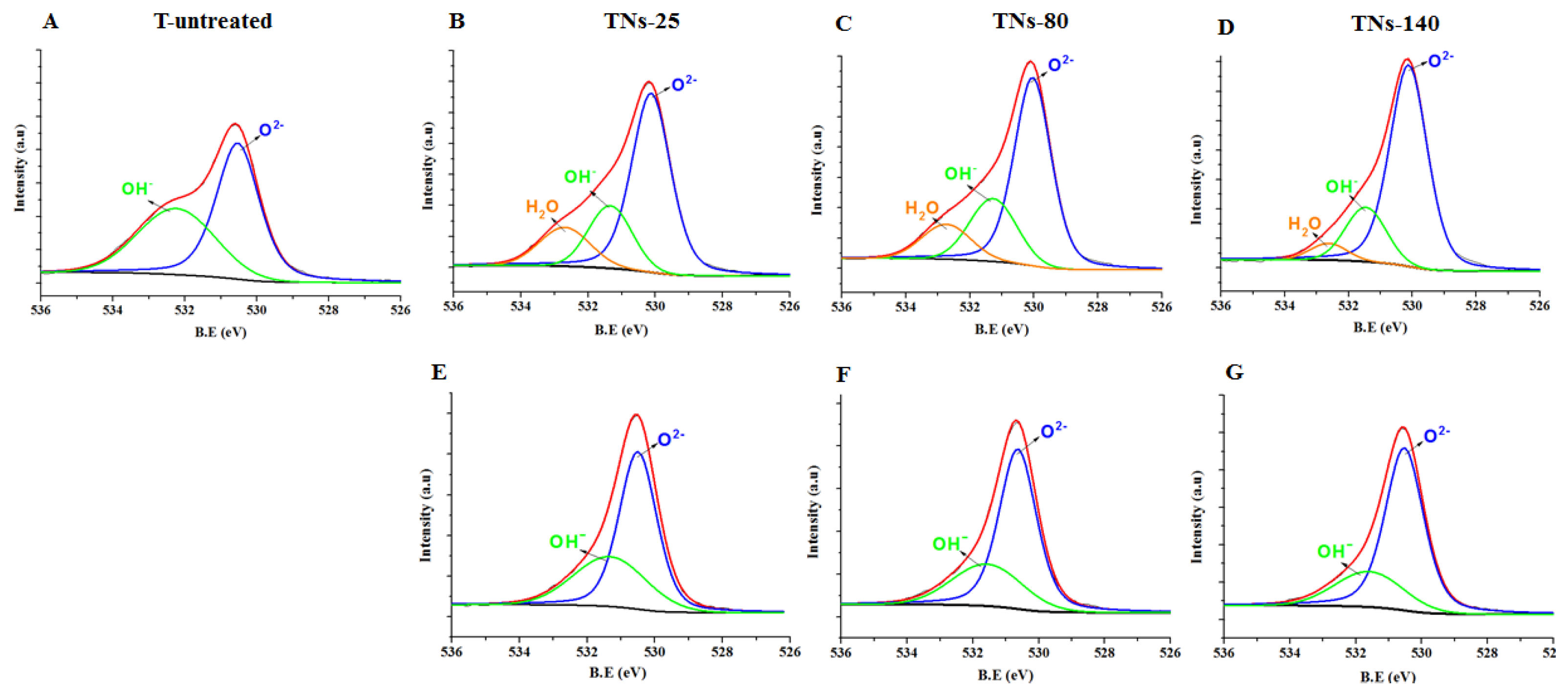
3.1. Characterizations of Control and Nanostructured Surfaces
3.2. Characterization of hMSCs Grown onto TNs
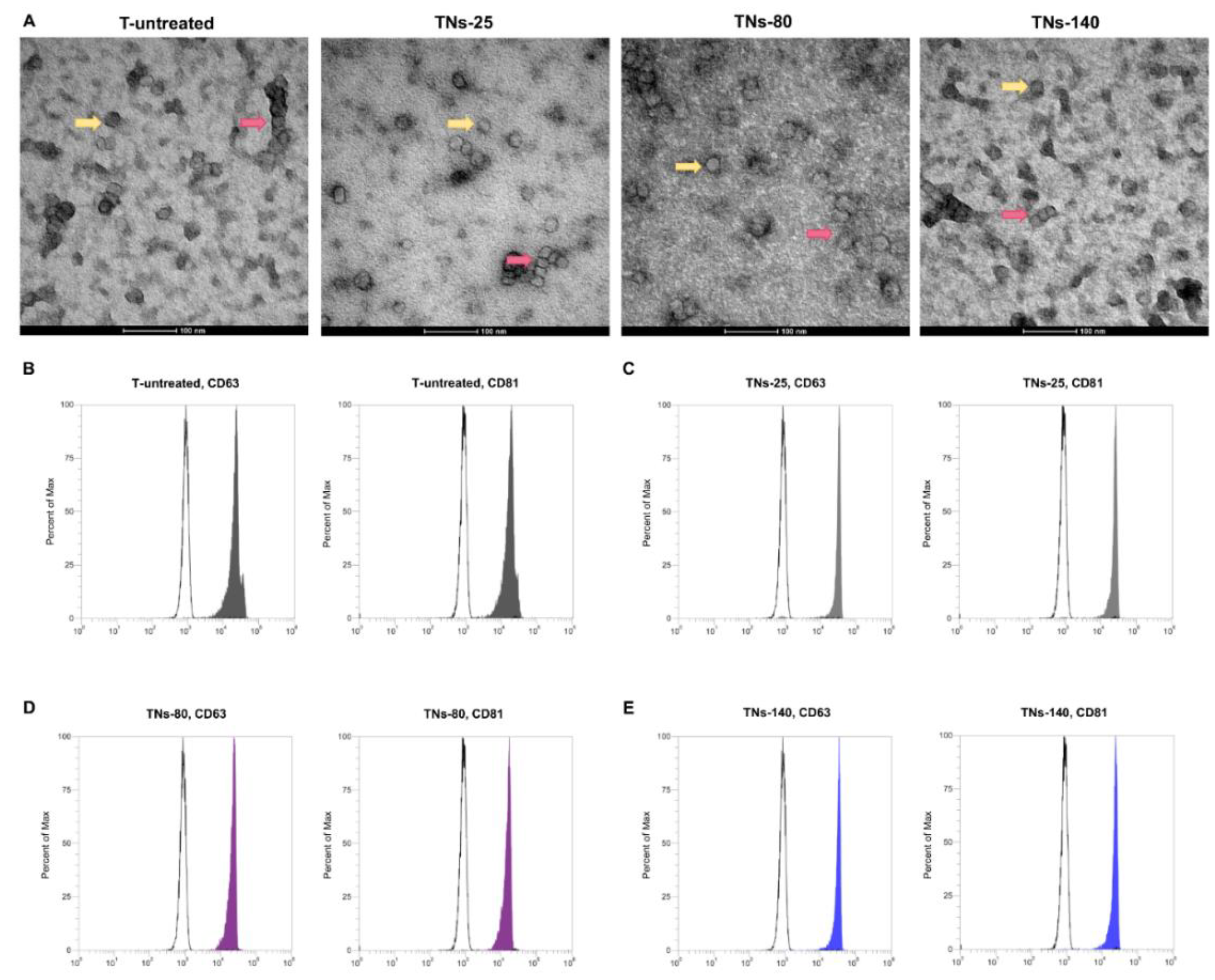
3.3. Characterization of Exosomes Recovered from hMSCs Seeded onto TNs
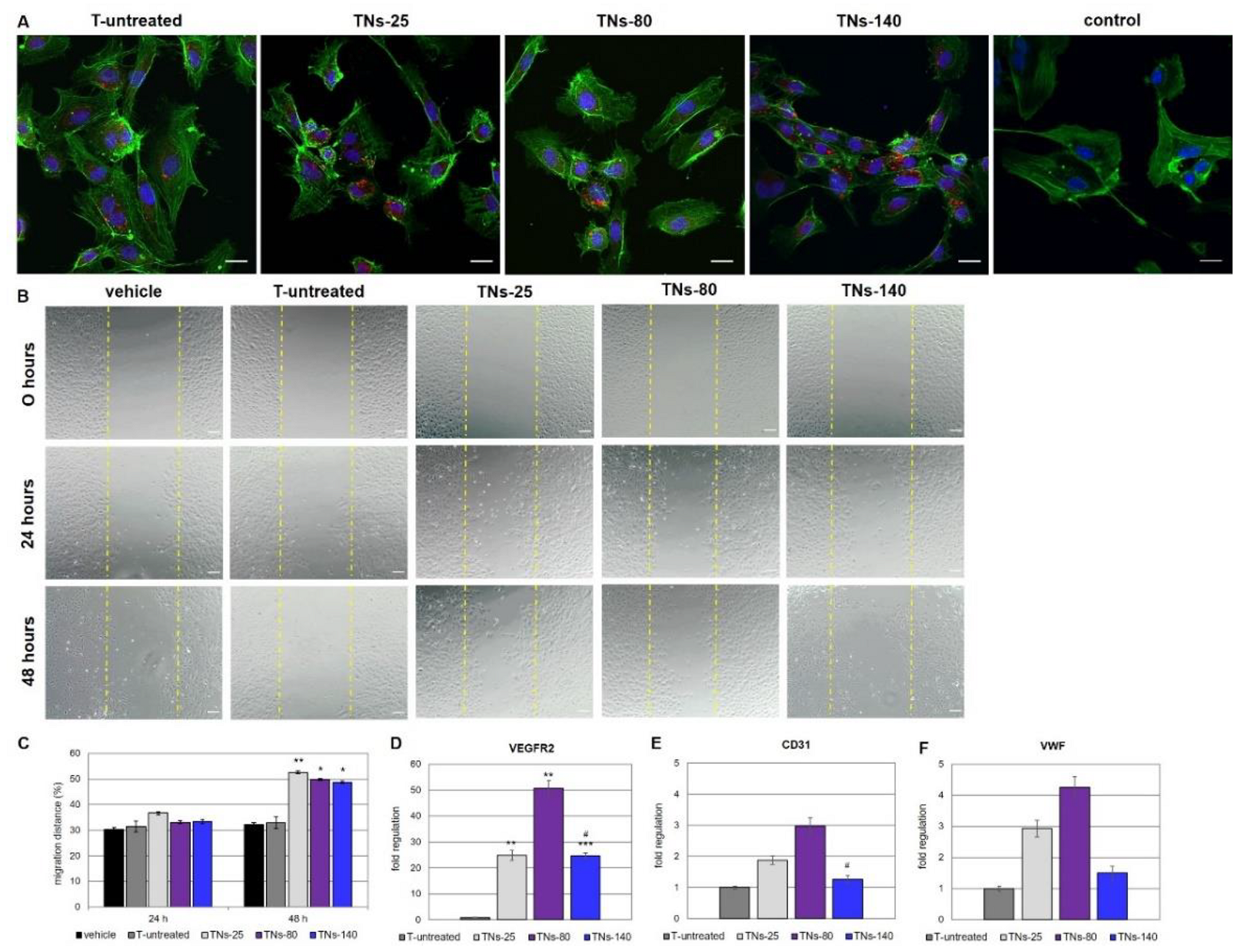
3.4. Effects of hMSCs-Derived Exosomes on the Behavior of Endothelial Cells
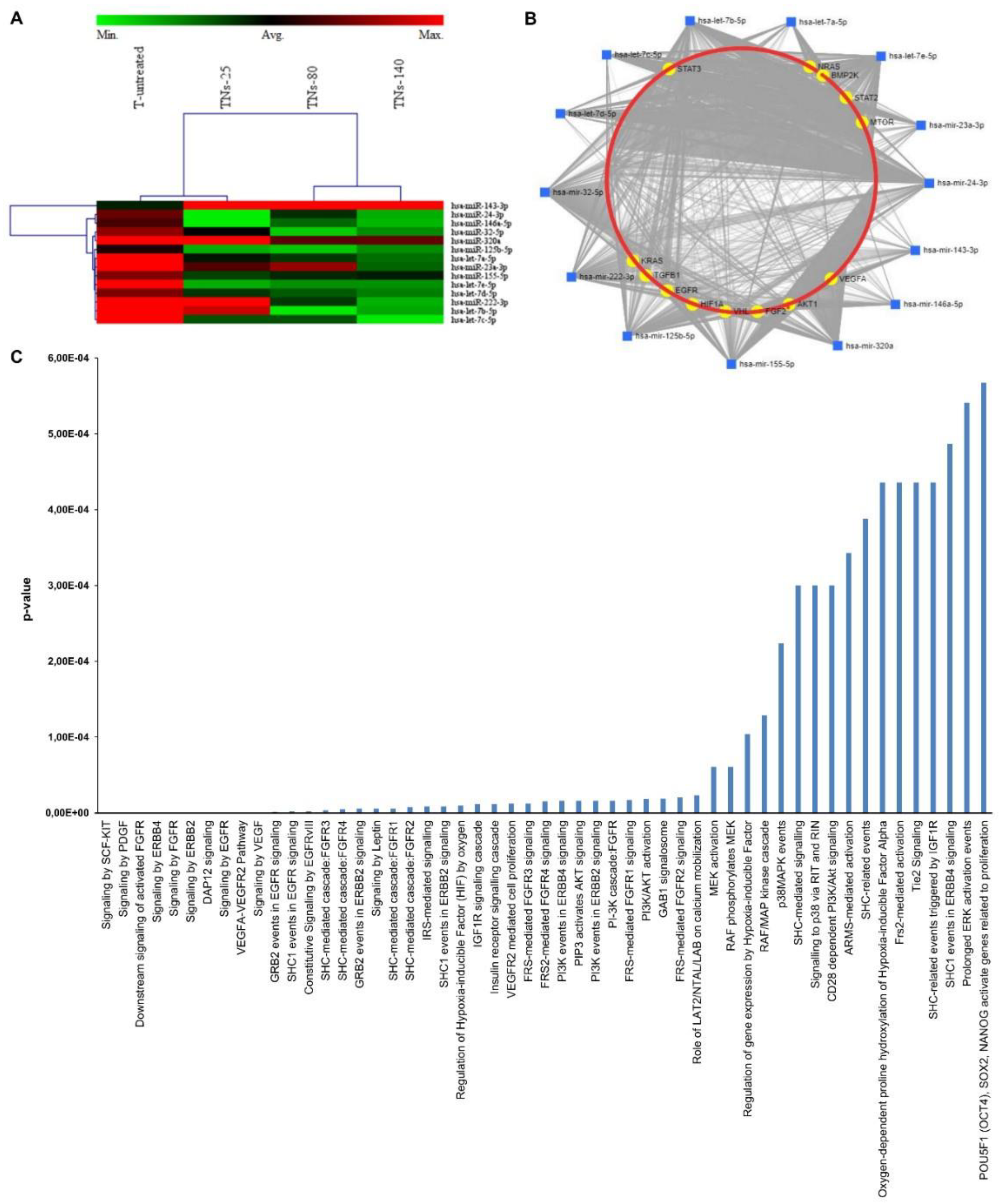
3.5. miRNA Expression Profiling of Exosomes Recovered from hMSCs Grown onto TNs
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mao, A.S.; Mooney, D.J. Regenerative medicine: Current therapies and future directions. Proc. Natl. Acad. Sci. USA 2015, 112, 14452–14459. [Google Scholar] [CrossRef] [Green Version]
- Mani, G.; Feldman, M.D.; Patel, D.; Agrawal, C.M. Coronary stents: A materials perspective. Biomaterials 2007, 28, 1689–1710. [Google Scholar] [CrossRef]
- German, M.J.; Osei-Bempong, C.; Knuth, C.A.; Deehan, D.J.; Oldershaw, R.A. Investigating the biological response of human mesenchymal stem cells to titanium surfaces. J. Orthop. Surg. Res. 2014, 9, 135. [Google Scholar] [CrossRef]
- Marrella, A.; Lee, T.Y.; Lee, D.H.; Karuthedom, S.; Syla, D.; Chawla, A.; Khademhosseini, A.; Jang, H.L. Engineering vascularized and innervated bone biomaterials for improved skeletal tissue regeneration. Mater. Today 2018, 21, 362–376. [Google Scholar] [CrossRef]
- Wu, C.; Ramaswamy, Y.; Liu, X.; Wang, G.; Zreiqat, H. Plasma-sprayed CaTiSiO5 ceramic coating on Ti-6Al-4V with excellent bonding strength, stability and cellular bioactivity. J. R. Soc. Interface 2009, 6, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Sivolella, S.; Brunello, G.; Ferroni, L.; Berengo, M.; Meneghello, R.; Savio, G.; Piattelli, A.; Gardin, C.; Zavan, B. A Novel In Vitro Technique for Assessing Dental Implant Osseointegration. Tissue Eng. Part C Methods 2016, 22, 132–141. [Google Scholar] [CrossRef]
- Gardin, C.; Ferroni, L.; Bressan, E.; Calvo-Guirado, J.L.; Degidi, M.; Piattelli, A.; Zavan, B. Adult stem cells properties in terms of commitment, aging and biological safety of grit-blasted and Acid-etched ti dental implants surfaces. Int. J. Mol. Cell. Med. 2014, 3, 225–236. [Google Scholar]
- Elsayed, H.; Brunello, G.; Gardin, C.; Ferroni, L.; Badocco, D.; Pastore, P.; Sivolella, S.; Zavan, B.; Biasetto, L. Bioactive Sphene-Based Ceramic Coatings on cpTi Substrates for Dental Implants: An In Vitro Study. Materials 2018, 11, 2234. [Google Scholar] [CrossRef] [Green Version]
- Shahryari, A.; Azari, F.; Vali, H.; Omanovic, S. The response of fibrinogen, platelets, endothelial and smooth muscle cells to an electrochemically modified SS316LS surface: Towards the enhanced biocompatibility of coronary stents. Acta Biomater. 2010, 6, 695–701. [Google Scholar] [CrossRef]
- Khudhair, D.; Bhatti, A.; Li, Y.; Hamedani, H.A.; Garmestani, H.; Hodgson, P.; Nahavandi, S. Anodization parameters influencing the morphology and electrical properties of TiO2 nanotubes for living cell interfacing and investigations. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 59, 1125–1142. [Google Scholar] [CrossRef]
