Phytotoxicity in Seedlings of Rhizophora mangle (L.) Exposed to 2,4-Dichlorophenoxyacetic Acid under Experimental Conditions

 , ,
, ,
Abstract
:1. Introduction
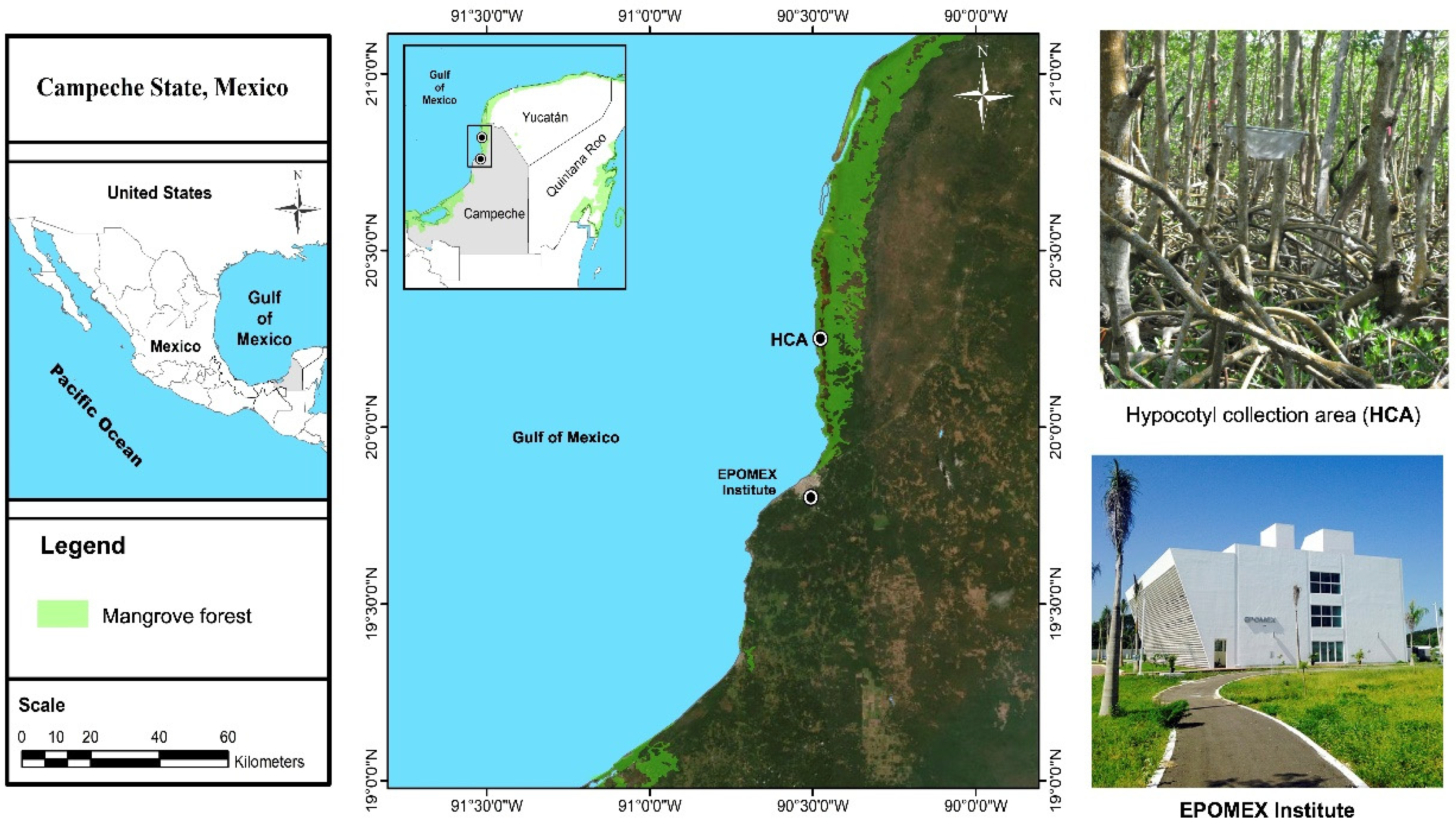
Description of the Rhizophora mangle Hypocotyl Collection Area
2. Materials and Methods
2.1. Construction and Environmental Stabilization of the System
2.2. Stages of Reforestation and Conditioning
2.3. Application of 2,4-Dichlorophenoxyacetic (2,4-D)
2.4. Effects on Chlorophyll Growth and Production
2.4.1. Effects on Growth in the Seedlings of Rhizophora mangle
2.4.2. Chlorophyll Concentration
2.5. Statistical Analysis
3. Results
3.1. Physicochemical Parameters before and during the Application of the Herbicide
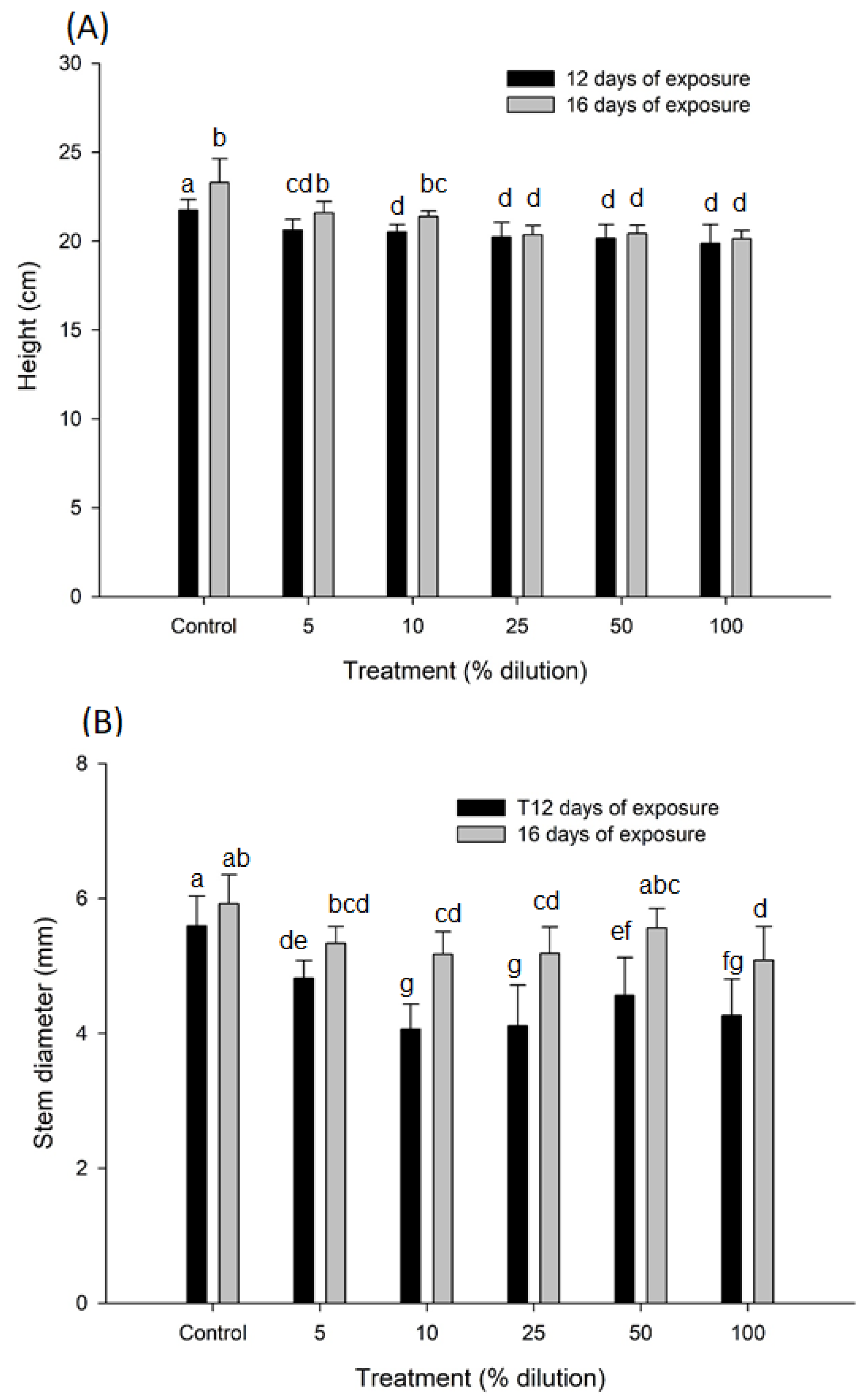
3.2. Responses to Growth
3.3. Behavior of the Concentration of Chlorophyll a
3.4. Biomass Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turner, B.L.; Kasperson, R.E.; Matson, P.A.; McCarthy, J.J.; Corell, R.W.; Christensen, L.; Eckley, N.; Kasperson, J.X.; Luers, A.; Martello, M.L.; et al. A framework for vulnerability analysis in sustainability science. Proc. Natl. Acad. Sci. USA 2003, 100, 8074–8079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazda, Y.; Magi, M.; Kogo, M.; Hong, P.N. Mangroves as a coastal protection from waves in the Tong King delta, Vietnam. Mangroves Salt Marshes 1997, 1, 127–135. [Google Scholar] [CrossRef]
- Barbier, E.B. Ecosystems as Natural Assets. Found. Trends Microecon. 2008, 4, 611–681. [Google Scholar] [CrossRef]
- Alongi, D.; Sasekumar, A.; Chong, V.; Pfitzner, J.; Trott, L.; Tirendi, F.; Dixon, P.; Brunskill, G. Sediment accumulation and organic material flux in a managed mangrove ecosystem: Estimates of land–ocean–atmosphere exchange in peninsular Malaysia. Mar. Geol. 2004, 208, 383–402. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.; Chan-Keb, C.; Muñiz-Salazar, R.; Pérez-Balan, R.; Osti-Sáenz, J.; Gutiérrez-Alcántara, E.; Reyes-Castellano, J.; May-Colli, L.; Conde-Medina, K.; Ruiz-Hernández, J. Relationship between blue carbon and methane and the hydrochemistry of mangroves in southeast Mexico. Appl. Ecol. Environ. Res. 2020, 18, 1091–1106. