1. Introduction
There are decades of evidence that sablefish (
Anoplopoma fimbria) caught on the continental slope off of Alaska can survive capture from depths down to 1000 m. The National Marine Fisheries Service has been tagging and releasing sablefish in waters off of Alaska annually since 1979; from 1979 to 2009 over 300,000 tagged fish were released and 27,000 had been recovered [
1]. These fish are handled with care after they are brought onboard to minimize injuries that may occur from thrashing and being held out of water, but there is still mortality associated with capture and release [
2]. Sablefish that are tagged do not experience the stress of being discarded during regular hook-and-line and pot longline fishing operations, which are the major gear types used in the directed fishery [
3]. Fish exert effort while being brought up from deep depths and can also be injured during capture and discarding. Stressors from these processes may lead to higher rates of mortality, injury, and impaired health than in carefully tagged and released sablefish. However, the stress of capture and tagging in itself may also lead to mortality associated with physical trauma or changes in behavior.
Survival and health effects of capture have relevance to assumptions of tagging studies and fishery discard mortality rates. An overall sablefish discard mortality rate for longline gear of 11.7% was estimated from a mark and recapture study, when there was an assumed 96.5% survival rate for those with minor injuries [
2]. The authors suggested that this estimate of discard mortality may be artificially low because fish were handled carefully, such as being held in water and given recuperation time when onboard prior to tagging, unlike during regular fishing operations. Tagging studies require a large number of fish and many years or decades of recaptures and not all tagged fish are recovered. Results cannot be used to experimentally test the impacts of specific stressors and do not allow for monitoring of individual fish. Unlike tagging studies, controlled experiments have been used to identify predictors of mortality, such as reflexes and other indications of health [
4,
5].
The directed sablefish fishery is in deep water between ~300 and 700 m and currently requires that all sablefish caught be retained. When an individual’s fishing quota has been filled, all remaining catch must be discarded [
3]. On pot boats in particular this requires some on-deck time out of water to discard sablefish. Pot gear is becoming more prevalent in the fishery throughout Alaska, since 2017 when it became legal in the Gulf of Alaska; the proportion of the longline catch in pots in the Gulf of Alaska grew from 12% in 2018 to 44% in 2020 [
3]. The amount of time a fish spends onboard before discarding depends on the deck size, quantity of fish caught, and how many pots are emptied before discarding starts. Sablefish harvested with pot gear can spend a considerable amount of time out of the water while the catch is tended to and sorted, which could lead to prolonged exposure to air and heightened stress levels. With hook-and-line gear, it is likely that fish would be released quickly at the rail of the boat, limiting time out of water, but still experiencing the stress of capture.
Sablefish are fast growing, mature between the ages of 5 and 10, and are long-lived; the maximum observed age is 92 [
3,
6]. When there are sporadic, large year classes, small fish are proportionally more abundant and are difficult to avoid in fisheries. These smaller fish are generally <65 cm, <6 years old, immature, and have not yet contributed to the population (0% mature at age 2; 50% of fish are mature at ~age 7–8, depending on the study) [
7]. These year classes have been more common than ever before, with 2014, 2016, and 2017 being the highest since 1977 [
3]. Small fish have a lower value per pound, with a progressively higher value with increasing size. Due to these economic concerns, there is interest in removing the prohibition on discarding sablefish when there is still available quota. In response, the North Pacific Fishery Management Council (NPFMC) requested an analysis be prepared to evaluate implications of “careful release” of sablefish during fishing operations on stock assessment, fishery management, fishery economics, catch accounting, law enforcement, and fishery monitoring [
8]; the term “careful release” is used in the NPFMC analysis and the term “discard” is used in Alaska groundfish management and stock assessments. Presumably, if this management action occurs, the fleet would be allowed to carefully release fish following capture rather than being required to retain everything that is caught. The effect of any future regulation would be that fish may be sorted on-deck on pot boats and then discarded, even when quota is unfilled. Fish on hook-and-line longline boats would likely be discarded before being hauled on-board, but they would still experience the stress of capture.
