
Performance of Colombian Silk Fibroin Hydrogels for Hyaline Cartilage Tissue Engineering
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtention of Sterile Silk Fibroin Solution
2.2. Fabrication of Silk Fibroin Hydrogels
2.3. Scanning Electron Microscopy (SEM)
2.4. Attenuated Total Reflectance Fourier Transform Infrared (ATR-FTIR)
2.5. Degradation of Silk Fibroin Hydrogels by Serine Proteases
2.6. Culture of hMSCs
2.7. Measurement of Cell Metabolic Activity
2.8. Chondrogenic Differentiation of hMSCs in Silk Fibroin Hydrogels
2.9. Statistical Analysis
2.10. Ethics Approval
3. Results and Discussion
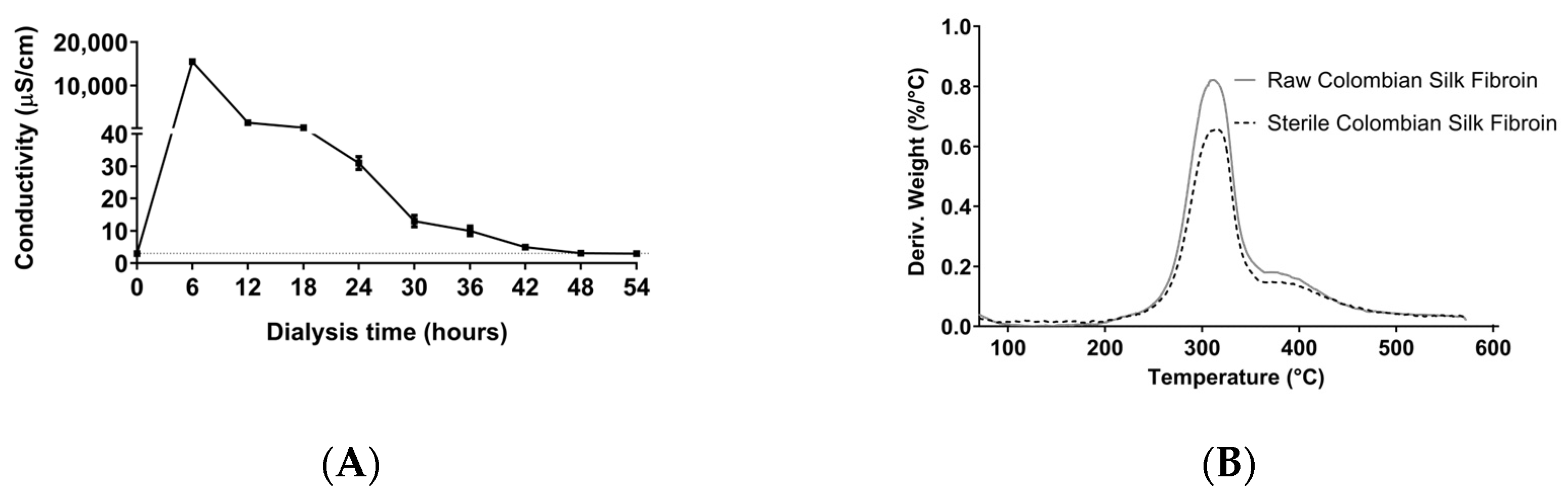
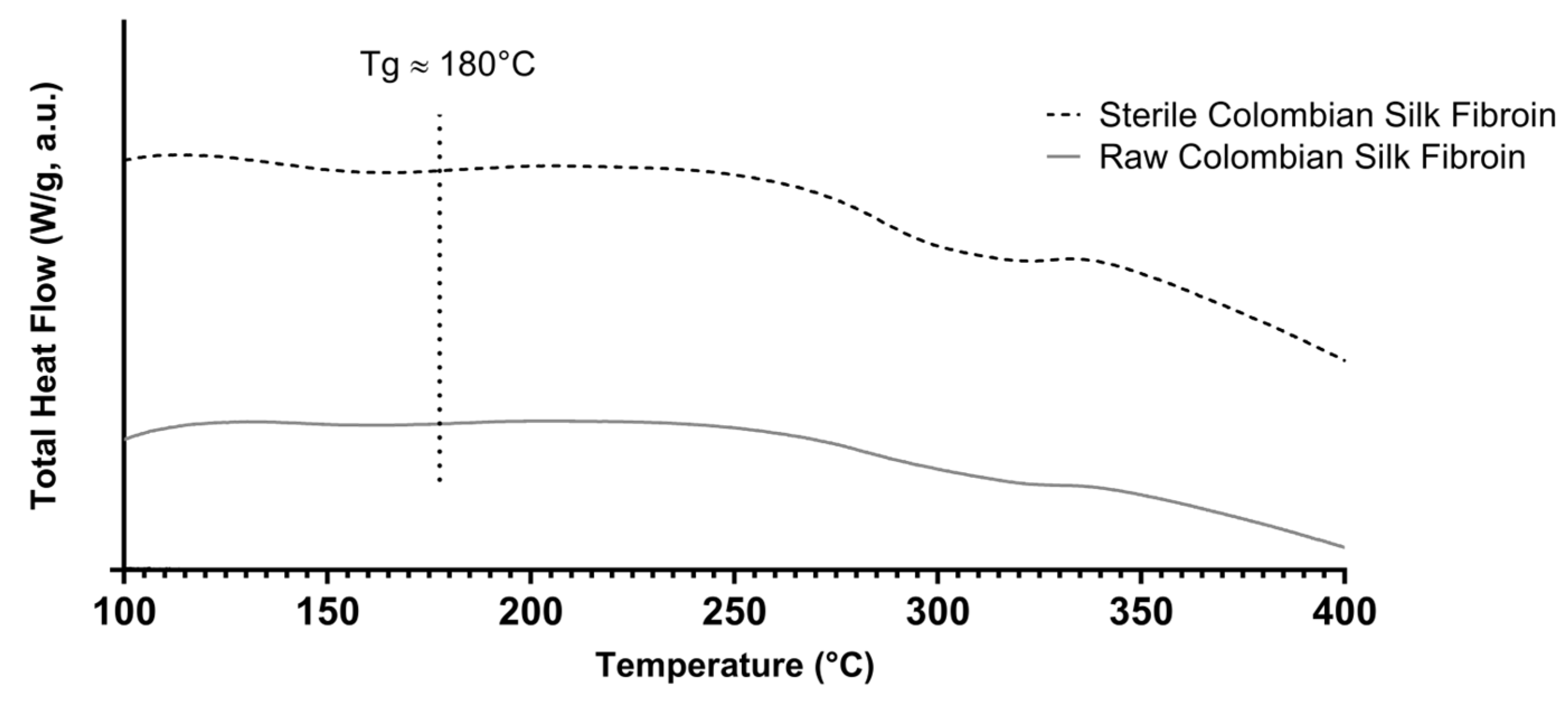
3.1. Obtention of Colombian Silk Fibroin Solution
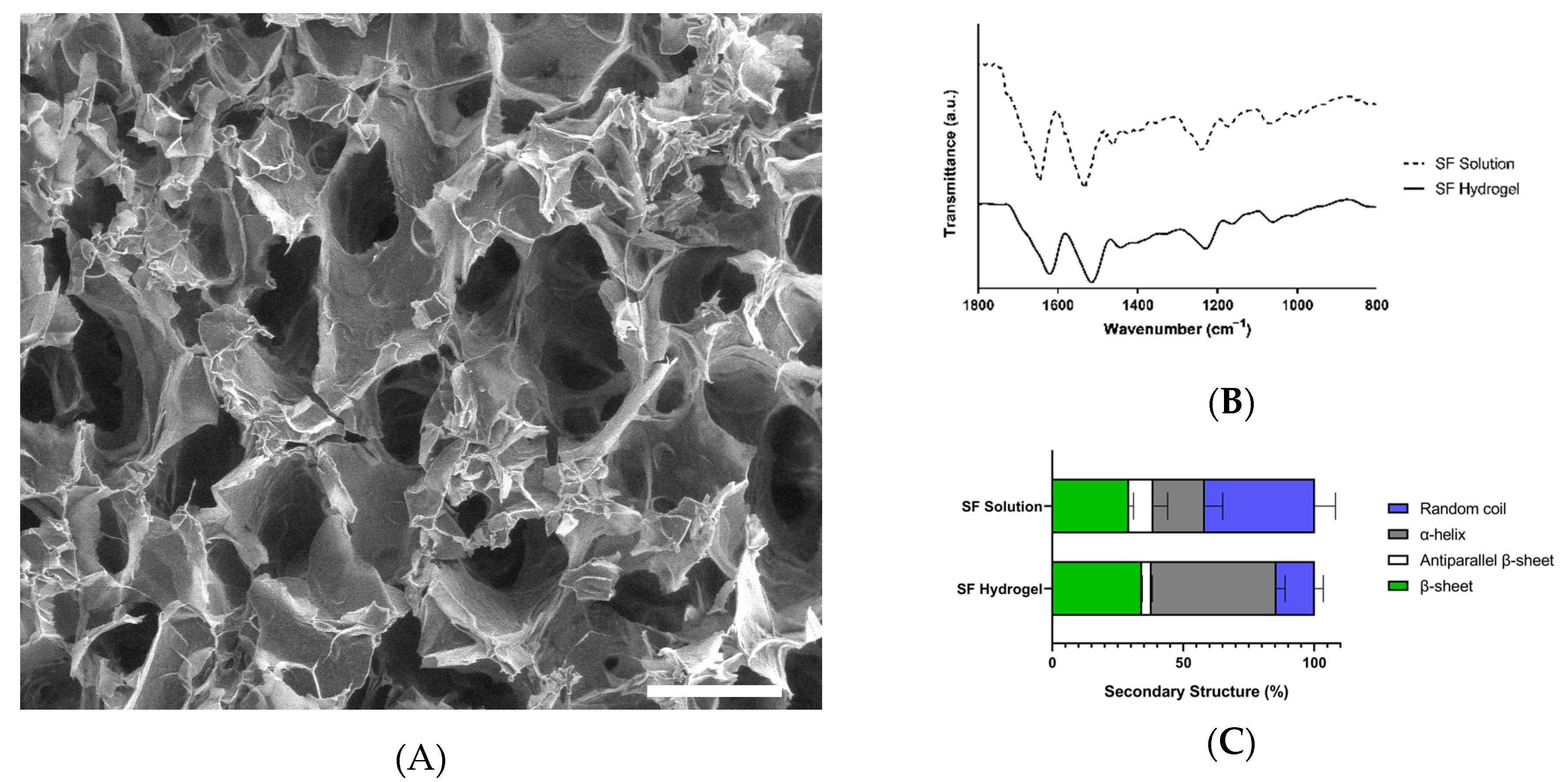
3.2. Characterization of Colombian Silk Fibroin Hydrogels
3.3. Biodegradation of Colombian Silk Fibroin Hydrogels
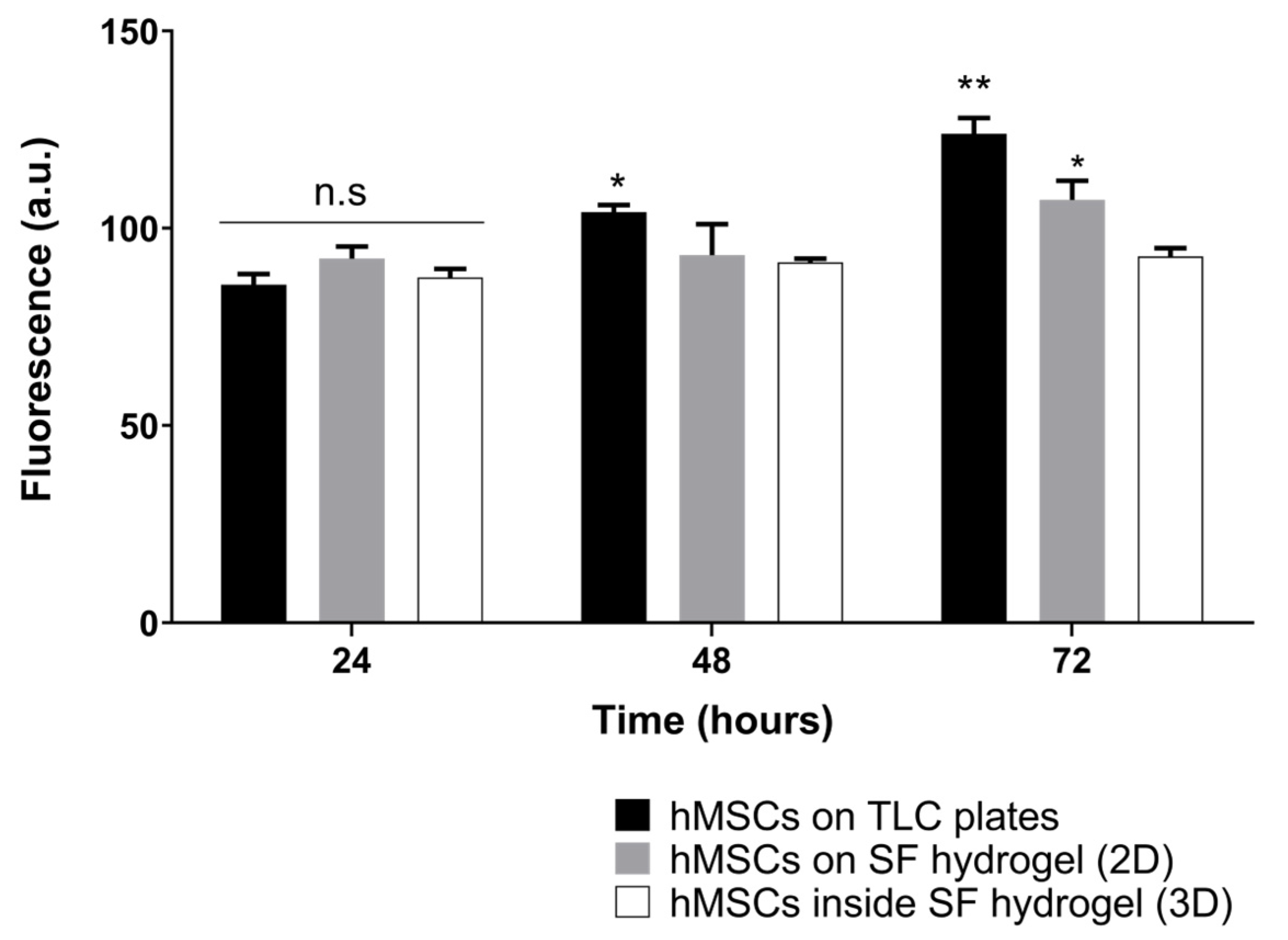
3.4. Cell Metabolic Activity in Colombian Silk Fibroin Hydrogels
3.5. Chondrogenesis in Colombian Silk Fibroin Hydrogels
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Han, L.; Grodzinsky, A.J.; Ortiz, C. Nanomechanics of the Cartilage Extracellular Matrix. Annu. Rev. Mater. Res. 2011, 41, 133–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Windt, T.S.; Hendriks, J.A.A.; Zhao, X.; Vonk, L.A.; Creemers, L.B.; Dhert, W.J.A.; Randolph, M.A.; Saris, D.B.F. Concise Review: Unraveling Stem Cell Cocultures in Regenerative Medicine: Which Cell Interactions Steer Cartilage Regeneration and How? Stem Cells Transl. Med. 2014, 3, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Bannuru, R.R.; Osani, M.C.; Vaysbrot, E.E.; Arden, N.K.; Bennell, K.; Bierma-Zeinstra, S.M.A.; Kraus, V.B.; Lohmander, L.S.; Abbott, J.H.; Bhandari, M.; et al. OARSI guidelines for the non-surgical management of knee, hip, and polyarticular osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1578–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morouço, P.; Fernandes, C.; Santos-Rocha, R. Osteoarthritis, Exercise, and Tissue Engineering: A Stimulating Triad for Health Professionals. J. Aging Res. 2019, 2019, 1935806. [Google Scholar] [CrossRef] [PubMed]
- Martín, A.R.; Patel, J.M.; Zlotnick, H.M.; Carey, J.L.; Mauck, R.L. Emerging therapies for cartilage regeneration in currently excluded ‘red knee’ populations. NPJ Regen. Med. 2019, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Zuluaga-Vélez, A.; Quintero-Martinez, A.; Orozco, L.M.; Sepúlveda-Arias, J.C. Silk fibroin nanocomposites as tissue engineering scaffolds—A systematic review. Biomed. Pharmacother. 2021, 141, 111924. [Google Scholar] [CrossRef]
