
Recent Progress of In Vitro 3D Culture of Male Germ Stem Cells

Abstract
:1. Introduction
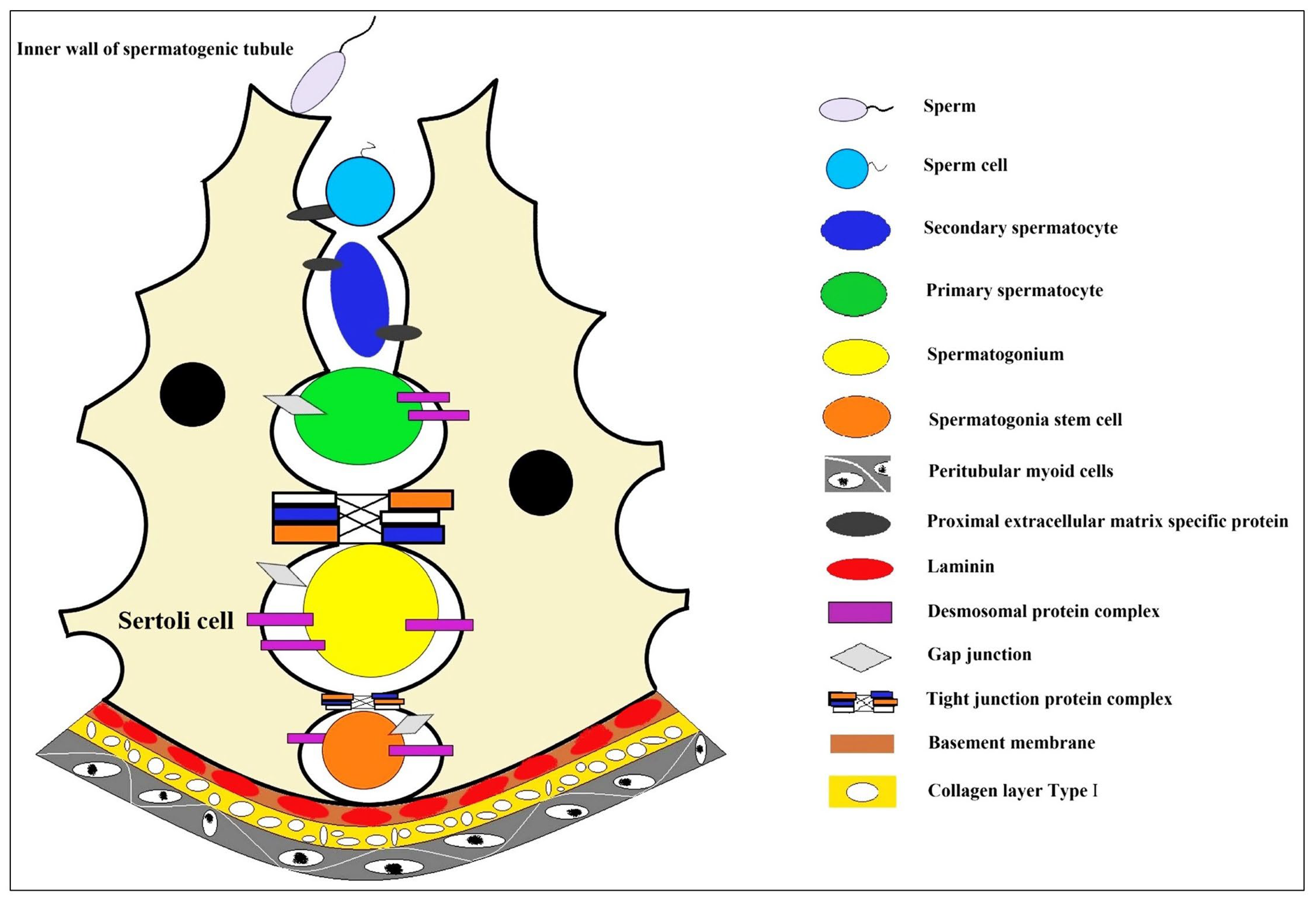
2. The Niche of Spermatogonial Stem Cells (SSCs)
2.1. The Spermatogonial Stem Cells
2.2. The Microenvironment of Spermatocyte Development
3. The 3D Cell Culture of SSCs
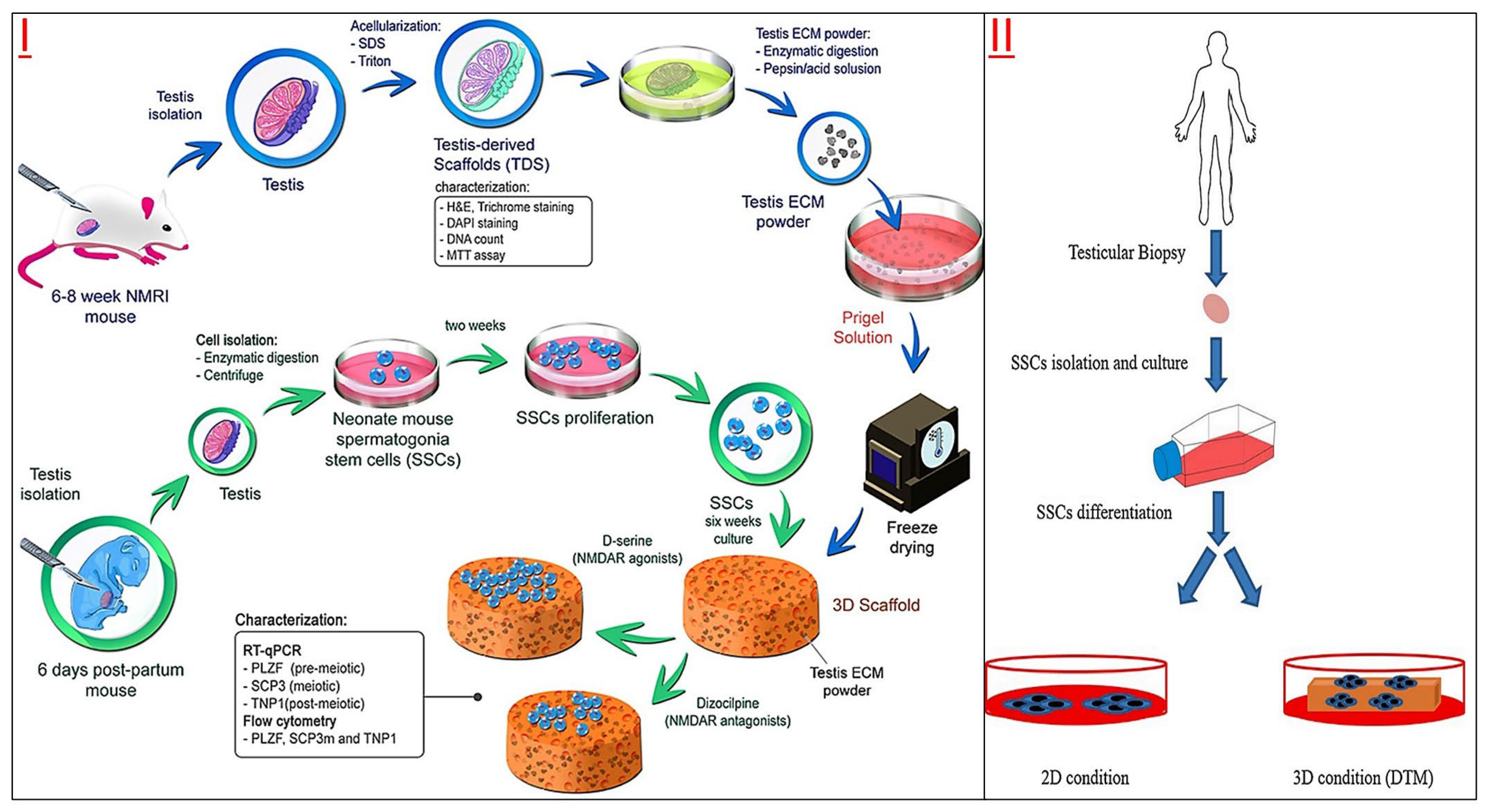
3.1. Scaffold-Based 3D Cell Culture for SSCs
3.1.1. Scaffold-Based 3D Cell Culture
3.1.2. DTM-Based Scaffold Culture for SSCs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Material | Cell Type | Species | Main Biological Findings | Reference |
|---|---|---|---|---|
| Sertoli cells and Leydig cells with extracellular matrix (ECM) composition | SSCs | Mice | The coculture 3D structure prepares an in vivo-like niche and supports the proliferation of germ cells. | [66] |
| An artificial testicular tissue using a DTM -hyaluronic gel matrix | SSCs | Mice | The decellularized testicular matrix supplemented with D-serine and glutamic acid could provide an appropriate niche environment for the proliferation of SSCs. | [67] |
| DTM hydrogel | SSCs | Mice | The differentiation of spermatogonia could be regulated by D-serine in the 3D culture system. | [37] |
| Azoospermia tissue DTM | SSCs | NMRI mice | The presence of D-serine and retinoic acid has a positive effect on spermatogenesis in the 3D culture system. | [68] |
| Sheep DTM | SSCs | Human | The natural structure of DTM prepares the suitable niche environment for the spermatogenesis in vitro. | [69] |
| Sheep DTM | SSCs | Human | SSCs culture in DTM created a way of demonstrating spermatogenesis in vitro. | [70] |
| Human DTM | SSCs | Human | Despite the lack of testis-specific tissue structure, three-dimensional culture in vitro could harbor spermatogonium cells and provide their essential niche environment, so that these spermatogonium cells retain specific functions in long-term culture. These findings also open up the possibility of recreating the testicular microenvironment (such as organoid tissue) from primary testicular cells in vitro. | [71,72] |
| ECM | SSCs | Rats | In the 3D rat testicular cell co-culture model, the proliferation, differentiation, and androgen receptor (AR) protein expression of spermatogonia cells could be regulated by experimental methods. | [73] |
| Human DTM | SSCs | Human | The niche microenvironment created by the multicellular 3D testis organoid model could maintain the long-term viability of spermatogonia cells. It could also promote the differentiation of SSCs into postmeiotic germ cells, simulating the process of spermatogenesis in vivo, so that about 0.2% of SSCs differentiate into sperm cells. | [74] |
3.1.3. Non-DTM-Based Scaffold Culture for SSCs
| Culture Material | Cell Type | Species | Main Biological Findings | Reference |
|---|---|---|---|---|
| Alginic acid | SSCs | Mice | Alginate scaffold structure could maintain the morphology and cell density of SSCs for a long time, and promote the expression of pluripotent genes. | [75] |
| Gonadal somatic cells and transwell-COL membranes | SSCs | Mice | In a 3D organoid culture system, SSCs are transformed into induced germline stem cells (iGSCs) with maternal imprinting patterns through transgenic manipulation. | [76] |
| Agar/polyvinyl alcohol nanofiber scaffold | SSCs | Mice | In the three-dimensional culture system of AGAR/PVA scaffold, the differentiation of mouse SSCs into spermatoblasts could be enhanced synergically with the medium supplemented with growth factors. | [77] |
| PRP + CaCl2 | SSCs | Human | PRP scaffold could reconstruct a suitable niche environment for the in vitro proliferation of SSCs. | [78] |
| 3D Stemfit® culture dishes (3D scaffold)(MicroFIT, Seongnam, Korea) | SSCs | Mice | Using 3D scaffolds, SSCs could be reprogrammed to become gPSCs without biological substrates. | [79] |
| Agarose gel stands | SSC-LCs derived from iPSCs | Mice | iPSCs could hom in a three-dimensional testicular niche environment, which plays a crucial role in inducing iPSCs to differentiate into spermatogonial stem cell-like cells. | [80] |
| Human Sertoli cells | SSCs | Human | 3D culture could significantly increase the number and size of SSCs clones and the expression of spermatogonial marker genes. | [81] |
| Agarose | SSCs | Mice | Agarose 3D culture induced spermatogenesis process in vitro. | [82] |
| Methylcellulose Culture System (MCS) | SSCs | Mice | The MCS 3D culture system could induce the differentiation of normal immature spermatogonium into meiotic and postmeiotic cells and produce sperm-like cells. | [83] |
| MCS AND SACS | STC | Rhesus monkeys | The 3D culture system could partially simulate the microenvironment in the seminiferous tubules and promote the differentiation of type A spermatogonium to spermatocyte. | [84] |
| Collagen gel matrix | SSCs | Balb-c mice | The three-dimensional culture system of collagen gel provided a microenvironment that mimics the spermatogenic epithelium and could induce spermatogenic processes in spermatogonium in vitro. | [85] |