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Alongi, D.M. Present state and future of the world’s mangrove forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.J.; Lam, M.H.; Lam, P.K.; Richardson, B.J.; Man, B.K.; Li, A.M. Concentrations of Persistent Organic Pollutants in Surface Sediments of the Mudflat and Mangroves at Mai Po Marshes Nature Reserve, Hong Kong. Mar. Pollut. Bull. 2000, 40, 1210–1214. [Google Scholar] [CrossRef]
- Peters, E.C.; Gassman, N.J.; Firman, J.C.; Richmond, R.H.; Power, E.A. Ecotoxicology of tropical marine ecosystems. Environ. Toxicol. Chem. 1997, 16, 12–40. [Google Scholar] [CrossRef]
- Duke, N.C.; Bell, A.M.; Pederson, D.K.; Roelfsema, C.; Nash, S.B. Herbicides implicated as the cause of severe mangrove dieback in the Mackay region, NE Australia: Consequences for marine plant habitats of the GBR World Heritage Area. Mar. Pollut. Bull. 2005, 51, 308–324. [Google Scholar] [CrossRef]
- Lee, S.Y. Tropical mangrove ecology: Physical and biotic factors influencing ecosystem structure and function. Austral Ecol. 1999, 24, 355–366. [Google Scholar] [CrossRef]
- Nath, B.; Birch, G.; Chaudhuri, P. Trace metal biogeochemistry in mangrove ecosystems: A comparative assessment of acidified (by acid sulfate soils) and non-acidified sites. Sci. Total. Environ. 2013, 463–464, 667–674. [Google Scholar] [CrossRef]
- Rai, P.K. Heavy Metal Pollution in Aquatic Ecosystems and its Phytoremediation using Wetland Plants: An ecosustainable approach. Int. J. Phytoremediation 2008, 10, 133–160. [Google Scholar] [CrossRef]
- Sánchez Páez, H.; Ulloa Delgado, G.A.; Alvarez León, R.; Gil Torres, W.O. Colombia. In Ministerio del Medio Ambiente; Asociación Colombiana de Reforestadores.; Organización Internacional de las Maderas Tropica-les. Hacia la recuperación de los Manglares del Caribe Colombiano; de Bogotá, S., Ed.; Ministerio del Medio Ambiente: Bogotá, Colombia, 2000; ISBN 9583316849. [Google Scholar]