- Ansari, M.; Eshghanmalek, M. Biomaterials for repair and regeneration of the cartilage tissue. Bio-Design Manuf. 2019, 2, 41–49. [Google Scholar] [CrossRef]
- Huey, D.J.; Hu, J.C.; Athanasiou, K.A. Unlike Bone, Cartilage Regeneration Remains Elusive. Science 2012, 338, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Hunziker, E.B.; Lippuner, K.; Keel, M.J.B.; Shintani, N. An educational review of cartilage repair: Precepts & practice—Myths & misconceptions—Progress & prospects. Osteoarthr. Cartil. 2015, 23, 334–350. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, S.; Kundu, S.C. Silk protein-based hydrogels: Promising advanced materials for biomedical applications. Acta Biomater. 2016, 31, 17–32. [Google Scholar] [CrossRef]
- Kundu, B.; Rajkhowa, R.; Kundu, S.C.; Wang, X. Silk fibroin biomaterials for tissue regenerations. Adv. Drug Deliv. Rev. 2013, 65, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.P.; Nguyen, Q.V.; Nguyen, V.-H.; Le, T.-H.; Huynh, V.Q.N.; Vo, D.-V.N.; Trinh, Q.T.; Kim, S.Y.; Le, Q. Van Silk Fibroin-Based Biomaterials for Biomedical Applications: A Review. Polymers 2019, 11, 1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farokhi, M.; Aleemardani, M.; Solouk, A.; Mirzadeh, H.; Teuschl, A.H.; Redl, H. Crosslinking strategies for silk fibroin hydrogels: Promising biomedical materials. Biomed. Mater. 2021, 16, 022004. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Davoudi, Z.; Xing, X.; Yu, X.; Cheng, X.; Li, Z.; Deng, H.; Wang, Q. Advanced Silk Fibroin Biomaterials for Cartilage Regeneration. ACS Biomater. Sci. Eng. 2018, 4, 2704–2715. [Google Scholar] [CrossRef] [PubMed]
- Farokhi, M.; Mottaghitalab, F.; Fatahi, Y.; Saeb, M.R.; Zarrintaj, P.; Kundu, S.C.; Khademhosseini, A. Silk fibroin scaffolds for common cartilage injuries: Possibilities for future clinical applications. Eur. Polym. J. 2019, 115, 251–267. [Google Scholar] [CrossRef]
- Zhou, Z.; Cui, J.; Wu, S.; Geng, Z.; Su, J. Silk fibroin-based biomaterials for cartilage/osteochondral repair. Theranostics 2022, 12, 5103–5124. [Google Scholar] [CrossRef] [PubMed]
- Bucciarelli, A.; Motta, A. Use of Bombyx mori silk fibroin in tissue engineering: From cocoons to medical devices, challenges, and future perspectives. Biomater. Adv. 2022, 139, 212982. [Google Scholar] [CrossRef]