| Poly L-lactic acid(PLLA) nanofiber scaffold | SSCs | Mice | PLLA could promote the formation of spermatogonia clones and induce cell differentiation during culture | [86] |
| Methylcellulose(MCS) | SSCs | Human | 3D MCS culture system could induce spermatogenic processes of spermatogonium isolated from living tissue | [87] |
| Sertoli cells in SACS + laminin + growth factors | SSCs | Human | Laminin could replace Sertoli cells to construct a three-dimensional culture system, that enables specific spermatogonium to self-renew or differentiate | [89] |
| Sertoli cell feeder layer in goat | SSCs | Sheep | In a culture system with Sertoli cells as feeder layers, SSCs could emerge as cell clones after a short time culture. | [90] |
| SIM mouse(Sando’s inbred mouse) embryo-derived thioguanine and ouabain resistant (STO), and a laminin-coated plate. | SSCs | Bovine | In the culture system of the STO feeder layer, SSCs could form many cell clones and express SSCs marker genes at a high level. In the three-dimensional culture system of laminin, the pluripotent genes of SSCs were highly expressed. | [91] |
| DSA lectin-coated dishes with the attachment of Sertoli cells. | SSCs | Buffalo | DSA lectin-coated dishes supported long-term maintenance and self-renewal of SSC-like cells. | [92] |
| NMRI Mouse STO and Growth Factors | SSCs | Mice | The SSC-STO co-culture provided a microenvironment for efficient maintenance and proliferation of SSCs. | [93] |
| 3D alginate hydrogel with Sertoli cells | SSCs | Mice | Alginate gel three-dimensional culture could promote the proliferation of SSCs and the maintenance of cell stemness and improve the survival rate of SSCs transplantation. | [94] |
| Sertoli, and Leydig cells | SSCs | Murine C18-4 | The testicular co-culture model could make spermatogonial cells present better cell structure in vitro and promote intercellular communication between different cell types. | [95] |
| Membrane-bottomed microwell array added to Transwell insert + STO cells | SSCs | Mice | The microporous culture system of the STO cell feeding layer could promote the formation of SSCs cell clonoid structure more efficiently. | [96] |

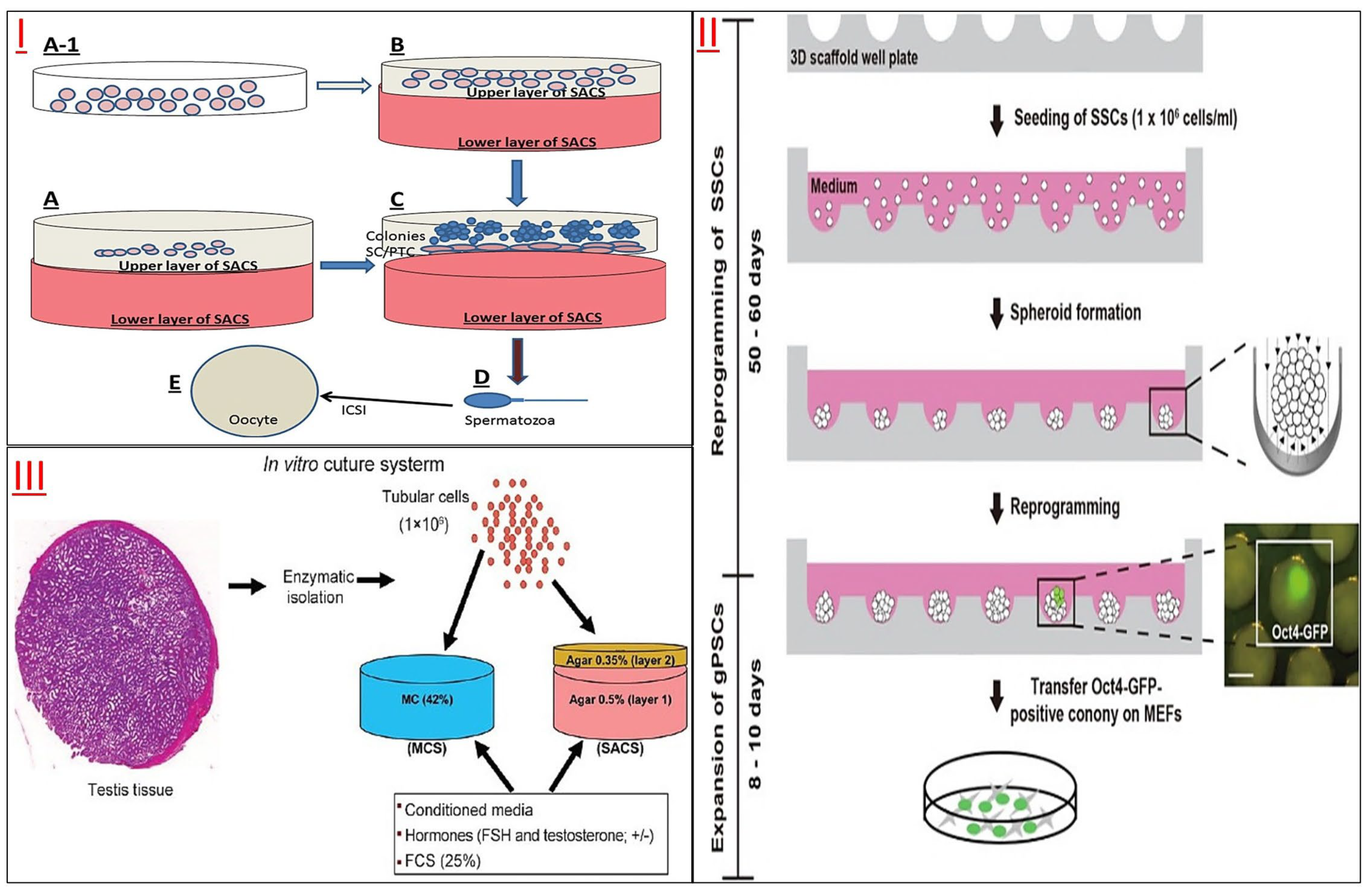
3.2. 3D Printing Cell Culture for SSCs
3.2.1. 3D Printing Cell Culture
3.2.2. 3D-Printed Scaffold Culture for SSCs

| Culture Form | Culture Material | Cell Type | Species | Main Biological Findings | Reference |
|---|---|---|---|---|---|
| Cells no-printed | Ram ECM + Alg-Gel | SSCs | NMRI mice | Testicular cells cultured on T-ECM-enriched scaffolds showed high cell viability and expressed genes related to spermatogonial differentiation. | [111] |
| Agarose gel | SSCs | Mice | In the double-layer scaffold culture system, the organoid structure formed by the mixture of primary testicular cells and germline stem cells showed a curved spermatotubule-like microenvironment structure. | [112] | |
| Ram DTM with alginate–gelatin | SSCs | Mice | The 3D-printed scaffold of 5% T-ECM could enable good adhesion of SSCs and promote their cell survival in vitro. | [38] | |
| Cells printed | PCL powder and gelatin | SSCs | Human | The PCL powder and gelatin nanocomposite scaffold could provide a suitable microenvironment for self-renewal of SSCs | [114] |
| Hydrogels comprised of alginate and gelatin | MCS | Sheep | The 3D-printed culture system of 7%Alg-8%Gel mixture could maintain good cell viability, and the cell function could be kept for a long time when incubated with the cell-loaded hydrogel. | [115,116] | |
| AGC-10 matrix bioink composed of alginate and rat tail-derived collagen | SSCs | Patients with NOA | After bioprinting, human testicular cells could proliferate well, have high cell viability, and maintain their biological function. | [97] |
3.2.3. 3D-Printed Scaffold-SSCs Matrix
4. Conclusions and Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Brinster, R.L. Male germline stem cells: From mice to men. Science 2007, 316, 404–405. [Google Scholar] [CrossRef]
- Lei, Q.; Lai, X.; Eliveld, J.; Chuva, D.S.L.S.; van Pelt, A.; Hamer, G. In Vitro meiosis of male germline stem cells. Stem Cell Rep. 2020, 15, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Yang, D.; Du, X.; Yu, X.; Zhang, M.; Tang, F.; Ma, F.; Li, N.; Bai, C.; Li, G.; et al. Interaction between dmrt1 and plzf protein regulates self-renewal and proliferation in male germline stem cells. Mol. Cell. Biochem. 2021, 476, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Ma, Y.D.; Liu, S.Q.; Huang, R.W.; Chen, J.J.; An, L.L.; Wu, J. SARS-CoV-2 structural proteins modulated blood-testis barrier-related proteins through autophagy in the primary sertoli cells. Viruses 2023, 15, 1272. [Google Scholar] [CrossRef]
