

Increased UHMWPE Particle-Induced Osteolysis in Fetuin-A-Deficient Mice

 ,
,  , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethics
2.2. Ultra-High Molecular Weight Polyethylene UHMWPE Particles
2.3. Animal Surgery
2.4. Microcomputed Tomography
2.5. Bone Histomorphometry
2.6. Analyses of the Parameters of Bone Metabolism
2.7. Statistical Analysis
3. Results
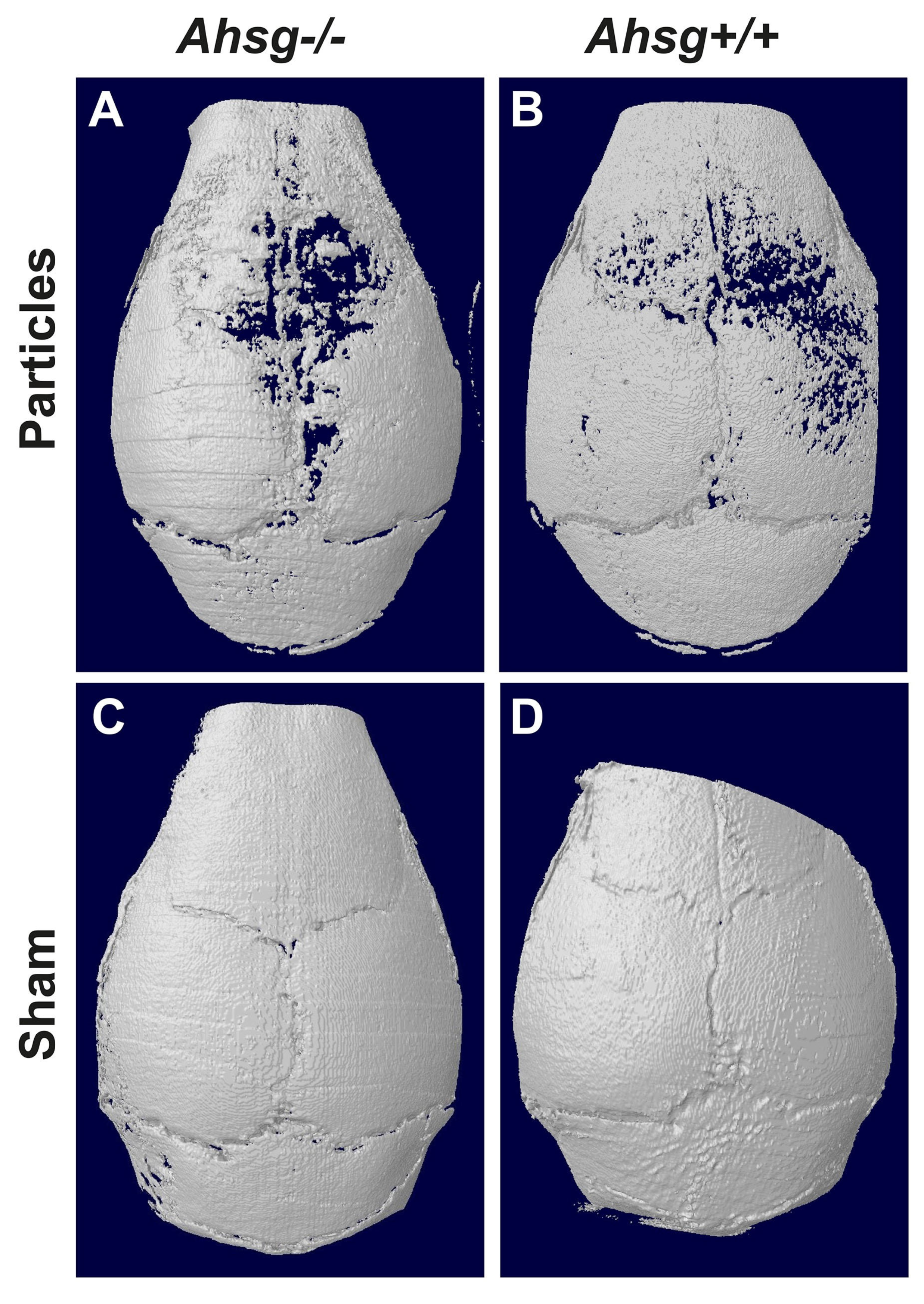
3.1. Micro Computed Tomography
3.2. Bone Histomorphometry
3.3. Parameters of Bone Metabolism
4. Discussion
4.1. In Absence of Fetuin-A, the Particle-Induced Inflammatory Response and Osteolysis Is Increased
4.2. Comparing the Phenotypes before Particle Treatment: Fetuin-A-Deficient Mice Presented Differences Regarding Bone Micro-Structure in µ-CT and Serum Analytes
4.3. Potential Mechanisms of Fetuin-A in Particle Treatment: Fetuin-A Attenuates UHMWPE Particle-Induced Local Inflammation by Aiding the Removal of Calcified Cellular and Tissue Debris
4.4. Correlation between Fetuin-A and Bone Metabolism in Humans: Fetuin-A Is Slightly Possitivly Associated with Areal Bone Mineral Density in Older Adults
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sadoghi, P.; Liebensteiner, M.; Agreiter, M.; Leithner, A.; Bohler, N.; Labek, G. Revision surgery after total joint arthroplasty: A complication-based analysis using worldwide arthroplasty registers. J. Arthroplast. 2013, 28, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Gallo, J.; Goodman, S.B.; Konttinen, Y.T.; Raska, M. Particle disease: Biologic mechanisms of periprosthetic osteolysis in total hip arthroplasty. Innate. Immun. 2013, 19, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, S.B.; Gallo, J. Periprosthetic Osteolysis: Mechanisms, Prevention and Treatment. J. Clin. Med. 2019, 8, 2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, A.; Bonfield, W.; Kadoya, Y.; Yamac, T.; Freeman, M.A.; Scott, G.; Revell, P.A. The size and shape of particulate polyethylene wear debris in total joint replacements. Proc. Inst. Mech. Eng. H 1997, 211, 11–15. [Google Scholar] [CrossRef]
- Matthews, J.B.; Besong, A.A.; Green, T.R.; Stone, M.H.; Wroblewski, B.M.; Fisher, J.; Ingham, E. Evaluation of the response of primary human peripheral blood mononuclear phagocytes to challenge with in vitro generated clinically relevant UHMWPE particles of known size and dose. J. Biomed. Mater. Res. 2000, 52, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Datta, H.K.; Ng, W.F.; Walker, J.A.; Tuck, S.P.; Varanasi, S.S. The cell biology of bone metabolism. J. Clin. Pathol. 2008, 61, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Drees, P.; Eckardt, A.; Gay, R.E.; Gay, S.; Huber, L.C. Mechanisms of disease: Molecular insights into aseptic loosening of orthopedic implants. Nat. Clin. Pract. Rheumatol. 2007, 3, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Revell, P.A. The combined role of wear particles, macrophages and lymphocytes in the loosening of total joint prostheses. J. R. Soc. Interface 2008, 5, 1263–1278. [Google Scholar] [CrossRef] [Green Version]