Capture and handling fish puts stressors on various tissues and biological systems. When fish experience a stress, such as those incurred during capture and on-deck sorting, they have a physiological response that leads to either (1) recovery, (2) acute mortality, or (3) chronic stress. Any level of stress may push fish out of their biological equilibrium, making them susceptible to disease and parasites that can cause impaired growth or reproduction or delayed mortality [
5,
9]. A reflex action is an involuntary movement in response to a stimulus. Reflex impairment tests have been used extensively to develop models to predict acute and delayed mortality of fish after simulated trawling [
10,
11,
12], fishery trawling [
13,
14], simulated hook-and-line angling [
15], sorting of fish in aquaculture [
16], and other stressful processes. It is important to use multiple reflexes in conjunction because the combination of variables increases the precision of reflexes as a predictor of survival [
5,
17]. Observations of blood physiology, and sometimes injury, have been used in conjunction with reflex tests [
14,
18,
19]; however, we have not identified any systematic studies that evaluated the effects of stress on long-term health by evaluating both reflex testing and histological evaluation of tissues. In-depth health assessments using gross and histologic evaluation of tissues can identify inflammation, degeneration, necrosis, cellular proliferation, and other cellular changes, as well as infections with parasites and microorganisms [
20]. These conditions may be indicative of future survival or productivity, depending on the type and changes and their severity. They can aid in identification of the mechanism of acute or delayed mortality, which can be used to relate reflex results to health concerns.
The objectives of this study were to evaluate (1) if capture from deep-water affects the immediate reflex performance of sablefish and if reflex impairment is related to acute or delayed mortality or tissue health; (2) if time out of water, as part of a laboratory experiment, affects tissue health, delayed effects on reflexes, or acute or delayed mortality; and (3) if tissue health can be used to describe the mechanism of mortalities.
2. Materials and Methods
2.1. Sampling
A commercial longline vessel, F/V Seaview, was chartered to conduct fishing operations between: 57.37 N, −134.77 W and 57.73 N, −134.89 W in Lynn Canal, Southeast Alaska from 30 June–1 July 2019. Gear was set using 18/0 circle hooks spaced 2.7 m apart at depths from 437 to 658 m. Gear was soaked for 3 h to minimize the potential for copepod infestation (Orchomenella cf. pacifica). Just after fish were hauled onboard, they were chosen for the study if they were not bleeding from the gills, which is indicative of acute mortality, and if their total length was <59 cm.
2.2. Reflex Tests
Just after capture, fish were placed individually into a plastic container 90 cm long by 34 cm wide by 40 cm high filled with 60 L of seawater chilled for performing reflex tests 3.5–4.0 °C. The container was large enough so that movement was not restricted, they had ample distance between them and the water’s surface, and the temperature would remain stable over the 1 to 2 min. that it took to perform the tests. We conducted a series of tests to determine if a lack of a reflex was indicative of subsequent health or mortality. A total of six reflex tests were performed. First a gag reflex test was conducted by holding the head with one hand for stability and inserting a finger into the buccal cavity and touching the throat. A positive response was flaring of the operculum and opening of the mouth more widely. Second, an operculum flare reflex was tested by opening the mouth wide and releasing, with the expectation of a flare and then a return to an unflared, closed mouth. Third, the vestibular-ocular response was tested by holding the fish above the surface of the water with two hands by the middle with minimal pressure and rotating it 90 degrees to the right and the left around the long body axis. A positive reflex was when the eyes tracked the tester. Fourth, a reflex to sound vibration was tested by tapping on the side of the holding tank and watching for a startled movement, sideways or forward [
11]. Fifth, when the fish was sitting calmly, the tail was grabbed and released and a positive response was when the fish swam forward. Sixth, the fish was turned upside-down and a positive reflex was recorded if it quickly righted itself (tests reviewed by Davis [
5]).