- Pérez-Silos, V.; Moncada-Saucedo, N.K.; Peña-Martínez, V.; Lara-Arias, J.; Marino-Martínez, I.A.; Camacho, A.; Romero-Díaz, V.J.; Lara Banda, M.; García-Ruiz, A.; Soto-Dominguez, A.; et al. A Cellularized Biphasic Implant Based on a Bioactive Silk Fibroin Promotes Integration and Tissue Organization during Osteochondral Defect Repair in a Porcine Model. Int. J. Mol. Sci. 2019, 20, 5145. [Google Scholar] [CrossRef] [Green Version]
- Zuluaga-Vélez, A.; Cómbita-Merchán, D.F.; Buitrago-Sierra, R.; Santa, J.F.; Aguilar-Fernández, E.; Sepúlveda-Arias, J.C. Silk fibroin hydrogels from the Colombian silkworm Bombyx mori L: Evaluation of physicochemical properties. PLoS ONE 2019, 14, e0213303. [Google Scholar] [CrossRef]
- Rockwood, D.N.; Preda, R.C.; Yücel, T.; Wang, X.; Lovett, M.L.; Kaplan, D.L. Materials fabrication from Bombyx mori silk fibroin. Nat. Protoc. 2011, 6, 1612–1631. [Google Scholar] [CrossRef]
- Chung, Y.-M.; Simmons, K.L.; Gutowska, A.; Jeong, B. Sol−Gel Transition Temperature of PLGA- g -PEG Aqueous Solutions. Biomacromolecules 2002, 3, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Partlow, B.; Liu, J.; Zheng, Z.; Su, B.; Wang, Y.; Kaplan, D.L. Injectable silk-polyethylene glycol hydrogels. Acta Biomater. 2015, 12, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Goormaghtigh, E.; Cabiaux, V.; Ruysschaert, J.-M. Determination of Soluble and Membrane Protein Structure by Fourier Transform Infrared Spectroscopy. In Physicochemical Methods in the Study of Biomembranes; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1994; pp. 405–450. [Google Scholar]
- Belton, D.J.; Plowright, R.; Kaplan, D.L.; Perry, C.C. A robust spectroscopic method for the determination of protein conformational composition—Application to the annealing of silk. Acta Biomater. 2018, 73, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojdyr, M. Fityk: A general-purpose peak fitting program. J. Appl. Crystallogr. 2010, 43, 1126–1128. [Google Scholar] [CrossRef]
- Brown, J.; Lu, C.-L.; Coburn, J.; Kaplan, D.L. Impact of silk biomaterial structure on proteolysis. Acta Biomater. 2015, 11, 212–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez-Gómez, J.M.; Melchor-Moncada, J.J.; Veloza, L.A.; Sepúlveda-Arias, J.C. Purification and characterization of a metalloprotease produced by the C8 isolate of Serratia marcescens using silkworm pupae or casein as a protein source. Int. J. Biol. Macromol. 2019, 135, 97–105. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Y.; Li, L.; Guo, B.; Ma, P.X. Non-cytotoxic conductive carboxymethyl-chitosan/aniline pentamer hydrogels. React. Funct. Polym. 2014, 82, 81–88. [Google Scholar] [CrossRef]