- Khosla, S. Minireview: The OPG/RANKL/RANK system. Endocrinology 2001, 142, 5050–5055. [Google Scholar] [CrossRef]
- Trouvin, A.P.; Goeb, V. Receptor activator of nuclear factor-kappaB ligand and osteoprotegerin: Maintaining the balance to prevent bone loss. Clin. Interv. Aging 2010, 5, 345–354. [Google Scholar] [CrossRef]
- Ingham, E.; Fisher, J. Biological reactions to wear debris in total joint replacement. Proc. Inst. Mech. Eng. H 2000, 214, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Baumann, B.; Seufert, J.; Jakob, F.; Noth, U.; Rolf, O.; Eulert, J.; Rader, C.P. Activation of NF-kappaB signalling and TNFalpha-expression in THP-1 macrophages by TiAlV- and polyethylene-wear particles. J. Orthop. Res. 2005, 23, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Kitaura, H.; Zhou, P.; Ross, F.P.; Teitelbaum, S.L. IL-1 mediates TNF-induced osteoclastogenesis. J. Clin. Investig. 2005, 115, 282–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickson, I.R.; Poole, A.R.; Veis, A. Localisation of plasma alpha2HS glycoprotein in mineralising human bone. Nature 1975, 256, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Triffitt, J.T.; Gebauer, U.; Ashton, B.A.; Owen, M.E.; Reynolds, J.J. Origin of plasma alpha2HS-glycoprotein and its accumulation in bone. Nature 1976, 262, 226–227. [Google Scholar] [CrossRef]
- Schäfer, C.; Heiss, A.; Schwarz, A.; Westenfeld, R.; Ketteler, M.; Floege, J.; Müller-Esterl, W.; Schinke, T.; Jahnen-Dechent, W. The serum protein α2–Heremans-Schmid glycoprotein/fetuin-A is a systemically acting inhibitor of ectopic calcification. J. Clin. Investig. 2003, 112, 357–366. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Heiss, A.; Schäfer, C.; Ketteler, M. Fetuin-A regulation of calcified matrix metabolism. Circ. Res. 2011, 108, 1494–1509. [Google Scholar] [CrossRef] [Green Version]
- Schinke, T.; Amendt, C.; Trindl, A.; Poschke, O.; Muller-Esterl, W.; Jahnen-Dechent, W. The serum protein alpha2-HS glycoprotein/fetuin inhibits apatite formation in vitro and in mineralizing calvaria cells. A possible role in mineralization and calcium homeostasis. J. Biol. Chem. 1996, 271, 20789–20796. [Google Scholar] [CrossRef] [Green Version]