2.3. Holding and Experiment
After reflex tests, sablefish were held in aerated, chilled water in insulated totes with lids for 1–2 days on the vessel. Water was maintained at 4.0–5.0 °C. The fish were transported to the National Oceanic and Atmospheric Administration’s, Auke Bay Laboratories in Juneau, Alaska, where they were maintained in 24 h darkness in seawater chilled to 3.5–4.0 °C; during National Marine Fisheries Service (NMFS) bottom hook-and-line longline surveys on the continental slope of the Gulf of Alaska temperatures typically range from ~3.75 to 4.5 °C from 300 to 800 m (K. Siwicke, NMFS, pers. comm.). Red light was used when feeding or conducting experiments because deepwater fish are only sensitive to blue light at wavelengths from 450–500 nm, as those encompass the bioluminescent and residual daylight available below 200 m [
21].
Fish were randomly divided into groups of eight and maintained in three Frigid Units Inc. insulated Living Stream System® tanks with their false bottoms removed and Styrofoam used as a lid to maintain the temperature. These rectangular, 530 L tanks have inflow and outflow ends; water was not recirculated and the flow rate was 34 L/h. Intake water was pumped from 25 m deep in a deep-water channel, was chilled in a separate 2270 L tank, and was fed to the rectangular tanks through insulated pipes. After the first week, fish were fed pieces of salmon and squid twice a week until satiation over the course of 30 min.
Fish were held in the laboratory for 35–45 days prior to experiments to allow time for delayed health issues to reveal themselves and to allow fish to recover from acute effects of capture that could affect their behavior or health. Eight fish were assigned to each treatment, either 3, 6, or 11 min out of water or a control with no time out of water. Fish were used in only one treatment. These times were chosen because they capture hypothetical times fish may be on-deck when being sorted and discarded during pot fishing with, hypothetical, allowable ”careful release” or during required discarding after quota has been filled. Three additional fish were collected at-sea for tissue histology as a way to see what tissue damage or natural irregularities may have been present after the acute stress of capture.
For the experiment the fish was held out of water by placing it in a dry fish tote with a base of 90 by 90 cm. The fish was placed back into a tank with water after the specified 3, 6, or 11 min. had elapsed. The fish sometimes moved around in the tote, but the bottom was smooth and did not cause abrasion. After a holding period of 7–10 days, the same reflexes were tested, fish were euthanized using a blow to the head, length and weight were measured, and tissues were examined and collected for a microscopic evaluation, as described below. The holding period allowed for observations of acute or delayed mortality. Reflex tests were not performed just after the experiments to avoid adding any potential effects of the tests, which would have confounded the results of the time out of water experiment. The difference in length was examined for growth and the difference in weight was examined for increases and decreases, which may be indicative of the stress of capture or holding. In total, fish were in the laboratory for 45–52 days.
2.4. Necropsy and Histology
After the experimental treatment and euthanasia, the eyes were examined to highlight any abrasions, lesions, or parasites on the cornea by applying a solution of fluorescein (100 mg/L) and sodium bicarbonate (800 mg/L). Fluorescein is commonly used on human eyes and fish skin to detect corneal damage or lesions. The solution pools into or adheres to irregular and damaged surfaces and is visible in darkness when illuminated with ultraviolet light (480–600 nm) ([
22]). Eyes were also examined for any cloudiness or other aberrations.
Tissues were collected for histological evaluations, including the gills, skin, eyes, brain, heart, liver, spleen, intestines, stomach, gonads, kidney, muscle, and, in a few cases, the vent and pharynx. Tissues were immediately placed in Excell Plus Fixative™ and stored at room temperature. After completion of the experimental portion of the study, samples were shipped to the Oregon Veterinary Diagnostic Laboratory (OVDL) at Oregon State University for further processing.
