Prunus spinosa Extract Loaded in Biomimetic Nanoparticles Evokes In Vitro Anti-Inflammatory and Wound Healing Activities

 , , , , , ,
, , , , , ,  , and
, and 
Abstract
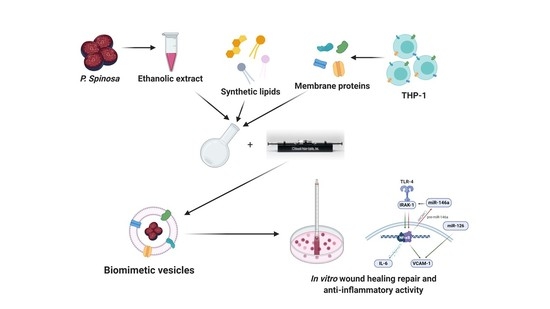
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Fruits Collection and P. spinosa Extract Preparation
2.3. Formulation of Biomimetic Nanovesicles
2.4. Physicochemical Characterization of Biomimetic Nanovesicles
2.5. Cell Culture
2.6. In Vitro Wound Healing Assay
2.7. RT–qPCR Analysis
2.8. Statistical Analysis
3. Results
3.1. Physicochemical Characterization of Empty and Extract-Loaded Vesicles
3.2. Wound Healing Repair Activity of PSF-Loaded Vesicles
3.3. Anti-Inflammatory Activity of PSF-Loaded Vesicles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound healing: A cellular perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef] [PubMed]
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in Wound Repair: Molecular and Cellular Mechanisms. J. Invest. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, N.; Wong, S.; Mohamed, I.; Mohamed, N.; Chin, K.-Y.; Ima-Nirwana, S.; Shuid, A. Wound Healing Properties of Selected Natural Products. Int. J. Environ. Res. Public Health 2018, 15, 2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraternale, D.; Giamperi, L.; Bucchini, A.; Sestili, P.; Paolillo, M.; Ricci, D. Prunus spinosa fresh fruit juice: Antioxidant activity in cell-free and cellular systems. Nat. Prod. Commun. 2009, 4, 1665–1670. [Google Scholar] [CrossRef] [Green Version]
- Pinacho, R.; Cavero, R.Y.; Astiasarán, I.; Ansorena, D.; Calvo, M.I. Phenolic compounds of blackthorn (Prunus spinosa L.) and influence of in vitro digestion on their antioxidant capacity. J. Funct. Foods 2015, 19, 49–62. [Google Scholar] [CrossRef]
- Sabatini, L.; Fraternale, D.; Di Giacomo, B.; Mari, M.; Albertini, M.C.; Gordillo, B.; Rocchi, M.B.L.; Sisti, D.; Coppari, S.; Semprucci, F.; et al. Chemical composition, antioxidant, antimicrobial and anti-inflammatory activity of Prunus spinosa L. fruit ethanol extract. J. Funct. Foods 2020, 67, 103885. [Google Scholar] [CrossRef]
- Blesso, C.N. Dietary Anthocyanins and Human Health. Nutrients 2019, 11, 2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Peng, K.J.; Wang, Q.L.; Gu, Z.Y.; Lu, Y.Q.; Zhao, J.; Xu, F.; Liu, Y.L.; Tang, Y.; Deng, F.M.; et al. Effect of pomegranate peel polyphenol gel on cutaneous wound healing in alloxan-induced diabetic rats. Chin. Med. J. 2013, 126, 1700–1706. [Google Scholar] [CrossRef]
- Georgescu, M.; Marinas, O.; Popa, M.; Stan, T.; Lazar, V.; Bertesteanu, S.V.; Chifiriuc, M.C. Natural Compounds for Wound Healing. In Worldwide Wound Healing-Innovation in Natural and Conventional Methods; InTech: London, UK, 2016. [Google Scholar]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef]
- Wolfram, J.; Suri, K.; Huang, Y.; Molinaro, R.; Borsoi, C.; Scott, B.; Boom, K.; Paolino, D.; Fresta, M.; Wang, J.; et al. Evaluation of anticancer activity of celastrol liposomes in prostate cancer cells. J. Microencapsul. 2014, 31, 501–507. [Google Scholar] [CrossRef] [Green Version]