- Heiss, A.; Eckert, T.; Aretz, A.; Richtering, W.; Van Dorp, W.; Schäfer, C.; Jahnen-Dechent, W. Hierarchical role of fetuin-A and acidic serum proteins in the formation and stabilization of calcium phosphate particles. J. Biol. Chem. 2008, 283, 14815–14825. [Google Scholar] [CrossRef] [Green Version]
- Rudloff, S.; Janot, M.; Rodriguez, S.; Dessalle, K.; Jahnen-Dechent, W.; Huynh-Do, U. Fetuin-A is a HIF target that safeguards tissue integrity during hypoxic stress. Nat. Commun. 2021, 12, 549. [Google Scholar] [CrossRef]
- Jirak, P.; Stechemesser, L.; More, E.; Franzen, M.; Topf, A.; Mirna, M.; Paar, V.; Pistulli, R.; Kretzschmar, D.; Wernly, B.; et al. Clinical implications of fetuin-A. Adv. Clin. Chem. 2019, 89, 79–130. [Google Scholar] [CrossRef]
- Wang, H.; Sama, A.E. Anti-inflammatory role of fetuin-A in injury and infection. Curr. Mol. Med. 2012, 12, 625–633. [Google Scholar] [CrossRef] [Green Version]
- Dziegielewska, K.M.; Andersen, N.A.; Saunders, N.R. Modification of macrophage response to lipopolysaccharide by fetuin. Immunol. Lett. 1998, 60, 31–35. [Google Scholar] [CrossRef]
- Ombrellino, M.; Wang, H.; Yang, H.; Zhang, M.; Vishnubhakat, J.; Frazier, A.; Scher, L.A.; Friedman, S.G.; Tracey, K.J. Fetuin, a negative acute phase protein, attenuates TNF synthesis and the innate inflammatory response to carrageenan. Shock 2001, 15, 181–185. [Google Scholar] [CrossRef]
- Pappa, E.; Perrea, D.S.; Pneumaticos, S.; Nikolaou, V.S. Role of fetuin A in the diagnosis and treatment of joint arthritis. World J. Orthop. 2017, 8, 461–464. [Google Scholar] [CrossRef]
- Jablonski, H.; Polan, C.; Wedemeyer, C.; Hilken, G.; Schlepper, R.; Bachmann, H.S.; Grabellus, F.; Dudda, M.; Jäger, M.; Kauther, M.D. A single intraperitoneal injection of bovine fetuin-A attenuates bone resorption in a murine calvarial model of particle-induced osteolysis. Bone 2017, 105, 262–268. [Google Scholar] [CrossRef]
- Merkel, K.D.; Erdmann, J.M.; McHugh, K.P.; Abu-Amer, Y.; Ross, F.P.; Teitelbaum, S.L. Tumor necrosis factor-alpha mediates orthopedic implant osteolysis. Am. J. Pathol. 1999, 154, 203–210. [Google Scholar] [CrossRef]
- Wedemeyer, C.; Neuerburg, C.; Pfeiffer, A.; Heckelei, A.; Bylski, D.; von Knoch, F.; Schinke, T.; Hilken, G.; Gosheger, G.; von Knoch, M.; et al. Polyethylene particle-induced bone resorption in alpha-calcitonin gene-related peptide-deficient mice. J. Bone Miner. Res. 2007, 22, 1011–1019. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Schinke, T.; Trindl, A.; Müller-Esterl, W.; Sablitzky, F.; Kaiser, S.; Blessing, M. Cloning and targeted deletion of the mouse fetuin gene. J. Biol. Chem. 1997, 272, 31496–31503. [Google Scholar] [CrossRef] [Green Version]