- Wei, Y.D.; Du, X.M.; Yang, D.H.; Ma, F.L.; Yu, X.W.; Zhang, M.F.; Li, N.; Peng, S.; Liao, M.Z.; Li, G.P.; et al. Dmrt1 regulates the immune response by repressing the tlr4 signaling pathway in goat male germline stem cells. Zool. Res. 2021, 42, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H.; Brinster, R.L. Technology insight: In Vitro culture of spermatogonial stem cells and their potential therapeutic uses. Nat. Clin. Pract. Endocrinol. Metab. 2006, 2, 99–108. [Google Scholar] [CrossRef]
- Dorn, D.C.; Dorn, A. Structural characterization and primary in vitro cell culture of locust male germline stem cells and their niche. Stem Cell Res. 2011, 6, 112–128. [Google Scholar] [CrossRef]
- Lei, Q.; Pan, Q.; Li, N.; Zhou, Z.; Zhang, J.; He, X.; Peng, S.; Li, G.; Sidhu, K.; Chen, S.; et al. H19 regulates the proliferation of bovine male germline stem cells via igf-1 signaling pathway. J. Cell. Physiol. 2018, 234, 915–926. [Google Scholar] [CrossRef]
- Kubota, H.; Brinster, R.L. Culture of rodent spermatogonial stem cells, male germline stem cells of the postnatal animal. Methods Cell Biol. 2008, 86, 59–84. [Google Scholar]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Iwano, T.; Lee, J.; Kazuki, Y.; Inoue, K.; Miki, H.; Takehashi, M.; Toyokuni, S.; Shinkai, Y.; et al. Genetic and epigenetic properties of mouse male germline stem cells during long-term culture. Development 2005, 132, 4155–4163. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Inoue, K.; Miki, H.; Ogura, A.; Toyokuni, S.; Shinohara, T. Long-term proliferation in culture and germline transmission of mouse male germline stem cells. Biol. Reprod. 2003, 69, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Jamaluddin, M.; Ghosh, A.; Ingle, A.; Mohammed, R.; Ali, A.; Bahrami, M.; Kaiko, G.; Gibb, Z.; Filipe, E.C.; Cox, T.R.; et al. Bovine and human endometrium-derived hydrogels support organoid culture from healthy and cancerous tissues. Proc. Natl. Acad. Sci. USA 2022, 119, e2086927177. [Google Scholar] [CrossRef]
- Nagano, M.; Hu, B.; Yokobayashi, S.; Yamamura, A.; Umemura, F.; Coradin, M.; Ohta, H.; Yabuta, Y.; Ishikura, Y.; Okamoto, I.; et al. Nucleome programming is required for the foundation of totipotency in mammalian germline development. EMBO J. 2022, 41, e110600. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Yuan, Q.; Niu, M.; Wang, H.; Wen, L.; Yao, C.; Hou, J.; Chen, Z.; Fu, H.; Zhou, F.; et al. Efficient generation of functional haploid spermatids from human germline stem cells by three-dimensional-induced system. Cell Death Differ. 2018, 25, 749–766. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Wu, S.; Wei, Y.; Yu, X.; Ma, F.; Zhai, Y.; Yang, D.; Zhang, M.; Liu, W.; Zhu, H.; et al. Pax7 promotes cd49f-positive dairy goat spermatogonial stem cells’ self-renewal. J. Cell. Physiol. 2021, 236, 1481–1493. [Google Scholar] [CrossRef]
- Ibtisham, F.; Zhao, Y.; Wu, J.; Nawab, A.; Mei, X.; Li, G.; An, L. The optimized condition for the isolation and in vitro propagation of mouse spermatogonial stem cells. Biol. Futur. 2019, 70, 79–87. [Google Scholar] [CrossRef]
- Wang, H.; Wen, L.; Yuan, Q.; Sun, M.; Niu, M.; He, Z. Establishment and applications of male germ cell and sertoli cell lines. Reproduction 2016, 152, R31–R40. [Google Scholar] [CrossRef]
- Ho, N.P.; Takizawa, H. Inflammation regulates haematopoietic stem cells and their niche. Int. J. Mol. Sci. 2022, 23, 1125. [Google Scholar] [CrossRef]
- Shwartz, Y.; Gonzalez-Celeiro, M.; Chen, C.L.; Pasolli, H.A.; Sheu, S.H.; Fan, S.M.; Shamsi, F.; Assaad, S.; Lin, E.T.; Zhang, B.; et al. Cell types promoting goosebumps form a niche to regulate hair follicle stem cells. Cell 2020, 182, 578–593. [Google Scholar] [CrossRef]
- Voog, J.; Jones, D.L. Stem cells and the niche: A dynamic duo. Cell Stem Cell 2010, 6, 103–115. [Google Scholar] [CrossRef]
- Song, W.; Zhu, H.; Li, M.; Li, N.; Wu, J.; Mu, H.; Yao, X.; Han, W.; Liu, W.; Hua, J. Promyelocytic leukaemia zinc finger maintains self-renewal of male germline stem cells (mgscs) and its expression pattern in dairy goat testis. Cell Prolif. 2013, 46, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Li, B.; Wu, C.; Wu, J.; Yan, Y.; Shang, R.; Bai, C.; Li, G.; Hua, J. Melatonin promotes goat spermatogonia stem cells (sscs) proliferation by stimulating glial cell line-derived neurotrophic factor (gdnf) production in sertoli cells. Oncotarget 2016, 7, 77532–77542. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Mu, H.; Wu, J.; Liao, M.; Zhu, H.; Zheng, L.; He, X.; Niu, B.; Zhai, Y.; Bai, C.; et al. Mir-544 regulates dairy goat male germline stem cell self-renewal via targeting plzf. J. Cell. Biochem. 2015, 116, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Niu, B.; Wu, C.; Hua, J.; Wu, J. The construction and application of lentiviral overexpression vector of goat mir-204 in testis. Res. Vet. Sci. 2020, 130, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Diao, L.; Turek, P.J.; John, C.M.; Fang, F.; Reijo, P.R. Roles of spermatogonial stem cells in spermatogenesis and fertility restoration. Front. Endocrinol. 2022, 13, 895528. [Google Scholar] [CrossRef] [PubMed]