- Cuong, D.T.; Bayen, S.; Wurl, O.; Subramanian, K.; Wong, K.K.S.; Sivasothi, N.; Obbard, J.P. Heavy metal contamination in mangrove habitats of Singapore. Mar. Pollut. Bull. 2005, 50, 1732–1738. [Google Scholar] [CrossRef]
- Dietz, K.J.; Baier, M.; Krämer, U. Free radicals and reactive oxygen species as mediators of heavy metal toxicity in plants. In Heavy Metal Stress in Plants; Springer: Berlin/Heidelberg, Germany, 1999; pp. 73–97. [Google Scholar]
- Ivens, G.W. Arboricides for killing mangroves. In West African Rice Research Station Periodical Science Report, 8; West Africa Rice Development association, WARDA-ADRAO: ABIDJAN 01, Cote d’ Ivoire, 1957. [Google Scholar]
- Teas, H.J. Herbicide Toxicity in Mangroves; EPA-600/3-76-004; National Technical Information Service: Springfield City, FL, USA, 1976. [Google Scholar]
- Peng, L.; Wenjian, Z.; Zhenji, L. Distribution and accumulation of heavy metals in Avicennia marina community in Shenzhen. China. J. Environ. Sci. 1997, 9, 472–479. [Google Scholar]
- MacFarlane, G. Leaf biochemical parameters in Avicennia marina (Forsk.) Vierh as potential biomarkers of heavy metal stress in estuarine ecosystems. Mar. Pollut. Bull. 2002, 44, 244–256. [Google Scholar] [CrossRef]
- Rivera-Arriaga, E.; Williams-Beck, L.; Hernández, L.E.V.; Arjona, M.E.G. Crafting grassroots’ socio-environmental governance for a coastal biosphere rural community in Campeche, Mexico. Ocean Coast. Manag. 2021, 204, 105518. [Google Scholar] [CrossRef]
- CONANP (Comisión Nacional de Áreas Naturales Protegidas). Programa de Conservación y Manejo RESERVA de la BIOSFERA los Petenes; CONANP: Mexico City, México, 2006. [Google Scholar]
- Armitage, D.; Alexander, S.; Andrachuk, M.; Berdej, S.; Brown, S.; Nayak, P.; Pittman, J.; Rathwell, K. Communities, multi-level networks and governance transformations in the coastal commons. Gov. Coast. Commons 2017, 231–251. [Google Scholar] [CrossRef]
- Bautista, F.; Palacio, A.G. Caracterización y Manejo de los Suelos de la Península de Yucatán: Implicaciones Agropecuarias, Forestales y Ambientales, 1st ed.; Instituto Nacional de Ecología: Mexico City, Mexico, 2005. [Google Scholar]