- Gale, A.L.; Linardi, R.L.; McClung, G.; Mammone, R.M.; Ortved, K.F. Comparison of the Chondrogenic Differentiation Potential of Equine Synovial Membrane-Derived and Bone Marrow-Derived Mesenchymal Stem Cells. Front. Vet. Sci. 2019, 6, 178. [Google Scholar] [CrossRef]
- Uzieliene, I.; Bagdonas, E.; Hoshi, K.; Sakamoto, T.; Hikita, A.; Tachtamisevaite, Z.; Rakauskiene, G.; Kvederas, G.; Mobasheri, A.; Bernotiene, E. Different phenotypes and chondrogenic responses of human menstrual blood and bone marrow mesenchymal stem cells to activin A and TGF-β3. Stem Cell Res. Ther. 2021, 12, 251. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Zhang, Y.-Q.; Wei, Z.-G. Dissolution and processing of silk fibroin for materials science. Crit. Rev. Biotechnol. 2021, 41, 406–424. [Google Scholar] [CrossRef]
- Wu, X.; Gong, Y.; Xu, S.; Yan, Z.; Zhang, X.; Yang, S. Electrical Conductivity of Lithium Chloride, Lithium Bromide, and Lithium Iodide Electrolytes in Methanol, Water, and Their Binary Mixtures. J. Chem. Eng. Data 2019, 64, 4319–4329. [Google Scholar] [CrossRef]
- Fried, I.; Segal, M. Electrical conductivity of concentrated lithium bromide aqueous solutions. J. Chem. Eng. Data 1983, 28, 127–130. [Google Scholar] [CrossRef]
- Carissimi, G.; Lozano-Pérez, A.A.; Montalbán, M.G.; Aznar-Cervantes, S.D.; Cenis, J.L.; Víllora, G. Revealing the Influence of the Degumming Process in the Properties of Silk Fibroin Nanoparticles. Polymers 2019, 11, 2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasoju, N.; Hawkins, N.; Pop-Georgievski, O.; Kubies, D.; Vollrath, F. Silk fibroin gelation via non-solvent induced phase separation. Biomater. Sci. 2016, 4, 460–473. [Google Scholar] [CrossRef]
- Egan, G.; Phuagkhaopong, S.; Matthew, S.A.L.; Connolly, P.; Seib, F.P. Impact of silk hydrogel secondary structure on hydrogel formation, silk leaching and in vitro response. Sci. Rep. 2022, 12, 3729. [Google Scholar] [CrossRef]
- Cao, Z.; Chen, X.; Yao, J.; Huang, L.; Shao, Z. The preparation of regenerated silk fibroin microspheres. Soft Matter 2007, 3, 910. [Google Scholar] [CrossRef]
- Matsumoto, A.; Chen, J.; Collette, A.L.; Kim, U.-J.; Altman, G.H.; Cebe, P.; Kaplan, D.L. Mechanisms of Silk Fibroin Sol−Gel Transitions. J. Phys. Chem. B 2006, 110, 21630–21638. [Google Scholar] [CrossRef]
- Bai, S.; Zhang, X.; Lu, Q.; Sheng, W.; Liu, L.; Dong, B.; Kaplan, D.L.; Zhu, H. Reversible Hydrogel–Solution System of Silk with High Beta-Sheet Content. Biomacromolecules 2014, 15, 3044–3051. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Joseph, E.; Chaudhari, A.; Nisal, A.; Sharma, K.P. Viscoelastic Liquid from Regenerated Silk Fibroin in the Silk I Conformation: A Writeable and Shapeable Material. ACS Appl. Polym. Mater. 2022, 4, 4699–4708. [Google Scholar] [CrossRef]