Tissues were processed to paraffin-embedded blocks following standard operating procedures at the OVDL. In brief, after trimming tissues to fit cassettes, tissues were processed overnight to replace water with increasing concentrations of ethanol, xylene, and ultimately paraffin. Tissues were embedded in paraffin. From each block, a 3–5 μm section was cut, deparaffinized, stained with haematoxylin and eosin (H&E), dehydrated, and cover slipped. Slides were examined using bright-field microscopy by a board certified veterinary anatomic pathologist (CL) using a Nikon Eclipse E400 microscope. Each tissue was examined histologically for presence of any findings outside the normal limits including but not limited to inflammation (cellular infiltrates, exudate, granulations tissue, fibrosis), degeneration, necrosis, cellular proliferation, and presence of internal or external infections organisms such as parasites and microorganisms (including bacteria and fungi) [
23,
24]. Descriptions of any tissues changes included the type of change and a combined score for extent and severity: none, minimal, mild, moderate, or severe.
4. Discussion
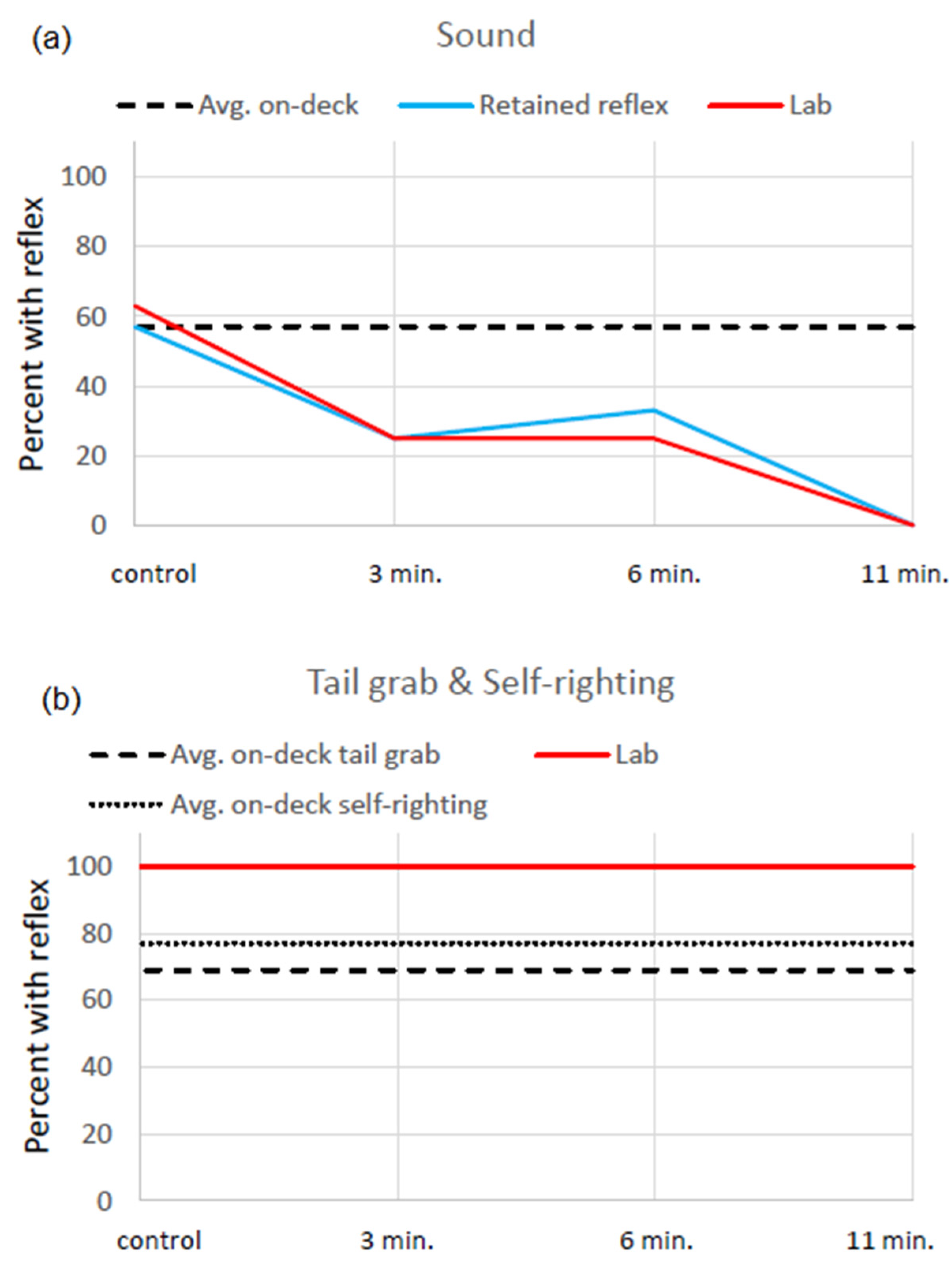
Sablefish in our study all survived capture and being held out of water for up to 11 min. in a laboratory environment. The tissue irregularities were minor, no clear associations could be established with capture or time out of water, and they were likely present prior to capture. Importantly, none of these histologic findings appeared to affect long-term health while in the laboratory and are highly unlikely to affect overall health in the future. The self-righting and tail grab reflexes were absent in some fish after capture but were present in the laboratory. The sound reflex was impaired after capture and was worse in the laboratory after the time out of water experiment, even after a period of 7–10 days of recuperation. The lack of reflexes may lead to acute or delayed mortality of tagged and released or fishery discarded fish.
The sound reflex was commonly in fish on-deck and those with time out of water treatments. On-deck there was a positive reflex in 57% (20/35) fish, demonstrating that there are acute effects of capture. For the laboratory control group, the sound reflex was apparent on-deck in 7 fish and retained in 4 fish at the end of the study (57%, 4/7). The loss of the reflex may be a delayed effect of capture or could be due to transport or holding in the laboratory. The laboratory treatment groups also had fish that lost their sound reflex; the proportion of fish that retained the reflex was moderate for 3 and 6 min. fish and nil for the 11 