- Beltrán-Partida, E.; Valdéz-Salas, B.; Moreno-Ulloa, A.; Escamilla, A.; Curiel, M.A.; Rosales-Ibáñez, R.; Villarreal, F.; Bastidas, D.M.; Bastidas, J.M. Improved in vitro angiogenic behavior on anodized titanium dioxide nanotubes. J. Nanobiotechnol. 2017, 15, 10. [Google Scholar] [CrossRef] [Green Version]
- Khrunyk, Y.Y.; Belikov, S.V.; Tsurkan, M.V.; Vyalykh, I.V.; Markaryan, A.Y.; Karabanalov, M.S.; Popov, A.A.; Wysokowski, M. Surface-Dependent Osteoblasts Response to TiO. Nanomaterials 2020, 10, 320. [Google Scholar] [CrossRef] [Green Version]
- Mohan, C.C.; Chennazhi, K.P.; Menon, D. In vitro hemocompatibility and vascular endothelial cell functionality on titania nanostructures under static and dynamic conditions for improved coronary stenting applications. Acta Biomater. 2013, 9, 9568–9577. [Google Scholar] [CrossRef]
- Souza, J.C.M.; Sordi, M.B.; Kanazawa, M.; Ravindran, S.; Henriques, B.; Silva, F.S.; Aparicio, C.; Cooper, L.F. Nano-scale modification of titanium implant surfaces to enhance osseointegration. Acta Biomater. 2019, 94, 112–131. [Google Scholar] [CrossRef]
- Gong, Z.; Hu, Y.; Gao, F.; Quan, L.; Liu, T.; Gong, T.; Pan, C. Effects of diameters and crystals of titanium dioxide nanotube arrays on blood compatibility and endothelial cell behaviors. Colloids Surf. B Biointerfaces 2019, 184, 110521. [Google Scholar] [CrossRef]
- Cao, Y.; Desai, T.A. TiO. ACS Biomater. Sci. Eng. 2020, 6, 923–932. [Google Scholar] [CrossRef]
- Junkar, I.; Kulkarni, M.; Benčina, M.; Kovač, J.; Mrak-Poljšak, K.; Lakota, K.; Sodin-Šemrl, S.; Mozetič, M.; Iglič, A. Titanium Dioxide Nanotube Arrays for Cardiovascular Stent Applications. ACS Omega 2020, 5, 7280–7289. [Google Scholar] [CrossRef] [Green Version]
- Gnecchi, M.; Zhang, Z.; Ni, A.; Dzau, V.J. Paracrine mechanisms in adult stem cell signaling and therapy. Circ. Res. 2008, 103, 1204–1219. [Google Scholar] [CrossRef]
- Tao, H.; Han, Z.; Han, Z.C.; Li, Z. Proangiogenic Features of Mesenchymal Stem Cells and Their Therapeutic Applications. Stem. Cells Int. 2016, 2016, 1314709. [Google Scholar] [CrossRef] [Green Version]
- Lamichhane, T.N.; Sokic, S.; Schardt, J.S.; Raiker, R.S.; Lin, J.W.; Jay, S.M. Emerging roles for extracellular vesicles in tissue engineering and regenerative medicine. Tissue Eng. Part B Rev. 2015, 21, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Teng, X.; Chen, L.; Chen, W.; Yang, J.; Yang, Z.; Shen, Z. Mesenchymal Stem Cell-Derived Exosomes Improve the Microenvironment of Infarcted Myocardium Contributing to Angiogenesis and Anti-Inflammation. Cell. Physiol. Biochem. 2015, 37, 2415–2424. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef]
- Wang, X.; Omar, O.; Vazirisani, F.; Thomsen, P.; Ekström, K. Mesenchymal stem cell-derived exosomes have altered microRNA profiles and induce osteogenic differentiation depending on the stage of differentiation. PLoS ONE 2018, 13, e0193059. [Google Scholar] [CrossRef]
- Bellin, G.; Gardin, C.; Ferroni, L.; Chachques, J.C.; Rogante, M.; Mitrečić, D.; Ferrari, R.; Zavan, B. Exosome in Cardiovascular Diseases: A Complex World Full of Hope. Cells 2019, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- Shabbir, A.; Cox, A.; Rodriguez-Menocal, L.; Salgado, M.; Van Badiavas, E. Mesenchymal Stem Cell Exosomes Induce Proliferation and Migration of Normal and Chronic Wound Fibroblasts, and Enhance Angiogenesis In Vitro. Stem. Cells Dev. 2015, 24, 1635–1647. [Google Scholar] [CrossRef]