- Westenfeld, R.; Schäfer, C.; Smeets, R.; Brandenburg, V.M.; Floege, J.; Ketteler, M.; Jahnen-Dechent, W. Fetuin-A (AHSG) prevents extraosseous calcification induced by uraemia and phosphate challenge in mice. Nephrol. Dial. Transpl. 2007, 22, 1537–1546. [Google Scholar] [CrossRef]
- Wedemeyer, C.; von Knoch, F.; Pingsmann, A.; Hilken, G.; Sprecher, C.; Saxler, G.; Henschke, F.; Loer, F.; von Knoch, M. Stimulation of bone formation by zoledronic acid in particle-induced osteolysis. Biomaterials 2005, 26, 3719–3725. [Google Scholar] [CrossRef]
- Kauther, M.D.; Bachmann, H.S.; Neuerburg, L.; Broecker-Preuss, M.; Hilken, G.; Grabellus, F.; Koehler, G.; von Knoch, M.; Wedemeyer, C. Calcitonin substitution in calcitonin deficiency reduces particle-induced osteolysis. BMC Musculoskelet Disord 2011, 12, 186. [Google Scholar] [CrossRef] [Green Version]
- Parfitt, A.M. Bone histomorphometry: Proposed system for standardization of nomenclature, symbols, and units. Calcif. Tissue Int. 1988, 42, 284–286. [Google Scholar] [CrossRef]
- Wang, H.C.; Zhang, M.H.; Soda, K.; Sama, A.; Tracey, K.J. Fetuin protects the fetus from TNF. Lancet 1997, 350, 861–862. [Google Scholar] [CrossRef]
- Li, W.; Zhu, S.; Li, J.H.; Huang, Y.; Zhou, R.R.; Fan, X.G.; Yang, H.A.; Gong, X.; Eissa, N.T.; Jahnen-Dechent, W.; et al. A Hepatic Protein, Fetuin-A, Occupies a Protective Role in Lethal Systemic Inflammation. PLoS ONE 2011, 6, e16945. [Google Scholar] [CrossRef] [Green Version]
- Köppert, S.; Büscher, A.; Babler, A.; Ghallab, A.; Buhl, E.M.; Latz, E.; Hengstler, J.G.; Smith, E.R.; Jahnen-Dechent, W. Cellular Clearance and Biological Activity of Calciprotein Particles Depend on Their Maturation State and Crystallinity. Front. Immunol. 2018, 9, 1991. [Google Scholar] [CrossRef]
- Köppert, S.; Ghallab, A.; Peglow, S.; Winkler, C.F.; Graeber, S.; Buscher, A.; Hengstler, J.G.; Jahnen-Dechent, W. Live Imaging of Calciprotein Particle Clearance and Receptor Mediated Uptake: Role of Calciprotein Monomers. Front. Cell Dev. Biol. 2021, 9, 633925. [Google Scholar] [CrossRef]
- Landgraeber, S.; Toetsch, M.; Wedemeyer, C.; Saxler, G.; Tsokos, M.; von Knoch, F.; Neuhauser, M.; Loer, F.; von Knoch, M. Over-expression of p53/BAK in aseptic loosening after total hip replacement. Biomaterials 2006, 27, 3010–3020. [Google Scholar] [CrossRef]
- Landgraeber, S.; von Knoch, M.; Loer, F.; Wegner, A.; Tsokos, M.; Hussmann, B.; Totsch, M. Extrinsic and intrinsic pathways of apoptosis in aseptic loosening after total hip replacement. Biomaterials 2008, 29, 3444–3450. [Google Scholar] [CrossRef]
- Jersmann, H.P.A.; Dransfield, I.; Hart, S.P. Fetuin/alpha(2)-HS glycoprotein enhances phagocytosis of apoptotic cells and macropinocytosis by human macrophages. Clin. Sci. 2003, 105, 273–278. [Google Scholar] [CrossRef]