- Yokonishi, T.; Mckey, J.; Ide, S.; Capel, B. Sertoli cell ablation and replacement of the spermatogonial niche in mouse. Nat. Commun. 2020, 11, 40. [Google Scholar] [CrossRef]
- Blomberg, J.M.; Andreassen, C.H.; Jorgensen, A.; Nielsen, J.E.; Juel, M.L.; Boisen, I.M.; Schwarz, P.; Toppari, J.; Baron, R.; Lanske, B.; et al. Rankl regulates male reproductive function. Nat. Commun. 2021, 12, 2450. [Google Scholar] [CrossRef]
- Griswold, M.D. Interactions between germ cells and sertoli cells in the testis. Biol. Reprod. 1995, 52, 211–216. [Google Scholar] [CrossRef]
- Chaicharoenaudomrung, N.; Kunhorm, P.; Noisa, P. Three-dimensional cell culture systems as an in vitro platform for cancer and stem cell modeling. World J. Stem Cells 2019, 11, 1065–1083. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, J.R.; Park, S.; Kim, Y.J.; Yoon, J.K.; Park, H.S.; Hyun, J.; Joung, Y.K.; Lee, T.I.; Bhang, S.H. Subaqueous 3d stem cell spheroid levitation culture using anti-gravity bioreactor based on sound wave superposition. Biomater. Res. 2023, 27, 51. [Google Scholar] [CrossRef]
- Kim, J.A.; Hong, S.; Rhee, W.J. Microfluidic three-dimensional cell culture of stem cells for high-throughput analysis. World J. Stem Cells 2019, 11, 803–816. [Google Scholar] [CrossRef]
- Baena, Y.A.R.; Casasco, A.; Monti, M. Hypes and hopes of stem cell therapies in dentistry: A review. Stem Cell Rev. Rep. 2022, 18, 1294–1308. [Google Scholar] [CrossRef]
- Park, Y.; Huh, K.M.; Kang, S.W. Applications of biomaterials in 3d cell culture and contributions of 3d cell culture to drug development and basic biomedical research. Int. J. Mol. Sci. 2021, 22, 2491. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is it time to start transitioning from 2d to 3d cell culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef]
- Wu, X.; Su, J.; Wei, J.; Jiang, N.; Ge, X. Recent advances in three-dimensional stem cell culture systems and applications. Stem Cells Int. 2021, 2021, 9477332. [Google Scholar] [CrossRef]
- Gallagher, C.; Murphy, C.; O’Brien, F.J.; Piskareva, O. Three-dimensional in vitro biomimetic model of neuroblastoma using collagen-based scaffolds. J. Vis. Exp. 2021, 173, 62627. [Google Scholar]
- Noghani, A.E.; Asadpour, R.; Saberivand, A.; Mazaheri, Z.; Rodriguez-Wallberg, K.A.; Hamidian, G. Differentiation of neonate mouse spermatogonia on two-dimensional and three-dimensional culture systems supplemented with d-serine and dizocilpine (mk-801). Theriogenology 2022, 191, 168–178. [Google Scholar] [CrossRef]
- Bashiri, Z.; Amiri, I.; Gholipourmalekabadi, M.; Falak, R.; Asgari, H.; Maki, C.B.; Moghaddaszadeh, A.; Koruji, M. Artificial testis: A testicular tissue extracellular matrix as a potential bio-ink for 3d printing. Biomater. Sci. 2021, 9, 3465–3484. [Google Scholar] [CrossRef]
- Ricci, C.; Azimi, B.; Panariello, L.; Antognoli, B.; Cecchini, B.; Rovelli, R.; Rustembek, M.; Cinelli, P.; Milazzo, M.; Danti, S.; et al. Assessment of electrospun poly(epsilon-caprolactone) and poly(lactic acid) fiber scaffolds to generate 3d in vitro models of colorectal adenocarcinoma: A preliminary study. Int. J. Mol. Sci. 2023, 24, 9443. [Google Scholar] [CrossRef]
- Kolehmainen, K.; Willerth, S.M. Preparation of 3d fibrin scaffolds for stem cell culture applications. J. Vis. Exp. 2012, 61, e3641. [Google Scholar]
- Kim, S.; Min, S.; Choi, Y.S.; Jo, S.H.; Jung, J.H.; Han, K.; Kim, J.; An, S.; Ji, Y.W.; Kim, Y.G.; et al. Tissue extracellular matrix hydrogels as alternatives to matrigel for culturing gastrointestinal organoids. Nat. Commun. 2022, 13, 1692. [Google Scholar] [CrossRef]
- Willemse, J.; van Tienderen, G.; van Hengel, E.; Schurink, I.; van der Ven, D.; Kan, Y.; de Ruiter, P.; Rosmark, O.; Westergren-Thorsson, G.G.; Schneeberger, K.; et al. Hydrogels derived from decellularized liver tissue support the growth and differentiation of cholangiocyte organoids. Biomaterials 2022, 284, 121473. [Google Scholar] [CrossRef]
- Lou, J.; Stowers, R.; Nam, S.; Xia, Y.; Chaudhuri, O. Stress relaxing hyaluronic acid-collagen hydrogels promote cell spreading, fiber remodeling, and focal adhesion formation in 3d cell culture. Biomaterials 2018, 154, 213–222. [Google Scholar] [CrossRef]
- Dou, X.Q.; Feng, C.L. Amino acids and peptide-based supramolecular hydrogels for three-dimensional cell culture. Adv. Mater. 2017, 29, 1604062. [Google Scholar] [CrossRef]
- Shi, X.; Janmey, P.A. Large polyacrylamide hydrogels for large-batch cell culture and mechanobiological studies. Macromol. Biosci. 2023, 23, e2300042. [Google Scholar] [CrossRef]
- Simaan-Yameen, H.; Bar-Am, O.; Saar, G.; Seliktar, D. Methacrylated fibrinogen hydrogels for 3d cell culture and delivery. Acta Biomater. 2023, 164, 94–110. [Google Scholar] [CrossRef]
- Liu, Q.; Dai, W.; Gao, Y.; Dong, L.; Jia, H.; Li, S.; Guo, L.; Fan, Y.; Zhang, X. The synergistic regulation of chondrogenesis by collagen-based hydrogels and cell co-culture. Acta Biomater. 2022, 154, 194–211. [Google Scholar] [CrossRef]