- Kummer, K.M.; Taylor, E.N.; Durmas, N.G.; Tarquinio, K.M.; Ercan, B.; Webster, T.J. Effects of different sterilization techniques and varying anodized TiO2 nanotube dimensions on bacteria growth. J. Biomed. Mater. Res. B Appl. Biomater. 2013, 101, 677–688. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic. Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Ferroni, L.; Gardin, C.; Dalla Paola, L.; Campo, G.; Cimaglia, P.; Bellin, G.; Pinton, P.; Zavan, B. Characterization of Dermal Stem Cells of Diabetic Patients. Cells 2019, 8, 729. [Google Scholar] [CrossRef] [Green Version]
- Piszczek, P.; Radtke, A.; Ehlert, M.; Jędrzejewski, T.; Sznarkowska, A.; Sadowska, B.; Bartmański, M.; Erdoğan, Y.K.; Ercan, B.; Jedrzejczyk, W. Comprehensive Evaluation of the Biological Properties of Surface-Modified Titanium Alloy Implants. J. Clin. Med. 2020, 9, 342. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Park, Y.C. Contact Angle Relaxation and Long-Lasting Hydrophilicity of Sputtered Anatase TiO. Langmuir 2019, 35, 2066–2077. [Google Scholar] [CrossRef]
- Oh, S.; Brammer, K.S.; Li, Y.S.; Teng, D.; Engler, A.J.; Chien, S.; Jin, S. Stem cell fate dictated solely by altered nanotube dimension. Proc. Natl. Acad. Sci. USA 2009, 106, 2130–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowden, K.; Dias-Netipanyj, M.F.; Popat, K.C. Effects of titania nanotube surfaces on osteogenic differentiation of human adipose-derived stem cells. Nanomedicine 2019, 17, 380–390. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, S.; Chang, J.; Huan, Z.; Li, H. TiO2 Nanotubes Enhance Vascularization and Osteogenic Differentiation Through Stimulating Interactions Between Bone Marrow Stromal Cells and Endothelial Cells. J. Biomed. Nanotechnol. 2018, 14, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Hutvágner, G.; Zamore, P.D. A microRNA in a multiple-turnover RNAi enzyme complex. Science 2002, 297, 2056–2060. [Google Scholar] [CrossRef] [Green Version]
- Khraiwesh, B.; Arif, M.A.; Seumel, G.I.; Ossowski, S.; Weigel, D.; Reski, R.; Frank, W. Transcriptional control of gene expression by microRNAs. Cell 2010, 140, 111–122. [Google Scholar] [CrossRef]
- Karar, J.; Maity, A. PI3K/AKT/mTOR Pathway in Angiogenesis. Front. Mol. Neurosci. 2011, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yang, H.; Zhu, L.; Luo, Y.; Nie, L.; Li, G. Role of EGFR/ErbB2 and PI3 K/AKT/e-NOS in Lycium barbarum polysaccharides Ameliorating Endothelial Dysfunction Induced by Oxidative Stress. Am. J. Chin. Med. 2019, 47, 1523–1539. [Google Scholar] [CrossRef]
- Ganta, V.C.; Choi, M.; Kutateladze, A.; Annex, B.H. VEGF165b Modulates Endothelial VEGFR1-STAT3 Signaling Pathway and Angiogenesis in Human and Experimental Peripheral Arterial Disease. Circ. Res. 2017, 120, 282–295. [Google Scholar] [CrossRef] [Green Version]