- Bautista, F.; Palacio, A.; Mendoza, J.; Kú, V.; Pool, L.; Cantarell, W. Suelos. In La Biodiversidad en Campeche: Estudio de Estado; Villalobos-Zapata, G.J., Mendoza-Vega, J., Eds.; Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad (CONABIO): Tlalpan, Mexico, 2010; p. 748. ISBN 978-607-7887-22-5. [Google Scholar]
- Troche-Souza, C.; Rodríguez-Zúñiga, M.T.; Velázquez-Salazar, S.; Valderrama-Landeros, L.; Villeda-Chavéz, E.; Alcántara-Maya, A.; Vázquez-Balderas, B.; Cruz-López, M.I.; Ressl, R. Manglares de México: Extensión, Distribución y Monitoreo (1970/1980–2015), 1st ed.; Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad.: Ciudad de México, Mexico, 2017; ISBN 978-607-8328-78-9. [Google Scholar]
- Velázquez-Salazar, S.; Rodríguez-Zúñiga, M.; Alcántara-Maya, J.; Villeda-Chávez, E.; Valderra-ma-Landeros, L.; Troche-Souza, C.; Vázquez-Balderas, B.; Pérez-Espinosa, I.; Cruz-López, M.I.; Ressl, R.; et al. Manglares de México. Actualización y Análisis de los Datos 2020, 1st ed.; Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad: Tlalpan, México, 2021; ISBN 9786078570508. [Google Scholar]
- Naidoo, G. The mangroves of South Africa: An ecophysiological review. S. Afr. J. Bot. 2016, 107, 101–113. [Google Scholar] [CrossRef]
- Chan-Keb, C.A.; Agraz-Hernández, C.M.; Perez-Balan, R.A.; Gómez-Solano, M.I.; Maldonado-Montiel, T.D.; Ake-Canche, B.; Gutiérrez-Alcántara, E.J. Acute toxicity of water and aqueous extract of soils from Champotón river in Lactuca sativa L. Toxicol. Rep. 2018, 5, 593–597. [Google Scholar] [CrossRef]
- Yanes, C.V.; Irene, A.; Muñoz, B.; Isabel, M.; Silva, A.; Gual Díaz, M.; Cristina, Y.; Dirzo, S. Árboles Mexicanos Potencialmente Valiosos Para la Restauración Ecológica y la Reforestación; Bases de datos SNIB-CONABIO, proyecto J084; Comisión Nacional Para el Conocimiento y uso de la Biodiversidad: Tlalpan, Mexico, 1999. [Google Scholar]
- Gill, A.M.; Tomlinson, P.B. Studies on the Growth of Red Mangrove (Rhizophora mangle L.) 4. The Adult Root System. Biotropica 1977, 9, 145. [Google Scholar] [CrossRef]