- Suzuki, Y.; Asakura, T. Local conformation of serine residues in a silk model peptide, (Ala–Gly–Ser–Gly–Ala–Gly)5, studied with solid-state NMR:REDOR. Polym. J. 2010, 42, 354–356. [Google Scholar] [CrossRef]
- Kambe, Y.; Mizoguchi, Y.; Kuwahara, K.; Nakaoki, T.; Hirano, Y.; Yamaoka, T. Beta-sheet content significantly correlates with the biodegradation time of silk fibroin hydrogels showing a wide range of compressive modulus. Polym. Degrad. Stab. 2020, 179, 109240. [Google Scholar] [CrossRef]
- Su, D.; Yao, M.; Liu, J.; Zhong, Y.; Chen, X.; Shao, Z. Enhancing Mechanical Properties of Silk Fibroin Hydrogel through Restricting the Growth of β-Sheet Domains. ACS Appl. Mater. Interfaces 2017, 9, 17489–17498. [Google Scholar] [CrossRef] [PubMed]
- Samal, S.K.; Kaplan, D.L.; Chiellini, E. Ultrasound Sonication Effects on Silk Fibroin Protein. Macromol. Mater. Eng. 2013, 298, 1201–1208. [Google Scholar] [CrossRef]
- de Vos, W.M.; Voorhorst, W.G.; Dijkgraaf, M.; Kluskens, L.D.; Van der Oost, J.; Siezen, R.J. Purification, characterization, and molecular modeling of pyrolysin and other extracellular thermostable serine proteases from hyperthermophilic microorganisms. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2001; pp. 383–393. [Google Scholar]
- Sharma, C.; Jha, N.K.; Meeran, M.F.N.; Patil, C.R.; Goyal, S.N.; Ojha, S. Serratiopeptidase, A Serine Protease Anti-Inflammatory, Fibrinolytic, and Mucolytic Drug, Can Be a Useful Adjuvant for Management in COVID-19. Front. Pharmacol. 2021, 12, 603997. [Google Scholar] [CrossRef]
- Pawcenis, D.; Smoleń, M.; Aksamit-Koperska, M.A.; Łojewski, T.; Łojewska, J. Evaluating the impact of different exogenous factors on silk textiles deterioration with use of size exclusion chromatography. Appl. Phys. A 2016, 122, 576. [Google Scholar] [CrossRef] [Green Version]
- Elliott, W.H.; Bonani, W.; Maniglio, D.; Motta, A.; Tan, W.; Migliaresi, C. Silk Hydrogels of Tunable Structure and Viscoelastic Properties Using Different Chronological Orders of Genipin and Physical Cross-Linking. ACS Appl. Mater. Interfaces 2015, 7, 12099–12108. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.-Z.; Weng, Y.-J.; Zhang, Y.-Q. Enhancing the In Vitro Biological Activity of Degraded Silk Sericin and Its Analog Metabolites. Biomolecules 2022, 12, 161. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, B. Biodegradation of Silk Biomaterials. Int. J. Mol. Sci. 2009, 10, 1514–1524. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, X.; Li, G.; Wang, P.; Yang, Y. Tailoring degradation rates of silk fibroin scaffolds for tissue engineering. J. Biomed. Mater. Res. Part A 2019, 107, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Osama, I.; Gorenkova, N.; McKittrick, C.M.; Wongpinyochit, T.; Goudie, A.; Seib, F.P.; Carswell, H.V.O. In vitro studies on space-conforming self-assembling silk hydrogels as a mesenchymal stem cell-support matrix suitable for minimally invasive brain application. Sci. Rep. 2018, 8, 13655. [Google Scholar] [CrossRef]
- Simona, B.R.; Hirt, L.; Demkó, L.; Zambelli, T.; Vörös, J.; Ehrbar, M.; Milleret, V. Density gradients at hydrogel interfaces for enhanced cell penetration. Biomater. Sci. 2015, 3, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Ruijtenberg, S.; van den Heuvel, S. Coordinating cell proliferation and differentiation: Antagonism between cell cycle regulators and cell type-specific gene expression. Cell Cycle 2016, 15, 196–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Mulati, M.; Talib, S.Z.A.; Kaldis, P.; Takeda, S.; Okawa, A.; Inose, H. The Indispensable Role of Cyclin-Dependent Kinase 1 in Skeletal Development. Sci. Rep. 2016, 6, 20622. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; McCaffery, J.M.; Spencer, R.G.S.; Francomano, C.A. Hyaline cartilage engineered by chondrocytes in pellet culture: Histological, immunohistochemical and ultrastructural analysis in comparison with cartilage explants. J. Anat. 2004, 205, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Lian, C.; Wang, X.; Qiu, X.; Wu, Z.; Gao, B.; Liu, L.; Liang, G.; Zhou, H.; Yang, X.; Peng, Y.; et al. Collagen type II suppresses articular chondrocyte hypertrophy and osteoarthritis progression by promoting integrin β1−SMAD1 interaction. Bone Res. 2019, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberton, P.; Dugonitsch, H.; Hartmann, B.; Li, P.; Farkas, Z.; Saller, M.; Clausen-Schaumann, H.; Aszodi, A. Aggrecan Hypomorphism Compromises Articular Cartilage Biomechanical Properties and Is Associated with Increased Incidence of Spontaneous Osteoarthritis. Int. J. Mol. Sci. 2019, 20, 1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, A.E.; Ackerman-Yost, J.C.; Nixon, A.J. A Comparison of Three-Dimensional Culture Systems to Evaluate In Vitro Chondrogenesis of Equine Bone Marrow-Derived Mesenchymal Stem Cells. Tissue Eng. Part A 2013, 19, 2275–2283. [Google Scholar] [CrossRef] [Green Version]