- Hwang, C.W.; Johnston, P.V.; Gerstenblith, G.; Weiss, R.G.; Tomaselli, G.F.; Bogdan, V.E.; Panigrahi, A.; Leszczynska, A.; Xia, Z. Stem cell impregnated nanofiber stent sleeve for on-stent production and intravascular delivery of paracrine factors. Biomaterials 2015, 52, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Gittens, R.A.; Olivares-Navarrete, R.; McLachlan, T.; Cai, Y.; Hyzy, S.L.; Schneider, J.M.; Schwartz, Z.; Sandhage, K.H.; Boyan, B.D. Differential responses of osteoblast lineage cells to nanotopographically-modified, microroughened titanium-aluminum-vanadium alloy surfaces. Biomaterials 2012, 33, 8986–8994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucuzian, A.A.; Gassman, A.A.; East, A.T.; Greisler, H.P. Molecular mediators of angiogenesis. J. Burn. Care Res. 2010, 31, 158–175. [Google Scholar] [CrossRef]
- Chang, H.K.; Kim, P.H.; Kim, D.W.; Cho, H.M.; Jeong, M.J.; Kim, D.H.; Joung, Y.K.; Lim, K.S.; Kim, H.B.; Lim, H.C.; et al. Coronary stents with inducible VEGF/HGF-secreting UCB-MSCs reduced restenosis and increased re-endothelialization in a swine model. Exp. Mol. Med. 2018, 50, 114. [Google Scholar] [CrossRef] [Green Version]
- Kaga, T.; Kawano, H.; Sakaguchi, M.; Nakazawa, T.; Taniyama, Y.; Morishita, R. Hepatocyte growth factor stimulated angiogenesis without inflammation: Differential actions between hepatocyte growth factor, vascular endothelial growth factor and basic fibroblast growth factor. Vascul. Pharmacol. 2012, 57, 3–9. [Google Scholar] [CrossRef]
- Chang, H.K.; Kim, P.H.; Cho, H.M.; Yum, S.Y.; Choi, Y.J.; Son, Y.; Lee, D.; Kang, I.; Kang, K.S.; Jang, G.; et al. Inducible HGF-secreting Human Umbilical Cord Blood-derived MSCs Produced via TALEN-mediated Genome Editing Promoted Angiogenesis. Mol. Ther. 2016, 24, 1644–1654. [Google Scholar] [CrossRef] [Green Version]
- Valente, S.; Ciavarella, C.; Pasanisi, E.; Ricci, F.; Stella, A.; Pasquinelli, G. Hepatocyte Growth Factor Effects on Mesenchymal Stem Cells Derived from Human Arteries: A Novel Strategy to Accelerate Vascular Ulcer Wound Healing. Stem. Cells Int. 2016, 2016, 3232859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mijiritsky, E.; Gardin, C.; Ferroni, L.; Lacza, Z.; Zavan, B. Albumin-impregnated bone granules modulate the interactions between mesenchymal stem cells and monocytes under in vitro inflammatory conditions. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 110, 110678. [Google Scholar] [CrossRef] [PubMed]
- Rammal, H.; Entz, L.; Dubus, M.; Moniot, A.; Bercu, N.B.; Sergheraert, J.; Gangloff, S.C.; Mauprivez, C.; Kerdjoudj, H. Osteoinductive Material to Fine-Tune Paracrine Crosstalk of Mesenchymal Stem Cells With Endothelial Cells and Osteoblasts. Front. Bioeng. Biotechnol. 2019, 7, 256. [Google Scholar] [CrossRef] [PubMed]
- Abnaof, K.; Mallela, N.; Walenda, G.; Meurer, S.K.; Seré, K.; Lin, Q.; Smeets, B.; Hoffmann, K.; Wagner, W.; Zenke, M.; et al. TGF-β stimulation in human and murine cells reveals commonly affected biological processes and pathways at transcription level. BMC Syst. Biol. 2014, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Finkenzeller, G.; Hager, S.; Stark, G.B. Effects of bone morphogenetic protein 2 on human umbilical vein endothelial cells. Microvasc. Res. 2012, 84, 81–85. [Google Scholar] [CrossRef]
- Suzuki, Y.; Montagne, K.; Nishihara, A.; Watabe, T.; Miyazono, K. BMPs promote proliferation and migration of endothelial cells via stimulation of VEGF-A/VEGFR2 and angiopoietin-1/Tie2 signalling. J. Biochem. 2008, 143, 199–206. [Google Scholar] [CrossRef]