- Maricusa Agraz Hernández, C.; Armando, C.; Keb, C.; Iriarte-Vivar, S.; Venegas, G.P.; Serratos, B.V.; Osti Sáenz, J.; Hernández, A.; Keb, C.A.C.; Iriarte-Vivar, S.I.; et al. Phenological variation of Rhizophora mangle and ground water chemistry associated to changes of the precipitation. Hidrobiológica 2015, 25, 49–61. [Google Scholar]
- Cissell, J.R.; Delgado, A.M.; Sweetman, B.M.; Steinberg, M.K. Monitoring mangrove forest dynamics in Campeche, Mexico, using Landsat satellite data. Remote. Sens. Appl. Soc. Environ. 2018, 9, 60–68. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.M.; Del Río-Rodríguez, R.E.; Chan-Keb, C.A.; Osti-Sáenz, J.; Muñiz-Salazar, R. Nutrient Removal Efficiency of Rhizophora mangle (L.) Seedlings Exposed to Experimental Dumping of Municipal Waters. Diversity 2018, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Skoog, D.A.; West, D.M.; Holler, F.J.; Crouch, S.R. Fundamentals of Analytical Chemistry; Cengage Learning Group, Inc.: Independence, KY, USA, 2013; ISBN 9780495558286. [Google Scholar]
- Gervais, J.; Luukinen, B.; Buhl, K.; Stone, D. 2,4-D Technical Fact, National Pesticide Information Center, Oregon State University Extension Services. Available online: http://npic.orst.edu/factsheets/archive/2,4-DTech.html (accessed on 18 November 2021).
- Duke, N.C.; Zuleika, S.; Pinzon, M. Aging Rhizophora Seedlings from Leaf Scar Nodes: A Technique for Studying Recruitment and Growth in Mangrove Forests. Biotropica 1992, 24, 173. [Google Scholar] [CrossRef]
- Hernández, C.M.A.; Zaragoza, C.G.; Iriarte-Vivar, S.; Flores-Verdugo, F.J.; Casasola, P.M. Forest structure, productivity and species phenology of mangroves in the La Mancha lagoon in the Atlantic coast of Mexico. Wetl. Ecol. Manag. 2011, 19, 273–293. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Manual of Seawater Analysis; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1965; Volume 125. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2010; ISBN 9780131008465. [Google Scholar]
- NPIC 2,4-D Technical Fact Sheet. Available online: http://npic.orst.edu/factsheets/24Dgen.html (accessed on 16 November 2021).