min. treatment. The continual decrease in the sound reflex with treatment indicates that, although there may be other contributing factors, time out of water contributed to the lack of a sound reflex in some fish, even after 7–10 days of recuperation. Similar to our study, simulated trawling of 1- and 2-year-old sablefish (36–56 cm), which included many stressors, had lasting effects on the sound and a startle reflex for 3–24 h [
11]. The combination of our results and those of the trawled sablefish show that there is consistency to the reflex impairment types of young sablefish (45–59 cm in our study). These impairments that are related to capture will likely have an effect on behavior when discarded.
The sound response may be important for locating prey or for evading predators, as sablefish are one of the few deep-sea fish that have been shown to make sound and, therefore, may hear sound [
25]. Sperm whales (
Physeter microcephalus) follow hook-and- line longline fishing vessels in the Gulf of Alaska and depredate on the gear while hauling [
26,
27]. Sperm whales make sounds for echolocation when depredating on longline gear and when feeding on sablefish [
28,
29]. Sablefish acoustics range from 344 Hz to 34 kHz [
24], which are very similar to sperm whales (100 Hz–30 kHz) [
30,
31]. Although sablefish have the ability to make sound, it is unknown if they detect sound or what frequencies they can detect, and what the hearing mechanism may be because they lack a swim bladder [
25]. Despite these unknowns, it is possible that sablefish can hear some sperm whale sounds. If a response to acoustics helps sablefish evade predators after being discarded or released, they would be at a risk of predation for at least a week afterward, as we sampled fish 7–10 days after applying the stress of being out of water. In addition, orcas (
Orcinus orca) depredate on longline gear and are known to make sounds during feeding and traveling [
32]. They are also nimble and can quickly home in on individual fish [
26]. The decreased sound (57% success), tail grab (69% success), and self-righting reflexes (77% success) on-deck, which can be attributed to capture alone, may together affect sablefish swim response and increase their susceptibility to predators present standing by during hauling, tag and release, and discarding fish. This may occur even after careful treatment of sablefish when tagging and in hook-and-line fisheries when fish are removed from hook at the rail of the vessel before being brought on-board. If there is a regulation in the future to allow discarding prior to quota being filled, whales may also start to follow boats that fish pot gear.