- Alcayaga-Miranda, F.; Varas-Godoy, M.; Khoury, M. Harnessing the Angiogenic Potential of Stem Cell-Derived Exosomes for Vascular Regeneration. Stem. Cells Int. 2016, 2016, 3409169. [Google Scholar] [CrossRef] [Green Version]
- Brammer, K.S.; Oh, S.; Gallagher, J.O.; Jin, S. Enhanced cellular mobility guided by TiO2 nanotube surfaces. Nano Lett. 2008, 8, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Eltgroth, M.L.; LaTempa, T.J.; Grimes, C.A.; Desai, T.A. The effect of TiO2 nanotubes on endothelial function and smooth muscle proliferation. Biomaterials 2009, 30, 1268–1272. [Google Scholar] [CrossRef]
- Park, J.; Bauer, S.; Schmuki, P.; von der Mark, K. Narrow window in nanoscale dependent activation of endothelial cell growth and differentiation on TiO2 nanotube surfaces. Nano Lett. 2009, 9, 3157–3164. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Olson, E.N. AngiomiRs—Key regulators of angiogenesis. Curr. Opin. Genet. Dev. 2009, 19, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Salinas-Vera, Y.M.; Marchat, L.A.; Gallardo-Rincón, D.; Ruiz-García, E.; Astudillo-De La Vega, H.; Echavarría-Zepeda, R.; López-Camarillo, C. AngiomiRs: MicroRNAs driving angiogenesis in cancer (Review). Int. J. Mol. Med. 2019, 43, 657–670. [Google Scholar] [CrossRef] [Green Version]
- Varghese, E.; Liskova, A.; Kubatka, P.; Mathews Samuel, S.; Büsselberg, D. Anti-Angiogenic Effects of Phytochemicals on miRNA Regulating Breast Cancer Progression. Biomolecules 2020, 10, 191. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Huang, L.F.; Zhao, L.; Zeng, Z.; Wang, X.; Cao, D.; Yang, L.; Ye, Z.; Chen, X.; Liu, B.; et al. Microvesicles (MIVs) secreted from adipose-derived stem cells (ADSCs) contain multiple microRNAs and promote the migration and invasion of endothelial cells. Genes Dis. 2020, 7, 225–234. [Google Scholar] [CrossRef]
- Thai, S.F.; Wallace, K.A.; Jones, C.P.; Ren, H.; Prasad, R.Y.; Ward, W.O.; Kohan, M.J.; Blackman, C.F. Signaling Pathways and MicroRNA Changes in Nano-TiO2 Treated Human Lung Epithelial (BEAS-2B) Cells. J. Nanosci. Nanotechnol. 2015, 15, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, J.; Hu, X.; Wang, Z.; Wu, S.; Yi, Y. Extracellular vesicles derived from human adipose-derived stem cells promote the exogenous angiogenesis of fat grafts via the let-7/AGO1/VEGF signalling pathway. Sci. Rep. 2020, 10, 5313. [Google Scholar] [CrossRef] [Green Version]
- Bao, M.H.; Feng, X.; Zhang, Y.W.; Lou, X.Y.; Cheng, Y.; Zhou, H.H. Let-7 in cardiovascular diseases, heart development and cardiovascular differentiation from stem cells. Int. J. Mol. Sci. 2013, 14, 23086–23102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Zhang, H.; Ding, W.; Fan, Z.; Ji, B.; Ding, C.; Ji, F.; Tang, H. miR-143 promotes angiogenesis and osteoblast differentiation by targeting HDAC7. Cell. Death Dis. 2020, 11, 179. [Google Scholar] [CrossRef] [Green Version]