- Kirui, D.K.; Celia, C.; Molinaro, R.; Bansal, S.S.; Cosco, D.; Fresta, M.; Shen, H.; Ferrari, M. Mild Hyperthermia Enhances Transport of Liposomal Gemcitabine and Improves In Vivo Therapeutic Response. Adv. Healthc. Mater. 2015, 4, 1092–1103. [Google Scholar] [CrossRef] [Green Version]
- Cosco, D.; Molinaro, R.; Morittu, V.M.; Cilurzo, F.; Costa, N.; Fresta, M. Anticancer activity of 9-cis-retinoic acid encapsulated in PEG-coated PLGA-nanoparticles. J. Drug Deliv. Sci. Technol. 2011, 21, 395–400. [Google Scholar] [CrossRef]
- Cosco, D.; Federico, C.; Maiuolo, J.; Bulotta, S.; Molinaro, R.; Paolino, D.; Tassone, P.; Fresta, M. Physicochemical features and transfection properties of chitosan/poloxamer 188/poly(D,L-lactide-co-glycolide) nanoplexes. Int. J. Nanomedicine 2014, 9, 2359–2372. [Google Scholar] [CrossRef] [Green Version]
- Paolino, D.; Cosco, D.; Molinaro, R.; Celia, C.; Fresta, M. Supramolecular devices to improve the treatment of brain diseases. Drug Discov. Today 2011, 16, 311–324. [Google Scholar] [CrossRef]
- Carter, A.M.; Tan, C.; Pozo, K.; Telange, R.; Molinaro, R.; Guo, A.; De Rosa, E.; Martinez, J.O.; Zhang, S.; Kumar, N.; et al. Phosphoprotein-based biomarkers as predictors for cancer therapy. Proc. Natl. Acad. Sci. USA 2020, 117, 18401–18411. [Google Scholar] [CrossRef]
- Mohammadi, M.R.; Corbo, C.; Molinaro, R.; Lakey, J.R.T. Biohybrid Nanoparticles to Negotiate with Biological Barriers. Small 2019, 15, e1902333. [Google Scholar] [CrossRef]
- Parodi, A.; Molinaro, R.; Sushnitha, M.; Evangelopoulos, M.; Martinez, J.O.; Arrighetti, N.; Corbo, C.; Tasciotti, E. Bio-inspired engineering of cell- and virus-like nanoparticles for drug delivery. Biomaterials 2017, 147, 155–168. [Google Scholar] [CrossRef]
- Corbo, C.; Cromer, W.E.; Molinaro, R.; Toledano Furman, N.E.; Hartman, K.A.; De Rosa, E.; Boada, C.; Wang, X.; Zawieja, D.C.; Agostini, M.; et al. Engineered biomimetic nanovesicles show intrinsic anti-inflammatory properties for the treatment of inflammatory bowel diseases. Nanoscale 2017, 9, 14581–14591. [Google Scholar] [CrossRef]
- Molinaro, R.; Pastò, A.; Corbo, C.; Taraballi, F.; Giordano, F.; Martinez, J.O.; Zhao, P.; Wang, X.; Zinger, A.; Boada, C.; et al. Macrophage-derived nanovesicles exert intrinsic anti-inflammatory properties and prolong survival in sepsis through a direct interaction with macrophages. Nanoscale 2019, 11, 13576–13586. [Google Scholar] [CrossRef]
- Molinaro, R.; Corbo, C.; Martinez, J.O.; Taraballi, F.; Evangelopoulos, M.; Minardi, S.; Yazdi, I.K.; Zhao, P.; De Rosa, E.; Sherman, M.B.; et al. Biomimetic proteolipid vesicles for targeting inflamed tissues. Nat. Mater. 2016, 15, 1037–1046. [Google Scholar] [CrossRef]