- Bian, H.; Zhu, T.; Liang, Y.; Hei, R.; Zhang, X.; Li, X.; Chen, J.; Lu, Y.; Gu, J.; Qiao, L.; et al. Expression Profiling and Functional Analysis of Candidate Col10a1 Regulators Identified by the TRAP Program. Front. Genet. 2021, 12, 1037. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, S.; Naccarato, T.; Prakash-Damani, M.; Chou, Y.; Chu, C.-Q.; Zhu, Y. Regeneration of hyaline-like cartilage in situ with SOX9 stimulation of bone marrow-derived mesenchymal stem cells. PLoS ONE 2017, 12, e0180138. [Google Scholar] [CrossRef] [Green Version]
- Ng, K.W.; Kugler, L.E.; Doty, S.B.; Ateshian, G.A.; Hung, C.T. Scaffold degradation elevates the collagen content and dynamic compressive modulus in engineered articular cartilage. Osteoarthr. Cartil. 2009, 17, 220–227. [Google Scholar] [CrossRef]
- Armiento, A.R.; Alini, M.; Stoddart, M.J. Articular fibrocartilage—Why does hyaline cartilage fail to repair? Adv. Drug Deliv. Rev. 2019, 146, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Amelot, A.; Mazel, C. The Intervertebral Disc: Physiology and Pathology of a Brittle Joint. World Neurosurg. 2018, 120, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.B.; Silva-Correia, J.; Ribeiro, V.P.; da Silva Morais, A.; Oliveira, J.M.; Reis, R.L. Engineering patient-specific bioprinted constructs for treatment of degenerated intervertebral disc. Mater. Today Commun. 2019, 19, 506–512. [Google Scholar] [CrossRef]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [Green Version]
- Zapalska-Sozoniuk, M.; Chrobak, L.; Kowalczyk, K.; Kankofer, M. Is it useful to use several “omics” for obtaining valuable results? Mol. Biol. Rep. 2019, 46, 3597–3606. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, J.; Ritchie, G.R.S.; Roumeliotis, T.I.; Jayasuriya, R.L.; Clark, M.J.; Brooks, R.A.; Binch, A.L.A.; Shah, K.M.; Coyle, R.; Pardo, M.; et al. Integrative epigenomics, transcriptomics and proteomics of patient chondrocytes reveal genes and pathways involved in osteoarthritis. Sci. Rep. 2017, 7, 8935. [Google Scholar] [CrossRef] [Green Version]
- Toyoda, E.; Sato, M.; Takahashi, T.; Maehara, M.; Okada, E.; Wasai, S.; Iijima, H.; Nonaka, K.; Kawaguchi, Y.; Watanabe, M. Transcriptomic and Proteomic Analyses Reveal the Potential Mode of Action of Chondrocyte Sheets in Hyaline Cartilage Regeneration. Int. J. Mol. Sci. 2019, 21, 149. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Liang, J.; Guo, B.; Zhang, Y.; Hui, Q.; Chang, P.; Tao, K. Adipose-Derived Stem Cells Cocultured with Chondrocytes Promote the Proliferation of Chondrocytes. Stem Cells Int. 2017, 2017, 1709582. [Google Scholar] [CrossRef] [Green Version]