- Poliseno, L.; Tuccoli, A.; Mariani, L.; Evangelista, M.; Citti, L.; Woods, K.; Mercatanti, A.; Hammond, S.; Rainaldi, G. MicroRNAs modulate the angiogenic properties of HUVECs. Blood 2006, 108, 3068–3071. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Lv, Q.; Ye, W.; Wong, C.K.; Cai, G.; Gu, D.; Ji, Y.; Zhao, C.; Wang, J.; Yang, B.B.; et al. MiRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS ONE 2006, 1, e116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.C.; Ding, Z.Y.; Wang, H.Q.; Ning, L.P.; Wang, C. Effect of microRNA-155 on angiogenesis after cerebral infarction of rats through AT1R/VEGFR2 pathway. Asian Pac. J. Trop. Med. 2015, 8, 829–835. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Pan, J.; Wang, H.; Ma, Z.; Yin, J.; Yuan, F.; Yuan, Q.; Zhou, L.; Liu, X.; Zhang, Y.; et al. von Willebrand factor rescued by miR-24 inhibition facilitates the proliferation and migration of osteosarcoma cells in vitro. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Yan, T.; Gao, Y. Silence of miR-32-5p promotes endothelial cell viability by targeting KLF2 and serves as a diagnostic biomarker of acute myocardial infarction. Diagn. Pathol. 2020, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Zhang, P.; Liang, P.; Huang, X. The expression of miR-125b regulates angiogenesis during the recovery of heat-denatured HUVECs. Burns 2015, 41, 803–811. [Google Scholar] [CrossRef]
- Cheng, H.S.; Sivachandran, N.; Lau, A.; Boudreau, E.; Zhao, J.L.; Baltimore, D.; Delgado-Olguin, P.; Cybulsky, M.I.; Fish, J.E. MicroRNA-146 represses endothelial activation by inhibiting pro-inflammatory pathways. EMBO Mol. Med. 2013, 5, 1017–1034. [Google Scholar] [CrossRef]
- Fang, Y.; Chen, S.; Liu, Z.; Ai, W.; He, X.; Wang, L.; Xie, P.; Jiang, B.; Fang, H. Endothelial stem cells attenuate cardiac apoptosis via downregulating cardiac microRNA-146a in a rat model of coronary heart disease. Exp. Ther. Med. 2018, 16, 4246–4252. [Google Scholar] [CrossRef]
- Liu, R.H.; Meng, Q.; Shi, Y.P.; Xu, H.S. Regulatory role of microRNA-320a in the proliferation, migration, invasion, and apoptosis of trophoblasts and endothelial cells by targeting estrogen-related receptor γ. J. Cell. Physiol. 2018, 234, 682–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence FOR (5′-3′) | Sequence REV (5′-3′) |
|---|---|---|
| bFGF 1 | TCCATCCTTTCTCCCTCGTTTC | TCAGTAGATGTTTCCCTCCAATGTTTC |
| BMP2 2 | CCACTAATCATGCCATTGTTCAGAC | CTGTACTAGCGACACCCACAA |
| HGF 3 | CTTCAATAGCATGTCAAGTGGAGTG | GCTGCGTCCTTTACCAATGATG |
| TFRC 4 | TGTTTGTCATAGGGCAGTTGGAA | ACACCCGAACCAGGAATCTC |
| TGFB1 5 | TGACAGCAGGGATAACACACT | CCGTGGAGCTGAAGCA |
| VEGFA 6 | GGACAGAAAGACAGATCACAGGTAC | GCAGGTGAGAGTAAGCGAAGG |
| Gene Name | Sequence FOR (5′-3′) | Sequence REV (5′-3′) |
|---|---|---|
| CD31 1 | TGAGACCAGCCTGATGAAACC | CGATCTCCGCTCACTACAACC |
| TFRC 2 | TGTTTGTCATAGGGCAGTTGGAA | ACACCCGAACCAGGAATCTC |
| VEGFR2 3 | GGAGGAGGAGGAAGTATGTGACC | AACCATACCACTGTCCGTCTG |
| VWF 4 | ACGTATGGTCTGTGTGGGATC | GACAAGACACTGCTCCTCCA |
| Sample | Contact Angle (deg °) | Rms (Nanophase Roughness) (nm) | Rms (Micronphase Roughness) (nm) | Surface Area (μm2) |
|---|---|---|---|---|
| T-untreated | 85.3 ± 7.1 | 5.5 ± 1 | 160 ± 4 | 0.95 ± 0.07 |
| TNs-25 | 15.8 ± 2.7 * | 17.6 ± 3 * | 145 ± 3 | 1.43 ± 0.08 * |
| TNs-80 | 19.3 ± 3.3 * | 28.9 ± 2 * | 113 ± 2 | 1.87 ± 0.11 s * |
| TNs-140 | 23.5 ± 5.5 * | 34.5 ± 3 ** | 97 ± 4 | 2.29 ± 0.09 ** |
| miRNA | TNs-25 | TNs-80 | TNs-140 | |||
|---|---|---|---|---|---|---|
| Fold-Change | p-Value | Fold-Change | p-Value | Fold-Change | p-Value | |
| hsa-miR-142-5p | 0.88 | 0.464 | 0.85 | 0.386 | 1.48 | 0.112 |