Although sablefish do lack a swim bladder, which is known to be used for detecting sound, the sound reflex test may be a measure of the fish’s responsiveness to sound in multiple ways. Fish are known to use otoliths to detect sound pressure and particle motion as well as the lateral line [
33]. Otoliths can be used to detect sound directly via the otolith’s response to motion and the lateral line is known to be responsive to water displacement [
34]. These pathways would likely be sensitive to the sound vibration that was applied in our study because it was in close proximity to the fish. Although it may not be possible to isolate the cause of the lack of reflex in sablefish or know the importance of any physical adaptations to detect sound [
35], it is important to note that there are multiple ways that fish may detect sound. There are very few studies of deepwater fish sound detection and sound creation because of the logistics of working with deep-water species and sound in situ and in the laboratory.
There were several irregularities noted in tissues that were not caused by the experiments, including prevalent issues in the gills, such as mild branchitis and structural changes to gill lamellae, including epithelial hyperplasia. If these issues were caused by trauma, we would expect to see hemorrhage, but this was absent. The issues in the gills are commonly seen in many fish in response to irritation [
24] and interpreted as non-specific. Epithelial hyperplasia in the gills in particular is a structural change that can affect the function of the gills, most notably the transfer of oxygen from water to blood, if severe enough. These changes were mild in fish euthanized immediately after capture as well as fish kept in tanks. We did not identify any negative effects of these gill issues; there was no evidence of respiratory distress and no mortalities. Furthermore, adult sablefish have shown high tolerance to increased temperatures and lower oxygen levels [
36]. Therefore, we interpreted the observed gill conditions as background lesions and not related to the experiment.
None of the granulomas and/or parasites observed in sablefish here, by themselves, would be considered potentially fatal, particularly at the observed low severity levels. In wild fish low level infections with parasites are the norm, most commonly involving metazoans including nematodes, and often leading to development of granulomas (the stereotypical inflammatory response of fish to many insults). Within areas of inflammation, parasites are not always present, as they migrate through tissues and may leave inflammation in their wake [
23,
24]. In the case of the stomach, mesentry, and kidney, parasites were present but there was no associated inflammation. The lack of associated inflammation suggests that fish did not mount an immune response to the parasites, a common phenomenon following co-evolution of parasite and host [
23,
24]. Conversely, the mild inflammation we observed in the brain, stomach (submucosa), and intestine (a) may have been caused by parasite infection(s) or other mild insults [
23,
24]. Both the inflammation and the parasites we observed were likely present in fish tissues in the wild prior to the experiment, and are unlikely to have caused any health issues. We did not measure how stress may have suppressed the immune system, as it has in sablefish after simulated trawling [
9]. This stress response requires energy to be reallocated to restoring homeostasis and may strain the fish and lead to delayed disease. We did not observe mortalities or disease over the 49– 52 day holding period and we did not treat the influent for pathogens, demonstrating that effects of any immunosuppression in the laboratory were not significant; however, the stressors and pathogens may differ in the wild.
It is important to consider what sizes of fish will be affected by fishing activity. We focused on fish in the smaller size range (ages 2–5) that are encountered by commercial longline gear [
3]. These fish are the most likely to be “carefully released” if it becomes an option before quota is filled, due to their low price per pound [
8]. Small fish are mostly immature and their ability to grow and spawn will determine the trajectory of the Alaska stock for many years to come. The fish in our study survived capture and time out of water with the most notable deficiency being reflexes. The effect of stressors may vary based on fish size. As an example, in a trawl simulation study there was a correlation between delayed mortality and behavior for age-2 sablefish and not age-1, even though their mortality rate was higher and more immediate [
11]. It is important to evaluate the effects of fishing stress on the range of sizes that may be encountered and discarded, which may extend to larger sizes than we studied. Sample size should also be a consideration. In particular we only performed histology on three fish on-deck. As these are an indication of natural health conditions and acute effects of capture, we recommend increasing the sample size of the fish sampled after capture in the future.