- Jury, M.; Matthiesen, I.; Rasti, B.F.; Ludwig, S.L.; Civitelli, L.; Winkler, T.E.; Selegard, R.; Herland, A.; Aili, D. Bioorthogonally cross-linked hyaluronan-laminin hydrogels for 3d neuronal cell culture and biofabrication. Adv. Healthc. Mater. 2022, 11, e2102097. [Google Scholar] [CrossRef]
- Szkolar, L.; Guilbaud, J.B.; Miller, A.F.; Gough, J.E.; Saiani, A. Enzymatically triggered peptide hydrogels for 3d cell encapsulation and culture. J. Pept. Sci. 2014, 20, 578–584. [Google Scholar] [CrossRef]
- Xu, X.; Feng, Q.; Ma, X.; Deng, Y.; Zhang, K.; Ooi, H.S.; Yang, B.; Zhang, Z.Y.; Feng, B.; Bian, L. Dynamic gelatin-based hydrogels promote the proliferation and self-renewal of embryonic stem cells in long-term 3d culture. Biomaterials 2022, 289, 121802. [Google Scholar] [CrossRef]
- Lou, J.; Mooney, D.J. Chemical strategies to engineer hydrogels for cell culture. Nat. Rev. Chem. 2022, 6, 726–744. [Google Scholar] [CrossRef] [PubMed]
- Norman, M.; Ferreira, S.A.; Jowett, G.M.; Bozec, L.; Gentleman, E. Measuring the elastic modulus of soft culture surfaces and three-dimensional hydrogels using atomic force microscopy. Nat. Protoc. 2021, 16, 2418–2449. [Google Scholar] [CrossRef] [PubMed]
- Salahuddin, B.; Wang, S.; Sangian, D.; Aziz, S.; Gu, Q. Hybrid gelatin hydrogels in nanomedicine applications. ACS Appl. Bio Mater. 2021, 4, 2886–2906. [Google Scholar] [CrossRef]
- Shah, D.D.; Raghani, N.R.; Chorawala, M.R.; Singh, S.; Prajapati, B.G. Harnessing three-dimensional (3d) cell culture models for pulmonary infections: State of the art and future directions. Naunyn Schmiedebergs Arch. Pharmacol. 2023, 396, 2861–2880. [Google Scholar] [CrossRef] [PubMed]
- Knight, E.; Przyborski, S. Advances in 3d cell culture technologies enabling tissue-like structures to be created in vitro. J. Anat. 2015, 227, 746–756. [Google Scholar] [CrossRef]
- Bassi, G.; Panseri, S.; Dozio, S.M.; Sandri, M.; Campodoni, E.; Dapporto, M.; Sprio, S.; Tampieri, A.; Montesi, M. Scaffold-based 3d cellular models mimicking the heterogeneity of osteosarcoma stem cell niche. Sci. Rep. 2020, 10, 22294. [Google Scholar] [CrossRef]
- El, H.S.; Zidi, B.; El, H.N.; Krause, K.H.; Matthes, T.; Preynat-Seauve, O. Cancer spheroids and organoids as novel tools for research and therapy: State of the art and challenges to guide precision medicine. Cells 2023, 12, 1001. [Google Scholar]
- Wishart, A.L.; Conner, S.J.; Guarin, J.R.; Fatherree, J.P.; Peng, Y.; Mcginn, R.A.; Crews, R.; Naber, S.P.; Hunter, M.; Greenberg, A.S.; et al. Decellularized extracellular matrix scaffolds identify full-length collagen vi as a driver of breast cancer cell invasion in obesity and metastasis. Sci. Adv. 2020, 6, eabc3175. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Hong, H.; Hu, R.; Liu, J.; Liu, C. Decellularized extracellular matrix scaffolds: Recent trends and emerging strategies in tissue engineering. Bioact. Mater. 2022, 10, 15–31. [Google Scholar] [CrossRef]
- Kasravi, M.; Ahmadi, A.; Babajani, A.; Mazloomnejad, R.; Hatamnejad, M.R.; Shariatzadeh, S.; Bahrami, S.; Niknejad, H. Immunogenicity of decellularized extracellular matrix scaffolds: A bottleneck in tissue engineering and regenerative medicine. Biomater. Res. 2023, 27, 10. [Google Scholar] [CrossRef]
- Hogan, K.J.; Smoak, M.M.; Koons, G.L.; Perez, M.R.; Bedell, M.L.; Jiang, E.Y.; Young, S.; Mikos, A.G. Bioinspired electrospun decellularized extracellular matrix scaffolds promote muscle regeneration in a rat skeletal muscle defect model. J. Biomed. Mater. Res. A 2022, 110, 1090–1100. [Google Scholar] [CrossRef]
- Barbulescu, G.I.; Bojin, F.M.; Ordodi, V.L.; Goje, I.D.; Barbulescu, A.S.; Paunescu, V. Decellularized extracellular matrix scaffolds for cardiovascular tissue engineering: Current techniques and challenges. Int. J. Mol. Sci. 2022, 23, 13040. [Google Scholar] [CrossRef]
- Zhu, W.; Cao, L.; Song, C.; Pang, Z.; Jiang, H.; Guo, C. Cell-derived decellularized extracellular matrix scaffolds for articular cartilage repair. Int. J. Artif. Organs 2021, 44, 269–281. [Google Scholar] [CrossRef]
- Agmon, G.; Christman, K.L. Controlling stem cell behavior with decellularized extracellular matrix scaffolds. Curr. Opin. Solid. State Mater. Sci. 2016, 20, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, S.; Fernandes, H.; Moroni, L. Decellularized extracellular matrix scaffolds for cartilage regeneration. Methods Mol. Biol. 2015, 1340, 133–151. [Google Scholar] [PubMed]
- Yin, L.; Wei, H.; Liang, S.; Yu, X. From the cover: An animal-free in vitro three-dimensional testicular cell coculture model for evaluating male reproductive toxicants. Toxicol. Sci. 2017, 159, 307–326. [Google Scholar] [CrossRef]
- Noghani, A.E.; Asadpour, R.; Saberivand, A.; Mazaheri, Z.; Hamidian, G. Effect of nmda receptor agonist and antagonist on spermatogonial stem cells proliferation in 2- and 3- dimensional culture systems. Mol. Biol. Rep. 2022, 49, 2197–2207. [Google Scholar] [CrossRef] [PubMed]