| hsa-miR-9-5p | 0.58 | 0.667 | 1.01 | 0.855 | 1.14 | 0.641 |
| hsa-miR-150-5p | 1.11 | 0.732 | 1.84 | 0.885 | 1.74 | 0.616 |
| hsa-miR-27b-3p | 1.10 | 0.770 | 0.43 | 0.743 | 0.44 | 0.462 |
| hsa-miR-101-3p | 1.21 | 0.850 | 1.97 | 0.109 | 1.32 | 0.658 |
| hsa-let-7d-5p | 0.40 | 0.002 | 0.33 | 0.016 | 0.25 | 0.000 |
| hsa-miR-103a-3p | 0.20 | 0.995 | 0.39 | 0.959 | 0.30 | 0.549 |
| hsa-miR-16-5p | 0.98 | 0.744 | 0.43 | 0.517 | 0.86 | 0.553 |
| hsa-miR-26a-5p | 0.37 | 0.375 | 0.40 | 0.523 | 0.34 | 0.231 |
| hsa-miR-32-5p | 0.47 | 0.005 | 0.09 | 0.022 | 0.21 | 0.002 |
| hsa-miR-26b-5p | 0.56 | 0.672 | 0.25 | 0.774 | 0.27 | 0.486 |
| hsa-let-7g-5p | 0.57 | 0.720 | 1.40 | 0.862 | 0.61 | 0.558 |
| hsa-miR-30c-5p | 0.36 | 0.092 | 0.24 | 0.180 | 0.24 | 0.887 |
| hsa-miR-96-5p | 0.96 | 0.725 | 0.65 | 0.678 | 1.20 | 0.912 |
| hsa-miR-185-5p | 0.41 | 0.038 | 0.36 | 0.030 | 0.44 | 0.064 |
| hsa-miR-142-3p | 0.17 | 0.592 | 0.36 | 0.737 | 0.72 | 0.526 |
| hsa-miR-24-3p | 0.04 | 0.011 | 0.45 | 0.020 | 0.17 | 0.000 |
| hsa-miR-155-5p | 0.37 | 0.003 | 0.42 | 0.011 | 0.44 | 0.002 |
| hsa-miR-146a-5p | 0.04 | 0.021 | 0.38 | 0.048 | 0.18 | 0.002 |
| hsa-miR-425-5p | 0.57 | 0.720 | 0.67 | 0.843 | 0.61 | 0.558 |
| hsa-miR-181b-5p | 2.43 | 0.791 | 2.43 | 0.752 | 3.60 | 0.522 |
| hsa-miR-302b-3p | 0.38 | 0.681 | 0.50 | 0.811 | 0.38 | 0.522 |
| hsa-miR-30b-5p | 0.60 | 0.385 | 0.48 | 0.473 | 0.20 | 0.145 |
| hsa-miR-21-5p | 1.26 | 0.880 | 0.15 | 0.651 | 0.53 | 0.433 |
| hsa-miR-30e-5p | 0.89 | 0.497 | 0.83 | 0.346 | 0.99 | 0.797 |
| hsa-miR-200c-3p | 0.97 | 0.728 | 0.58 | 0.685 | 0.63 | 0.435 |
| hsa-miR-15b-5p | 0.47 | 0.010 | 0.85 | 0.097 | 0.96 | 0.545 |
| hsa-miR-223-3p | 0.73 | 0.668 | 1.72 | 0.897 | 0.73 | 0.526 |
| hsa-miR-194-5p | 0.70 | 0.172 | 0.43 | 0.042 | 0.60 | 0.099 |
| hsa-miR-210-3p | 0.55 | 0.393 | 0.75 | 0.638 | 0.84 | 0.526 |
| hsa-miR-15a-5p | 0.57 | 0.720 | 0.67 | 0.843 | 0.40 | 0.558 |
| hsa-miR-181a-5p | 0.06 | 0.094 | 0.27 | 0.198 | 0.04 | 0.032 |
| hsa-miR-125b-5p | 0.25 | 0.043 | 0.17 | 0.017 | 0.36 | 0.028 |
| hsa-miR-99a-5p | 0.11 | 0.019 | 0.14 | 0.027 | 1.02 | 0.898 |
| hsa-miR-28-5p | 0.57 | 0.720 | 0.67 | 0.843 | 1.62 | 0.608 |
| hsa-miR-320a | 0.39 | 0.035 | 0.02 | 0.008 | 0.03 | 0.008 |
| hsa-miR-125a-5p | 2.43 | 0.012 | 0.08 | 0.290 | 2.09 | 0.148 |
| hsa-miR-29b-3p | 6.37 | 0.803 | 3.98 | 0.983 | 0.73 | 0.564 |
| hsa-miR-29a-3p | 1.02 | 0.917 | 0.39 | 0.050 | 0.48 | 0.058 |
| hsa-miR-141-3p | 0.53 | 0.298 | 0.72 | 0.546 | 0.54 | 0.195 |
| hsa-miR-19a-3p | 0.83 | 0.718 | 1.38 | 0.895 | 1.36 | 0.647 |
| hsa-miR-18a-5p | 0.57 | 0.720 | 0.67 | 0.843 | 1.61 | 0.608 |
| hsa-miR-374a-5p | 0.57 | 0.720 | 1.20 | 0.855 | 0.75 | 0.565 |
| hsa-miR-423-5p | 0.77 | 0.251 | 0.07 | 0.011 | 0.26 | 0.020 |
| hsa-let-7a-5p | 0.14 | 0.012 | 0.12 | 0.012 | 0.09 | 0.010 |
| hsa-miR-124-3p | 1.32 | 0.262 | 1.35 | 0.220 | 1.24 | 0.412 |
| hsa-miR-92a-3p | 0.34 | 0.010 | 0.45 | 0.049 | 0.73 | 0.064 |
| hsa-miR-23a-3p | 0.15 | 0.013 | 0.18 | 0.014 | 0.05 | 0.009 |
| hsa-miR-25-3p | 0.89 | 0.476 | 1.29 | 0.322 | 1.16 | 0.615 |
| hsa-let-7e-5p | 0.08 | 0.013 | 0.11 | 0.017 | 0.11 | 0.011 |
| hsa-miR-376c-3p | 0.90 | 0.483 | 0.09 | 0.025 | 0.62 | 0.085 |
| hsa-miR-126-3p | 0.71 | 0.084 | 1.21 | 0.211 | 0.94 | 0.618 |