- Hu, C.M.J.; Fang, R.H.; Zhang, L. Erythrocyte-inspired delivery systems. Adv. Healthc. Mater. 2012, 1, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Gao, J.; Fang, R.H.; Luk, B.T.; Kroll, A.V.; Dehaini, D.; Zhou, J.; Kim, H.W.; Gao, W.; Lu, W.; et al. Nanoparticles camouflaged in platelet membrane coating as an antibody decoy for the treatment of immune thrombocytopenia. Biomaterials 2016, 111, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.-Y.; Cheng, H.; Xie, B.-R.; Qiu, W.-X.; Zeng, J.-Y.; Li, C.-X.; Wan, S.-S.; Zhang, L.; Liu, W.-L.; Zhang, X.-Z. Cancer Cell Membrane Camouflaged Cascade Bioreactor for Cancer Targeted Starvation and Photodynamic Therapy. ACS Nano 2017, 11, 7006–7018. [Google Scholar] [CrossRef] [PubMed]
- Molinaro, R.; Evangelopoulos, M.; Hoffman, J.R.; Corbo, C.; Taraballi, F.; Martinez, J.O.; Hartman, K.A.; Cosco, D.; Costa, G.; Romeo, I.; et al. Design and Development of Biomimetic Nanovesicles Using a Microfluidic Approach. Adv. Mater. 2018, 30, 1702749. [Google Scholar] [CrossRef]
- Molinaro, R.; Martinez, J.O.; Zinger, A.; De Vita, A.; Storci, G.; Arrighetti, N.; De Rosa, E.; Hartman, K.A.; Basu, N.; Taghipour, N.; et al. Leukocyte-mimicking nanovesicles for effective doxorubicin delivery to treat breast cancer and melanoma. Biomater. Sci. 2020, 8, 333–341. [Google Scholar] [CrossRef]
- Boada, C.; Zinger, A.; Tsao, C.; Zhao, P.; Martinez, J.O.; Hartman, K.; Naoi, T.; Sukhoveshin, R.; Sushnitha, M.; Molinaro, R.; et al. Rapamycin-Loaded Biomimetic Nanoparticles Reverse Vascular Inflammation. Circ. Res. 2020, 126, 25–37. [Google Scholar] [CrossRef]
- Martinez, J.O.; Molinaro, R.; Hartman, K.A.; Boada, C.; Sukhovershin, R.; De Rosa, E.; Kirui, D.; Zhang, S.; Evangelopoulos, M.; Carter, A.M.; et al. Biomimetic nanoparticles with enhanced affinity towards activated endothelium as versatile tools for theranostic drug delivery. Theranostics 2018, 8, 1131–1145. [Google Scholar] [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef]
- Popović, B.M.; Blagojević, B.; Ždero Pavlović, R.; Mićić, N.; Bijelić, S.; Bogdanović, B.; Mišan, A.; Duarte, C.M.M.; Serra, A.T. Comparison between polyphenol profile and bioactive response in blackthorn (Prunus spinosa L.) genotypes from north Serbia-from raw data to PCA analysis. Food Chem. 2020, 302. [Google Scholar] [CrossRef]
- Gironés-Vilaplana, A.; Villaño, D.; Moreno, D.A.; García-Viguera, C. New isotonic drinks with antioxidant and biological capacities from berries (maqui, açaí and blackthorn) and lemon juice. Int. J. Food Sci. Nutr. 2013, 64, 897–906. [Google Scholar] [CrossRef]
- Guimarães, R.; Barros, L.; Dueñas, M.; Carvalho, A.M.; Queiroz, M.J.R.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Characterisation of phenolic compounds in wild fruits from Northeastern Portugal. Food Chem. 2013, 141, 3721–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biesalski, H.K. Polyphenols and inflammation: Basic interactions. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 724–728. [Google Scholar] [CrossRef]
- Natsume, M. Polyphenols: Inflammation. Curr. Pharm. Des. 2018, 24, 191–202. [Google Scholar] [CrossRef]