We had no mortalities throughout the study and all fish ate and swam normally in the laboratory while being observed; however, we only investigated some of the stressors fish experience during capture and on-deck sorting. Furthermore, our chosen times of 3, 6, and 11 min. were our best estimate of the time it may take to discard fish based on conversations with fishers and should accurately reflect on-deck times that fish are exposed to during commercial fishing operations. We handled fish carefully and paid close attention not to drop them or cause abrasion, which has been linked to mortality in trawled sablefish [
11]. The experiences of each fish will depend on many factors, including how quickly they are sorted, the gear specifications, the size of the boat, sorting area, weather conditions, efficiency of the crew during hauling, and how the fish are handled before being returned to the water. During these processes there is ample potential for external and internal injuries and causes for reflex impairment that we did not focus on.
In the future, we recommend that more potential stressors that occur during longline and pot gear capture and discarding be tested at-sea and experimentally in the laboratory. In the current study, we were able to look at the effects of the stress of capture on hook-and-line longline gear through being unhooked. We also isolated a single stressor that may occur during the discarding process on pot longline vessels: time out of water. It will be important to observe active fishing and to identify a suite of stressors that fish are exposed to during pot and hook-and-line longline fishing, including being dropped and being underneath the weight of other fish in pots, which may occur when fish are on top of one another prior to sorting. Each of these stressors should be tested individually and in combination to identify which factors are the most impactful to behavior, health, and mortality. In situ studies of discard survival would also be an important addition, and could be used for a specific fishing gear and sorting practice.
Blood physiology has been used to predict mortality after exposure to stressors, in conjunction with reflex tests [
14,
18,
19]. The fish’s physiological stress response can be monitored before the stress, just after, and again after recuperation time to determine if there is a response and how long it lasts. We were not able to add this component to our study because the addition of a blood draw just before and after the treatment of being out of water would add additional time out of water and add the stress of being handled for blood samples. If study designs in the future allow for blood sampling, the results may provide more information on the mechanisms behind subsequent declining health or mortality.
Conventional tagging studies in the marine environment may take decades, but rapid results are possible with pop-up satellite archival tags. They can be used to determine whether a fish died after discarding by measuring acceleration or movement by depth. However, these tags may affect the mortality and behavior of the fish and can only be used on fish of a certain size, because of the tag’s size [
37]. There are unanswered questions associated with the sound reflex. Doing further tests on sablefish reaction to sound frequencies, associated with orcas and sperm whales, may reveal the implications of having a negative sound reflex after being released after tagging or discarded in fisheries.
Although we did not find any tissue irregularities that could be unequivocally linked to capture or the time out of water experiments, it does not diminish the importance of in-depth evaluations of fish health, including histological evaluations of tissues. Pairing histological observations with reflex testing and observations of mortality can provide a full picture of the mechanisms behind mortality, such as ruptures, inflammation, degeneration, necrosis, cellular proliferation, and disease [
20]. These evaluations are recommended for sablefish and other species, particularly when working with new stressors or a new species. It is necessary to work with a histopathologist who has worked with fish and can recognize which issues are commonplace, which were caused by stress, their severity, and what the related risks are to health. In any species, a comprehensive evaluation of the naturally occurring irregularities are advised, to serve as a baseline for comparison to fish tissues from experiments.
In summary, sablefish survived capture in the short- and long-term in the laboratory, in a controlled environment, and being held out of water for up to 11 min. Some fish lacked reflexes, which may make them susceptible to whale predation after being discarded and other feeding and predation concerns at-depth. The sound reflex may worsen with time since capture and with time out of water, which could impact sablefish feeding, communication, and survival in the short- and long-term. Although we did not find any microscopic issues that were caused by the experiment, a histological evaluation is still an important part of any study that evaluates the utility of indicators of mortality.







{kind=link}
{kind=link}
{kind=link}