- Demoor, M.; Ollitrault, D.; Gomez-Leduc, T.; Bouyoucef, M.; Hervieu, M.; Fabre, H.; Lafont, J.; Denoix, J.-M.; Audigié, F.; Mallein-Gerin, F.; et al. Cartilage tissue engineering: Molecular control of chondrocyte differentiation for proper cartilage matrix reconstruction. Biochim. Biophys. Acta-Gen. Subj. 2014, 1840, 2414–2440. [Google Scholar] [CrossRef]
- Yodmuang, S.; McNamara, S.L.; Nover, A.B.; Mandal, B.B.; Agarwal, M.; Kelly, T.-A.N.; Chao, P.G.; Hung, C.; Kaplan, D.L.; Vunjak-Novakovic, G. Silk microfiber-reinforced silk hydrogel composites for functional cartilage tissue repair. Acta Biomater. 2015, 11, 27–36. [Google Scholar] [CrossRef]
- Yuan, T.; Li, Z.; Zhang, Y.; Shen, K.; Zhang, X.; Xie, R.; Liu, F.; Fan, W. Injectable Ultrasonication-Induced Silk Fibroin Hydrogel for Cartilage Repair and Regeneration. Tissue Eng. Part A 2021, 27, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.P.; da Silva Morais, A.; Maia, F.R.; Canadas, R.F.; Costa, J.B.; Oliveira, A.L.; Oliveira, J.M.; Reis, R.L. Combinatory approach for developing silk fibroin scaffolds for cartilage regeneration. Acta Biomater. 2018, 72, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, B.; Li, M.; Li, J.; Wan, Y. Enhanced dual network hydrogels consisting of thiolated chitosan and silk fibroin for cartilage tissue engineering. Carbohydr. Polym. 2020, 227, 115335. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuluaga-Vélez, A.; Toro-Acevedo, C.A.; Quintero-Martinez, A.; Melchor-Moncada, J.J.; Pedraza-Ordoñez, F.; Aguilar-Fernández, E.; Sepúlveda-Arias, J.C. Performance of Colombian Silk Fibroin Hydrogels for Hyaline Cartilage Tissue Engineering. J. Funct. Biomater. 2022, 13, 297. https://doi.org/10.3390/jfb13040297
Zuluaga-Vélez A, Toro-Acevedo CA, Quintero-Martinez A, Melchor-Moncada JJ, Pedraza-Ordoñez F, Aguilar-Fernández E, Sepúlveda-Arias JC. Performance of Colombian Silk Fibroin Hydrogels for Hyaline Cartilage Tissue Engineering. Journal of Functional Biomaterials. 2022; 13(4):297. https://doi.org/10.3390/jfb13040297
Chicago/Turabian StyleZuluaga-Vélez, Augusto, Carlos Andrés Toro-Acevedo, Adrián Quintero-Martinez, Jhon Jairo Melchor-Moncada, Francisco Pedraza-Ordoñez, Enrique Aguilar-Fernández, and Juan Carlos Sepúlveda-Arias. 2022. "Performance of Colombian Silk Fibroin Hydrogels for Hyaline Cartilage Tissue Engineering" Journal of Functional Biomaterials 13, no. 4: 297. https://doi.org/10.3390/jfb13040297