- Modirshanechi, G.; Eslampour, M.A.; Abdolmaleki, Z. Agonist and antagonist nmda receptor effect on cell fate during germ cell differentiation and regulate apoptotic process in 3d organ culture. Andrologia 2020, 52, e13764. [Google Scholar] [CrossRef]
- Ashouri, M.S.; Ashouri, M.S.; Banitalebi, D.M.; Pourmand, G.; Gholami, K.; Talebi, A.; Esfandyari, S.; Jabari, A.; Samadian, A.; Abbasi, M. Isolation, identification and differentiation of human spermatogonial cells on three-dimensional decellularized sheep testis. Acta Histochem. 2020, 122, 151623. [Google Scholar] [CrossRef]
- Ashouri, M.S.; Banitalebi, D.M.; Koruji, M.; Pourmand, G.; Farzaneh, P.; Ashouri, M.S.; Jabari, A.; Samadian, A.; Khadivi, F.; Abbasi, M. In Vitro spermatogenesis by three-dimensional culture of spermatogonial stem cells on decellularized testicular matrix. Galen. Med. J. 2019, 8, e1565. [Google Scholar] [CrossRef]
- Baert, Y.; De Kock, J.; Alves-Lopes, J.P.; Soder, O.; Stukenborg, J.B.; Goossens, E. Primary human testicular cells self-organize into organoids with testicular properties. Stem Cell Rep. 2017, 8, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Baert, Y.; Stukenborg, J.B.; Landreh, M.; De Kock, J.; Jornvall, H.; Soder, O.; Goossens, E. Derivation and characterization of a cytocompatible scaffold from human testis. Hum. Reprod. 2015, 30, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, Y.; Tian, Y.; Yan, H.; Ren, L.; Shi, W.; Zhu, J.; Zhang, T. The application of the improved 3d rat testicular cells co-culture model on the in vitro toxicity research of hz1006. Drug Chem. Toxicol. 2019, 42, 526–535. [Google Scholar] [CrossRef]
- Pendergraft, S.S.; Sadri-Ardekani, H.; Atala, A.; Bishop, C.E. Three-dimensional testicular organoid: A novel tool for the study of human spermatogenesis and gonadotoxicity in vitro. Biol. Reprod. 2017, 96, 720–732. [Google Scholar] [CrossRef]
- Hemadi, M.; Assadollahi, V.; Saki, G.; Pirnia, A.; Alasvand, M.; Zendehdel, A.; Gholami, M. Use of alginate hydrogel to improve long-term 3d culture of spermatogonial stem cells: Stemness gene expression and structural features. Zygote 2022, 30, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Li, X.; Tian, G.G.; Li, D.; Hou, C.; Ding, X.; Hou, L.; Lyu, Q.; Yang, Y.; Cooney, A.J.; et al. Offspring production of ovarian organoids derived from spermatogonial stem cells by defined factors with chromatin reorganization. J. Adv. Res. 2021, 33, 81–98. [Google Scholar] [CrossRef]
- Ziloochi, K.M.; Bagher, Z.; Asgari, H.R.; Najafi, M.; Koruji, M.; Mehraein, F. Differentiation of neonate mouse spermatogonial stem cells on three-dimensional agar/polyvinyl alcohol nanofiber scaffold. Syst. Biol. Reprod. Med. 2020, 66, 202–215. [Google Scholar] [CrossRef]
- Khadivi, F.; Koruji, M.; Akbari, M.; Jabari, A.; Talebi, A.; Ashouri, M.S.; Panahi, B.A.; Feizollahi, N.; Nikmahzar, A.; Pourahmadi, M.; et al. Application of platelet-rich plasma (prp) improves self-renewal of human spermatogonial stem cells in two-dimensional and three-dimensional culture systems. Acta Histochem. 2020, 122, 151627. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, M.; Lee, S.W.; Choi, N.Y.; Ham, S.; Lee, H.J.; Ko, K.; Ko, K. Reprogramming of spermatogonial stem cells into pluripotent stem cells in the spheroidal state. Anim. Cells Syst. 2019, 23, 392–398. [Google Scholar] [CrossRef]
- Rahmani, F.; Movahedin, M.; Mazaheri, Z.; Soleimani, M. Transplantation of mouse ipscs into testis of azoospermic mouse model: In Vivo and in vitro study. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1585–1594. [Google Scholar] [CrossRef]
- Gholami, K.; Pourmand, G.; Koruji, M.; Sadighigilani, M.; Navid, S.; Izadyar, F.; Abbasi, M. Efficiency of colony formation and differentiation of human spermatogenic cells in two different culture systems. Reprod. Biol. 2018, 18, 397–403. [Google Scholar] [CrossRef]
- Yi, H.; Xiao, S.; Zhang, Y. Stage-specific approaches promote in vitro induction for spermatogenesis. Vitr. Cell Dev. Biol. Anim. 2018, 54, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Abumadighem, A.; Solomon, R.; Stepanovsky, A.; Kapelushnik, J.; Shi, Q.; Meese, E.; Lunenfeld, E.; Huleihel, M. Development of spermatogenesis in vitro in three-dimensional culture from spermatogonial cells of busulfan-treated immature mice. Int. J. Mol. Sci. 2018, 19, 3804. [Google Scholar] [CrossRef] [PubMed]
- Huleihel, M.; Nourashrafeddin, S.; Plant, T.M. Application of three-dimensional culture systems to study mammalian spermatogenesis, with an emphasis on the rhesus monkey (Macaca mulatta). Asian J. Androl. 2015, 17, 972–980. [Google Scholar] [CrossRef]
- Khajavi, N.; Akbari, M.; Abdolsamadi, H.R.; Abolhassani, F.; Dehpour, A.R.; Koruji, M.; Habibi, R.M. Role of somatic testicular cells during mouse spermatogenesis in three-dimensional collagen gel culture system. Cell J. 2014, 16, 79–90. [Google Scholar] [PubMed]
- Eslahi, N.; Hadjighassem, M.R.; Joghataei, M.T.; Mirzapour, T.; Bakhtiyari, M.; Shakeri, M.; Pirhajati, V.; Shirinbayan, P.; Koruji, M. The effects of poly l-lactic acid nanofiber scaffold on mouse spermatogonial stem cell culture. Int. J. Nanomed. 2013, 8, 4563–4576. [Google Scholar]