- Brylka, L.J.; Köppert, S.; Babler, A.; Kratz, B.; Denecke, B.; Yorgan, T.A.; Etich, J.; Costa, I.G.; Brachvogel, B.; Boor, P.; et al. Post-weaning epiphysiolysis causes distal femur dysplasia and foreshortened hindlimbs in fetuin-A-deficient mice. PLoS ONE 2017, 12, e0187030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seto, J.; Busse, B.; Gupta, H.S.; Schäfer, C.; Krauss, S.; Dunlop, J.W.C.; Masic, A.; Kerschnitzki, M.; Zaslansky, P.; Boesecke, P.; et al. Accelerated Growth Plate Mineralization and Foreshortened Proximal Limb Bones in Fetuin-A Knockout Mice. PLoS ONE 2012, 7, e47338. [Google Scholar] [CrossRef]
- Fink, H.A.; Bůžková, P.; Garimella, P.S.; Mukamal, K.J.; Cauley, J.A.; Kizer, J.R.; Barzilay, J.I.; Jalal, D.I.; Ix, J.H. Association of Fetuin-A With Incident Fractures in Community-Dwelling Older Adults: The Cardiovascular Health Study. J. Bone Min. Res. 2015, 30, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Steffen, C.J.; Herlyn, P.K.E.; Cornelius, N.; Mittlmeier, T.; Fischer, D.C. Evaluation of fetuin-A as a predictor of outcome after surgery for osteoporotic fracture of the proximal femur. Arch. Orthop. Trauma. Surg. 2020, 140, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Babler, A.; Schmitz, C.; Buescher, A.; Herrmann, M.; Gremse, F.; Gorgels, T.; Floege, J.; Jahnen-Dechent, W. Microvasculopathy and soft tissue calcification in mice are governed by fetuin-A, magnesium and pyrophosphate. PLoS ONE 2020, 15, e0228938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, M.; Babler, A.; Moshkova, I.; Gremse, F.; Kiessling, F.; Kusebauch, U.; Nelea, V.; Kramann, R.; Moritz, R.L.; McKee, M.D.; et al. Lumenal calcification and microvasculopathy in fetuin-A-deficient mice lead to multiple organ morbidity. PLoS ONE 2020, 15, e0228503. [Google Scholar] [CrossRef] [Green Version]
- Langlois, J.; Hamadouche, M. New animal models of wear-particle osteolysis. Int. Orthop. 2011, 35, 245–251. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bone Volume | Ahsg−/− | Ahsg+/+ | ||
|---|---|---|---|---|
| [mm3] | Particles | Sham | Particles | Sham |
| Median (IQR) | 0.50 (0.02) | 0.53 (0.02) | 0.52 (0.01) | 0.54 (0.03) |
| Mean ± SD | 0.50 ± 0.01 | 0.53 ± 0.01 | 0.52 ± 0.01 | 0.54 ± 0.02 |
| Bone Volume/ Tissue Volume [×10−2] | Ahsg−/− | Ahsg+/+ | ||
|---|---|---|---|---|
| Particles | Sham | Particles | Sham | |
| Median (IQR) | 5.69 (0.18) | 6.02 (0.05) | 6.01 (0.15) | 6.19 (0.31) |
| Mean ± SD | 5.71 ± 0.09 | 6.03 ± 0.03 | 6.00 ± 0.09 | 6.25 ± 0.17 |
| Osteoclast Number | Ahsg−/− | Ahsg+/+ | ||
|---|---|---|---|---|
| Particles | Sham | Particles | Sham | |
| Median (IQR) | 14.5 (10.1) | 6.3 (2.9) | 16.7 (15.5) | 9.7 (8.4) |