- Albertini, M.C.; Fraternale, D.; Semprucci, F.; Cecchini, S.; Colomba, M.; Rocchi, M.B.L.; Sisti, D.; Di Giacomo, B.; Mari, M.; Sabatini, L.; et al. Bioeffects of Prunus spinosa L. Fruit ethanol extract on reproduction and phenotypic plasticity of Trichoplax adhaerens Schulze, 1883 (Placozoa). PeerJ 2019, 7. [Google Scholar] [CrossRef]
- Corbo, C.; Molinaro, R.; Taraballi, F.; Toledano Furman, N.E.; Hartman, K.A.; Sherman, M.B.; De Rosa, E.; Kirui, D.K.; Salvatore, F.; Tasciotti, E. Unveiling the in Vivo Protein Corona of Circulating Leukocyte-like Carriers. ACS Nano 2017, 11, 3262–3273. [Google Scholar] [CrossRef] [PubMed]
- Millán, C.G.; Marinero, M.L.S.; Castañeda, A.Z.; Lanao, J.M. Drug, enzyme and peptide delivery using erythrocytes as carriers. J. Control. Release 2004, 95, 27–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Dipietro, L.A. Toll-like receptor function in acute wounds. Adv. Wound Care 2017, 6, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Liang, J.; Xia, X.; Chen, X. miRNA-126 enhances viability, colony formation, and migration of keratinocytes HaCaT cells by regulating PI3 K/AKT signaling pathway. Cell Biol. Int. 2019, 43, 182–191. [Google Scholar] [CrossRef]
- Fei, J.; Ling, Y.M.; Zeng, M.J.; Zhang, K.W. Shixiang plaster, a traditional Chinese medicine, promotes healing in a rat model of diabetic ulcer through the receptor for advanced glycation end products (RAGE)/Nuclear Factor kappa B (NF-kB) and Vascular Endothelial Growth Factor (VEGF)/Vascular Cell Adhesion Molecule-1 (VCAM-1)/Endothelial Nitric Oxide Synthase (eNOS) signaling pathways. Med. Sci. Monit. 2019, 25, 9446–9457. [Google Scholar] [CrossRef]
- Shahzad Aslam, M.; Syarhabil Ahmad, M.; Riaz, H.; Atif Raza, S.; Hussain, S.; Salman Qureshi, O.; Maria, P.; Hamzah, Z.; Javed, O. Role of Flavonoids as Wound Healing Agent. Intech 2016. [Google Scholar] [CrossRef] [Green Version]
- Elmowafy, E.; El-Derany, M.O.; Biondo, F.; Tiboni, M.; Casettari, L.; Soliman, M.E. Quercetin loaded monolaurate sugar esters-based niosomes: Sustained release and mutual antioxidant—hepatoprotective interplay. Pharmaceutics 2020, 12, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.S.; Park, S.H.; Kim, M.S.; Han, S.Y.; Kim, H.S.; Kang, Y.H. Caffeic acid disturbs monocyte adhesion onto cultured endothelial cells stimulated by adipokine resistin. J. Agric. Food Chem. 2012, 60, 2730–2739. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Jakaria, M.; Kim, I.S.; Kim, J.; Ezazul Haque, M.; Choi, D.K. Regulation of toll-like receptor (TLR) signaling pathway by polyphenols in the treatment of age-linked neurodegenerative diseases: Focus on TLR4 signaling. Front. Immunol. 2019, 10, 1000. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No | Compound | Free PSF (µg/g) | PSF-DOPCs (µg/g) | PSF-DOPGs (µg/g) |
|---|---|---|---|---|
| 1 | 3-O-caffeoylquinic acid | 2112.0 ± 23.3 | 418.4 ± 13.0 | 94.0 ± 1.3 |