- Abofoul-Azab, M.; Lunenfeld, E.; Levitas, E.; Zeadna, A.; Younis, J.S.; Bar-Ami, S.; Huleihel, M. Identification of premeiotic, meiotic, and postmeiotic cells in testicular biopsies without sperm from sertoli cell-only syndrome patients. Int. J. Mol. Sci. 2019, 20, 470. [Google Scholar] [CrossRef]
- Piravar, Z.; Jeddi-Tehrani, M.; Sadeghi, M.R.; Mohazzab, A.; Eidi, A.; Akhondi, M.M. In Vitro culture of human testicular stem cells on feeder-free condition. J. Reprod. Infertil. 2013, 14, 17–22. [Google Scholar]
- Jabari, A.; Sadighi, G.M.; Koruji, M.; Gholami, K.; Mohsenzadeh, M.; Rastegar, T.; Khadivi, F.; Ghanami, G.N.; Nikmahzar, A.; Mojaverrostami, S.; et al. Three-dimensional co-culture of human spermatogonial stem cells with sertoli cells in soft agar culture system supplemented by growth factors and laminin. Acta Histochem. 2020, 122, 151572. [Google Scholar] [CrossRef]
- Pramod, R.K.; Mitra, A. In Vitro culture and characterization of spermatogonial stem cells on sertoli cell feeder layer in goat (Capra hircus). J. Assist. Reprod. Genet. 2014, 31, 993–1001. [Google Scholar] [CrossRef]
- Nasiri, Z.; Hosseini, S.M.; Hajian, M.; Abedi, P.; Bahadorani, M.; Baharvand, H.; Nasr-Esfahani, M.H. Effects of different feeder layers on short-term culture of prepubertal bovine testicular germ cells in-vitro. Theriogenology 2012, 77, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Kala, S.; Kaushik, R.; Singh, K.P.; Kadam, P.H.; Singh, M.K.; Manik, R.S.; Singla, S.K.; Palta, P.; Chauhan, M.S. In Vitro culture and morphological characterization of prepubertal buffalo (Bubalus bubalis) putative spermatogonial stem cell. J. Assist. Reprod. Genet. 2012, 29, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Rastegar, T.; Minaee, M.B.; Habibi, R.M.; Raghardi, K.I.; Amidi, F.; Abolhasani, F.; Barbarestani, M. Improvement of expression of alpha6 and beta1 integrins by the co-culture of adult mouse spermatogonial stem cells with sim mouse embryonic fibroblast cells (sto) and growth factors. Iran. J. Basic. Med. Sci. 2013, 16, 134–139. [Google Scholar] [PubMed]
- Veisi, M.; Mansouri, K.; Assadollahi, V.; Jalili, C.; Pirnia, A.; Salahshoor, M.R.; Hoseinkhani, Z.; Gholami, M.R. Evaluation of co-cultured spermatogonial stem cells encapsulated in alginate hydrogel with sertoli cells and their transplantation into azoospermic mice. Zygote 2022, 30, 344–351. [Google Scholar] [CrossRef]
- Yin, L.; Siracusa, J.S.; Measel, E.; Guan, X.; Edenfield, C.; Liang, S.; Yu, X. High-content image-based single-cell phenotypic analysis for the testicular toxicity prediction induced by bisphenol a and its analogs bisphenol s, bisphenol af, and tetrabromobisphenol a in a three-dimensional testicular cell co-culture model. Toxicol. Sci. 2020, 173, 313–335. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, S.; Ahn, J.; Park, J.; Ryu, B.Y.; Park, J.Y. Membrane-bottomed microwell array added to transwell insert to facilitate non-contact co-culture of spermatogonial stem cell and sto feeder cell. Biofabrication 2020, 12, 45031. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; Bedford, E.; Witherspoon, L.; Willerth, S.M.; Flannigan, R. Using clinically derived human tissue to 3-dimensionally bioprint personalized testicular tubules for in vitro culturing: First report. F S Sci. 2022, 3, 130–139. [Google Scholar] [CrossRef]
- Mahmoud, H. Concise review: Spermatogenesis in an artificial three-dimensional system. Stem Cells 2012, 30, 2355–2360. [Google Scholar] [CrossRef]
- Wong, K.C. 3d-printed patient-specific applications in orthopedics. Orthop. Res. Rev. 2016, 8, 57–66. [Google Scholar] [CrossRef]
- Ding, A.; Jeon, O.; Cleveland, D.; Gasvoda, K.L.; Wells, D.; Lee, S.J.; Alsberg, E. Jammed micro-flake hydrogel for four-dimensional living cell bioprinting. Adv. Mater. 2022, 34, e2109394. [Google Scholar] [CrossRef]
- Gagan, J.; Fraze, C.; Stout, D.A. Three-dimensional stem cell bioprinting. Cell Stem Cells Regen. Med. 2016, 2, 10. [Google Scholar] [CrossRef]
- Angelats, L.D.; Ginestra, P. Cell bioprinting: The 3d-bioplotter case. Materials 2019, 12, 4005. [Google Scholar] [CrossRef] [PubMed]
- Gebeyehu, A.; Surapaneni, S.K.; Huang, J.; Mondal, A.; Wang, V.Z.; Haruna, N.F.; Bagde, A.; Arthur, P.; Kutlehria, S.; Patel, N.; et al. Polysaccharide hydrogel based 3d printed tumor models for chemotherapeutic drug screening. Sci. Rep. 2021, 11, 372. [Google Scholar] [CrossRef]
- Vultur, A.; Schanstra, T.; Herlyn, M. The promise of 3d skin and melanoma cell bioprinting. Melanoma Res. 2016, 26, 205–206. [Google Scholar] [CrossRef]
- Xu, F.; Sridharan, B.; Wang, S.; Gurkan, U.A.; Syverud, B.; Demirci, U. Embryonic stem cell bioprinting for uniform and controlled size embryoid body formation. Biomicrofluidics 2011, 5, 22207. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Y.; Dai, Y.; Yang, L.; Chen, G. Application of 3d bioprinting in urology. Micromachines 2022, 13, 1073. [Google Scholar] [CrossRef]