| Mean ± SD | 13.5 ± 5.3 | 6.4 ± 1.5 | 15.5 ± 8.3 | 9.7 ± 5.3 |
| Eroded Surface Area [mm2] | Ahsg−/− | Ahsg+/+ | ||
|---|---|---|---|---|
| Particles | Sham | Particles | Sham | |
| Median (IQR) | 0.32 (0.02) | 0.10 (0.14) | 0.12 (0.10) | 0.05 (0.04) |
| Mean ± SD | 0.32 ± 0.01 | 0.11 ± 0.08 | 0.14 ± 0.06 | 0.06 ± 0.02 |
| Parameter | Day 0 (Baseline) (n = 10 per Group) | Day 14 (n = 5 per Group) | ||||
|---|---|---|---|---|---|---|
| Ahsg−/− | Ahsg+/+ | Ahsg−/− | Ahsg+/+ | |||
| Particles | Sham | Particles | Sham | |||
| DPD/Creatinin | 12.4 (2.1) 12.7 ± 1.5 | 15.5 (4.8) 16.4 ± 3.5 | 17.9 (6.1) 16.7± 3.5 | 16.1 15.9 ± 1.2 | 22.5 (6.7) 23.8 ± 3.5 | 19.71 (4.21) 19.0 ± 2.2 |
| RANKL | 86.5 (60.3) | 95.6 (43.9) | 122.6 | 96.2 (15.1) | 128.6 (69.9) | 146.4 (32.6) |
| 96.1 ± 31.0 | 95.4 ± 27.5 | 122.2 ± 3.2 | 94.51 ± 7.9 | 110.1 ± 47.0 | 138.0 ± 18.6 | |
| Osteo-protegerin | 2021 (29) 2046 ± 89 | 1760 (272) 1685 ± 259 | 2485 2426 ± 438 | 1950 (467) 1884 ± 275 | 1593 (316) 1654 ± 199 | 1653 (309) 1632 ± 189 |
| Osteocalcin | 88.7 (54.1) | 116.1 (18.8) | 90.0 | 64.7 (46.4) | 120.9 (50.9) | 64.6 (35.0) |
| 82.8 ± 29.3 | 115.7 ± 12.0 | 104.0 ± 51.5 | 71.5 ± 29.5 | 105.7 ± 27.1 | 63.1 ± 17.7 | |
| Alkaline Phosphatase | 195.0 (56.1) 186.9 ± 30.2 | 339.6 (46.3) 340.4 ± 25.4 | 100.8 101.5 ± 50.8 | 98.3 (18.5) 94.6 ± 10.7 | 266.8 (118.0) 263.5 ± 72.1 | 227.6 (109.8) 253.3 ± 58.9 |
| Calcium | 1.69 (0.31) 1.72 ± 0.17 | 2.27 (0.20) 2.26 ± 0.12 | 2.95 2.98 ± 0.6 | 3.0 (0.6) 2.9 ± 0.3 | 2.8 (0.7) 3.0 ± 0.4 | 2.9 (0.3) 2.9 ± 0.2 |
| Phosphate | 5.0 (0.6) 5.0 ± 0.6 | 5.1 (1.5) 5.2 ± 0.8 | 12.1 12.2 ± 1.1 | 11.6 (6.3) 12.4 ± 3.4 | 11.6 (2.6) 12.1 ± 1.6 | 12.0 (2.2) 12.5 ± 1.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polan, C.; Brenner, C.; Herten, M.; Hilken, G.; Grabellus, F.; Meyer, H.-L.; Burggraf, M.; Dudda, M.; Jahnen-Dechent, W.; Wedemeyer, C.; et al. Increased UHMWPE Particle-Induced Osteolysis in Fetuin-A-Deficient Mice. J. Funct. Biomater. 2023, 14, 30. https://doi.org/10.3390/jfb14010030
Polan C, Brenner C, Herten M, Hilken G, Grabellus F, Meyer H-L, Burggraf M, Dudda M, Jahnen-Dechent W, Wedemeyer C, et al. Increased UHMWPE Particle-Induced Osteolysis in Fetuin-A-Deficient Mice. Journal of Functional Biomaterials. 2023; 14(1):30. https://doi.org/10.3390/jfb14010030
Chicago/Turabian StylePolan, Christina, Christina Brenner, Monika Herten, Gero Hilken, Florian Grabellus, Heinz-Lothar Meyer, Manuel Burggraf, Marcel Dudda, Willi Jahnen-Dechent, Christian Wedemeyer, and et al. 2023. "Increased UHMWPE Particle-Induced Osteolysis in Fetuin-A-Deficient Mice" Journal of Functional Biomaterials 14, no. 1: 30. https://doi.org/10.3390/jfb14010030