| 2 | 3-O-p-cumaroylquinic acid | 70.0 ± 3.9 | 9.6 ± 0.1 | 3.5 ± 0.2 |
| 3 | Chlorogenic acid dehydrodimer | 75.8 ± 0.8 | 11.2 ± 0.1 | 2.7 ± 0.3 |
| 4 | 3-O-feruloylquinic acid | 218.4 ± 1.4 | 37.2 ± 0.3 | 9.3 ± 0.3 |
| 5 | 4-O-caffeoylquinic acid | 187.3 ± 3.9 | 41.2 ± 0.3 | 8.7 ± 0.1 |
| 7 | Chlorogenic acid dehydrodimer | 184.5 ± 2.3 | 24.8 ± 0.1 | 6.9 ± 0.2 |
| Total hydroxycinnamic acid derivatives | 2847.9 ± 23.5 | 542.3 ± 12.5 | 125.1 ± 1.5 | |
| 8 | Cyanidin 3-O-glucoside | 490.5 ± 6.3 | 26.4 ± 0.1 | 13.3 ± 0.1 |
| 10 | Cyanidin 3-O-rutinoside | 638.9 ± 1.9 | 11.2 ± 0.1 | 29.4 ± 0.2 |
| 11 | Peonidin 3-O-glucoside | 315.0 ± 4.4 | 4.6 ± 0.1 | 3.9 ± 0.2 |
| 12 | Peonidin 3-O-rutinoside | 650.3 ± 0.8 | 7.4 ± 0.1 | 7.2 ± 0.1 |
| Total anthocyanins | 2094.8 ± 11.8 | 49.7 ± 0.1 | 53.8 ± 0.1 | |
| 15 | Ellagic acid derivative | 40.6 ± 1.5 | 19.7 ± 0.2 | 10.1 ± 0.1 |
| 16 | 4-(vanilloyloxy)-2,6,6-trimethylcyclohexene-1-carboxylic acid | 74.6 ± 2.2 | 40.6 ± 0.7 | 40.6 ± 0.2 |
| Total hydroxybenzoic acid derivatives | 115.1 ± 0.7 | 60.3 ± 2.3 | 50.7 ± 0.3 | |
| 6 | Apigenin pentoside | 27.4 ± 0.1 | 9.9 ± 0.1 | 3.9 ± 0.1 |
| 9 | Apigenin pentoside isomer | 28.1 ± 1.6 | 6.3 ± 0.1 | 4.3 ± 0.1 |
| 17 | Quercetin hexoside | 240.7 ± 6.4 | 64.6 ± 0.1 | 54.5 ± 0.1 |
| 18 | Quercetin 3-O-hexoside-O-pentoside | 145.3 ± 1.7 | 78.0 ± 0.1 | 24.6 ± 0.2 |
| 19 | Rutin | 168.9 ± 1.2 | 75.8 ± 0.2 | 23.9 ± 0.1 |
| 20 | Quercetin galactoside | 366.8 ± 4.8 | 215.7 ± 0.1 | 73.7 ± 0.2 |
| 21 | Quercetin xyloside | 29.5 ± 0.4 | 29.0 ± 0.1 | 4.1 ± 0.1 |
| 22 | Quercetin arabinoside | 107.7 ± 0.4 | 78.9 ± 0.2 | 22.5 ± 0.1 |
| 23 | Quercetin pentoside | 193.1 ± 1.1 | 108.0 ± 0.1 | 23.2 ± 0.1 |
| 24 | Quercetin 3-O-rhamnoside | 43.6 ± 0.3 | 20.8 ± 0.1 | n.d. |
| Total flavonoid derivatives | 1351.0 ± 8.4 | 687.1 ± 13.0 | 234.7 ± 0.1 | |
| Total phenolic compounds | 6408.9 ± 4.1 | 1339.4 ± 13.1 | 464.3 ± 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiboni, M.; Coppari, S.; Casettari, L.; Guescini, M.; Colomba, M.; Fraternale, D.; Gorassini, A.; Verardo, G.; Ramakrishna, S.; Guidi, L.; et al. Prunus spinosa Extract Loaded in Biomimetic Nanoparticles Evokes In Vitro Anti-Inflammatory and Wound Healing Activities. Nanomaterials 2021, 11, 36. https://doi.org/10.3390/nano11010036
Tiboni M, Coppari S, Casettari L, Guescini M, Colomba M, Fraternale D, Gorassini A, Verardo G, Ramakrishna S, Guidi L, et al. Prunus spinosa Extract Loaded in Biomimetic Nanoparticles Evokes In Vitro Anti-Inflammatory and Wound Healing Activities. Nanomaterials. 2021; 11(1):36. https://doi.org/10.3390/nano11010036
Chicago/Turabian StyleTiboni, Mattia, Sofia Coppari, Luca Casettari, Michele Guescini, Mariastella Colomba, Daniele Fraternale, Andrea Gorassini, Giancarlo Verardo, Seeram Ramakrishna, Loretta Guidi, and et al. 2021. "Prunus spinosa Extract Loaded in Biomimetic Nanoparticles Evokes In Vitro Anti-Inflammatory and Wound Healing Activities" Nanomaterials 11, no. 1: 36. https://doi.org/10.3390/nano11010036