- Poels, J.; Abou-Ghannam, G.; Decamps, A.; Leyman, M.; Rieux, A.; Wyns, C. Transplantation of testicular tissue in alginate hydrogel loaded with vegf nanoparticles improves spermatogonial recovery. J. Control. Release 2016, 234, 79–89. [Google Scholar] [CrossRef]
- Li, Q.; You, N.; Zhang, M.; Wang, W.; Shi, W.; Li, F. Current progression in establishment and application of three-dimensional printing in urology. Minerva Med. 2022, 113, 1063–1064. [Google Scholar] [CrossRef]
- Chen, M.Y.; Skewes, J.; Desselle, M.; Wong, C.; Woodruff, M.A.; Dasgupta, P.; Rukin, N.J. Current applications of three-dimensional printing in urology. BJU Int. 2020, 125, 17–27. [Google Scholar] [CrossRef]
- Biazar, E.; Najafi, S.M.; Heidari, K.S.; Yazdankhah, M.; Rafiei, A.; Biazar, D. 3d bio-printing technology for body tissues and organs regeneration. J. Med. Eng. Technol. 2018, 42, 187–202. [Google Scholar] [CrossRef]
- Bashiri, Z.; Gholipourmalekabadi, M.; Falak, R.; Amiri, I.; Asgari, H.; Chauhan, N.; Koruji, M. In Vitro production of mouse morphological sperm in artificial testis bioengineered by 3d printing of extracellular matrix. Int. J. Biol. Macromol. 2022, 217, 824–841. [Google Scholar]
- Richer, G.; Hobbs, R.M.; Loveland, K.L.; Goossens, E.; Baert, Y. Long-term maintenance and meiotic entry of early germ cells in murine testicular organoids functionalized by 3d printed scaffolds and air-medium interface cultivation. Front. Physiol. 2021, 12, 757565. [Google Scholar] [CrossRef]
- Horvath-Pereira, B.O.; Almeida, G.; Da, S.J.L.; Do, N.P.; Horvath, P.B.; Fireman, J.; Pereira, M.; Carreira, A.; Miglino, M.A. Biomaterials for testicular bioengineering: How far have we come and where do we have to go? Front. Endocrinol. 2023, 14, 1085872. [Google Scholar]
- Bashiri, Z.; Zahiri, M.; Allahyari, H.; Esmaeilzade, B. Proliferation of human spermatogonial stem cells on optimized pcl/gelatin nanofibrous scaffolds. Andrologia 2022, 54, e14380. [Google Scholar] [CrossRef] [PubMed]
- Koch, F.; Thaden, O.; Conrad, S.; Trondle, K.; Finkenzeller, G.; Zengerle, R.; Kartmann, S.; Zimmermann, S.; Koltay, P. Mechanical properties of polycaprolactone (pcl) scaffolds for hybrid 3d-bioprinting with alginate-gelatin hydrogel. J. Mech. Behav. Biomed. Mater. 2022, 130, 105219. [Google Scholar]
- Giuseppe, M.D.; Law, N.; Webb, B.; Macrae, A.R.; Liew, L.J.; Sercombe, T.B.; Dilley, R.J.; Doyle, B.J. Mechanical behaviour of alginate-gelatin hydrogels for 3d bioprinting. J. Mech. Behav. Biomed. Mater. 2018, 79, 150–157. [Google Scholar] [CrossRef]
- Xu, E.; Niu, R.; Lao, J.; Zhang, S.; Li, J.; Zhu, Y.; Shi, H.; Zhu, Q.; Chen, Y.; Jiang, Y.; et al. Tissue-like cultured fish fillets through a synthetic food pipeline. NPJ Sci. Food 2023, 7, 17. [Google Scholar]
- Majidi, G.N.; Movahedin, M.; Mazaheri, Z. Three-dimensional culture of mouse spermatogonial stem cells using a decellularised testicular scaffold. Cell J. 2020, 21, 410–418. [Google Scholar]
- Schmidt, J.A.; Abramowitz, L.K.; Kubota, H.; Wu, X.; Niu, Z.; Avarbock, M.R.; Tobias, J.W.; Bartolomei, M.S.; Brinster, R.L. In Vivo and in vitro aging is detrimental to mouse spermatogonial stem cell function. Biol. Reprod. 2011, 84, 698–706. [Google Scholar] [CrossRef]
- Niu, B.; Wu, J.; Mu, H.; Li, B.; Wu, C.; He, X.; Bai, C.; Li, G.; Hua, J. Mir-204 regulates the proliferation of dairy goat spermatogonial stem cells via targeting to sirt1. Rejuvenation Res. 2016, 19, 120–130. [Google Scholar] [CrossRef]
- Rahbar, M.; Asadpour, R.; Azami, M.; Mazaheri, Z.; Hamali, H. Improving the process of spermatogenesis in azoospermic mice using spermatogonial stem cells co-cultured with epididymosomes in three-dimensional culture system. Life Sci. 2022, 310, 121057. [Google Scholar] [PubMed]
- Kofman, A.E.; Mcgraw, M.R.; Payne, C.J. Rapamycin increases oxidative stress response gene expression in adult stem cells. Aging 2012, 4, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V. Aging and immortality: Quasi-programmed senescence and its pharmacologic inhibition. Cell Cycle 2006, 5, 2087–2102. [Google Scholar] [CrossRef]
- Kofman, A.E.; Huszar, J.M.; Payne, C.J. Transcriptional analysis of histone deacetylase family members reveal similarities between differentiating and aging spermatogonial stem cells. Stem Cell Rev. Rep. 2013, 9, 59–64. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Kang, K.; Liu, S.; Ma, Y.; Yu, M.; Zhao, X. Recent Progress of In Vitro 3D Culture of Male Germ Stem Cells. J. Funct. Biomater. 2023, 14, 543. https://doi.org/10.3390/jfb14110543
Wu J, Kang K, Liu S, Ma Y, Yu M, Zhao X. Recent Progress of In Vitro 3D Culture of Male Germ Stem Cells. Journal of Functional Biomaterials. 2023; 14(11):543. https://doi.org/10.3390/jfb14110543
Chicago/Turabian StyleWu, Jiang, Kai Kang, Siqi Liu, Yaodan Ma, Meng Yu, and Xin Zhao. 2023. "Recent Progress of In Vitro 3D Culture of Male Germ Stem Cells" Journal of Functional Biomaterials 14, no. 11: 543. https://doi.org/10.3390/jfb14110543