| hsa-miR-144-3p | 0.30 | 0.526 | 0.49 | 0.697 | 0.65 | 0.469 |
| hsa-miR-424-5p | 4.59 | 0.070 | 5.41 | 0.035 | 3.19 | 0.273 |
| hsa-miR-30a-5p | 1.10 | 0.836 | 0.63 | 0.115 | 0.99 | 0.792 |
| hsa-miR-23b-3p | 0.24 | 0.014 | 0.20 | 0.142 | 0.06 | 0.003 |
| hsa-miR-151a-5p | 0.76 | 0.657 | 0.16 | 0.670 | 0.34 | 0.402 |
| hsa-miR-195-5p | 0.34 | 0.206 | 1.04 | 0.940 | 0.68 | 0.268 |
| hsa-miR-143-3p | 4.85 | 0.001 | 4.49 | 0.001 | 16.78 | 0.000 |
| hsa-miR-30d-5p | 0.81 | 0.210 | 0.33 | 0.019 | 0.39 | 0.011 |
| hsa-miR-191-5p | 0.10 | 0.366 | 0.49 | 0.585 | 0.06 | 0.222 |
| hsa-let-7i-5p | 1.45 | 0.131 | 1.11 | 0.798 | 1.46 | 0.123 |
| hsa-miR-302a-3p | 0.87 | 0.352 | 0.38 | 0.135 | 1.33 | 0.092 |
| hsa-miR-222-3p | 0.20 | 0.016 | 0.03 | 0.008 | 0.01 | 0.008 |
| hsa-let-7b-5p | 0.19 | 0.015 | 0.01 | 0.009 | 0.03 | 0.008 |
| hsa-miR-19b-3p | 0.77 | 0.147 | 0.78 | 0.158 | 1.39 | 0.058 |
| hsa-miR-17-5p | 1.20 | 0.778 | 2.27 | 0.846 | 2.03 | 0.964 |
| hsa-miR-93-5p | 1.61 | 0.078 | 1.87 | 0.064 | 2.72 | 0.000 |
| hsa-miR-186-5p | 0.58 | 0.242 | 0.96 | 0.793 | 1.35 | 0.373 |
| hsa-miR-196b-5p | 0.55 | 0.545 | 0.68 | 0.718 | 0.75 | 0.491 |
| hsa-miR-27a-3p | 1.72 | 0.378 | 1.03 | 0.388 | 0.20 | 0.244 |
| hsa-miR-22-3p | 1.80 | 0.123 | 0.98 | 0.751 | 0.76 | 0.244 |
| hsa-miR-130a-3p | 0.58 | 0.211 | 0.74 | 0.458 | 0.58 | 0.188 |
| hsa-let-7c-5p | 0.22 | 0.019 | 0.22 | 0.025 | 0.03 | 0.011 |
| hsa-miR-29c-3p | 0.32 | 0.430 | 0.91 | 0.801 | 0.98 | 0.683 |
| hsa-miR-140-3p | 0.57 | 0.084 | 0.65 | 0.127 | 0.87 | 0.445 |
| hsa-miR-128-3p | 0.15 | 0.565 | 0.17 | 0.699 | 0.11 | 0.402 |
| hsa-let-7f-5p | 0.76 | 0.716 | 0.58 | 0.840 | 1.14 | 0.588 |
| hsa-miR-122-5p | 8.10 | 0.583 | 4.20 | 0.965 | 1.10 | 0.580 |
| hsa-miR-20a-5p | 1.04 | 0.818 | 0.82 | 0.701 | 1.87 | 0.219 |
| hsa-miR-106b-5p | 2.61 | 0.540 | 2.26 | 0.690 | 1.51 | 0.855 |
| hsa-miR-7-5p | 0.57 | 0.720 | 7.99 | 0.582 | 2.29 | 0.656 |
| hsa-miR-100-5p | 0.27 | 0.021 | 0.27 | 0.026 | 0.66 | 0.136 |
| hsa-miR-302c-3p | 0.69 | 0.419 | 0.97 | 0.815 | 0.53 | 0.215 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gardin, C.; Ferroni, L.; Erdoğan, Y.K.; Zanotti, F.; De Francesco, F.; Trentini, M.; Brunello, G.; Ercan, B.; Zavan, B. Nanostructured Modifications of Titanium Surfaces Improve Vascular Regenerative Properties of Exosomes Derived from Mesenchymal Stem Cells: Preliminary In Vitro Results. Nanomaterials 2021, 11, 3452. https://doi.org/10.3390/nano11123452
Gardin C, Ferroni L, Erdoğan YK, Zanotti F, De Francesco F, Trentini M, Brunello G, Ercan B, Zavan B. Nanostructured Modifications of Titanium Surfaces Improve Vascular Regenerative Properties of Exosomes Derived from Mesenchymal Stem Cells: Preliminary In Vitro Results. Nanomaterials. 2021; 11(12):3452. https://doi.org/10.3390/nano11123452
Chicago/Turabian StyleGardin, Chiara, Letizia Ferroni, Yaşar Kemal Erdoğan, Federica Zanotti, Francesco De Francesco, Martina Trentini, Giulia Brunello, Batur Ercan, and Barbara Zavan. 2021. "Nanostructured Modifications of Titanium Surfaces Improve Vascular Regenerative Properties of Exosomes Derived from Mesenchymal Stem Cells: Preliminary In Vitro Results" Nanomaterials 11, no. 12: 3452. https://doi.org/10.3390/nano11123452