- Mitsch, W.J.; Gosselink, J. Wetlands, 3rd ed.; John Wiley & Sons: New York, NY, USA, 2000. [Google Scholar]
- Gleason, S.; Ewel, K.; Hue, N. Soil redox conditions and plant–soil relationships in a micronesian mangrove forest. Estuar. Coast. Shelf Sci. 2003, 56, 1065–1074. [Google Scholar] [CrossRef]
- Ashton, F.M.; Crafts, A.S. Mode of Action of Herbicides, 1st ed.; Wiley-Interscience: New York, NY, USA, 1973. [Google Scholar]
- Culic, P. The effects of 2,4-D on the growth of Rhizophora stylosa Griff. seedlings. In Physiology and Management of Mangroves, Tasks for Vegetation Science; Teas, H.J., Ed.; Springer: Dordrecht, The Netherlands, 1984. [Google Scholar]
- Filella, I.; Penuelas, J. The red edge position and shape as indicators of plant chlorophyll content, biomass and hydric status. Int. J. Remote. Sens. 1994, 15, 1459–1470. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Tremblay, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote. Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Wu, C.; Niu, Z.; Tang, Q.; Huang, W. Estimating chlorophyll content from hyperspectral vegetation indices: Modeling and validation. Agric. For. Meteorol. 2008, 148, 1230–1241. [Google Scholar] [CrossRef]
- Hallam, N.D. The Effect of 2,4-Dichlorophenoxyacetic Acid and Related Compounds on the Fine Structure of the Primary Leaves of Phaseolus vulgaris. J. Exp. Bot. 1970, 21, 1031–1038. [Google Scholar] [CrossRef]
- White, J.A.; Hemphill, D.D. An Ultrastructural Study of the Effects of 2,4-D on Tobacco Leaves. Weed Sci. 1972, 20, 478–481. [Google Scholar] [CrossRef]
- Flores-De-Santiago, F.; Kovacs, J.M.; Flores-Verdugo, F. The influence of seasonality in estimating mangrove leaf chlorophyll-a content from hyperspectral data. Wetl. Ecol. Manag. 2013, 21, 193–207. [Google Scholar] [CrossRef]
- Chambers, A.J.; Victoria, A. Field Crop Herbicide Guide 1997–1998: Detailed Information on Australian Field Crop Herbicides 1997–98., 5th ed.; Kondinin Group: Perth, WA, Australia, 1997. [Google Scholar]
- Bell, A.M.; Duke, N.C. Effects of Photosystem II inhibiting herbicides on mangroves—Preliminary toxicology trials. Mar. Pollut. Bull. 2005, 51, 297–307. [Google Scholar] [CrossRef]
- Moorthy, P.; Kathiresan, K. Influence of ultraviolet-β radiation on photosynthetic and biochemical characteristics of a mangrove Rhizophora apiculata. Photosynthetica 1997, 34, 465–471. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | gl (Factor, Error) | Statistic F | p Value |
|---|---|---|---|
| Temperature (°C) | (11, 84) | 0.09 | 0.990 |
| Potential redox (mV) | (11, 84) | 0.12 | 0.990 |
| Salinity (g/kg) | (11, 84) | 0.68 | 0.757 |
| pH | (11, 84) | 1.06 | 0.404 |
| Parameters | gl (Factor, Error) | Statistic F | p Value |
|---|---|---|---|
| Temperature (°C) | (5, 18) | 0.22 | 0.940 |
| Potential redox (mV) | (5, 18) | 4.99 | 0.005 |
| Salinity (g/kg) | (5, 18) | 0.77 | 0.580 |
| pH | (5, 18) | 1.78 | 0.170 |
| Parameter | Factor | Gl | F | p |
|---|---|---|---|---|
| Height (cm) | Treatment | 5 | 26.24 | <0.001 |
| Time | 1 | 20.36 | <0.001 | |
| Treatment xTime | 5 | 2.46 | <0.030 | |
| Error | 90 | |||
| Stem Diameter (mm) | Treatment | 5 | 17.07 | <0.001 |
| Time | 1 | 74.12 | <0.001 | |
| Treatment xTime | 5 | 2.36 | <0.046 | |
| Error | 90 | |||
| Chlorophyll a (µg/g) | Treatment | 5 | 471.65 | <0.001 |
| Time | 1 | 431.84 | <0.001 | |
| Treatment xTime | 5 | 27.48 | <0.001 | |
| Error | 47 |
| Parameters | gl (Factor, Error) | Statistic F | p Value |
|---|---|---|---|
| Root Biomass (g) | (5, 30) | 5.34 | <0.001 |
| Leaf and Stem Biomass (g) | (5, 30) | 3.03 | <0.025 |
| Total Biomass (g/plant) | (5, 30) | 6.17 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan-Keb, C.A.; Agraz-Hernández, C.M.; Pérez-Balan, R.A.; Gutiérrez-Alcántara, E.J.; Muñiz-Salazar, R.; Reyes-Castellano, J.E.; Osti-Sáenz, J. Phytotoxicity in Seedlings of Rhizophora mangle (L.) Exposed to 2,4-Dichlorophenoxyacetic Acid under Experimental Conditions. J. Mar. Sci. Eng. 2021, 9, 1417. https://doi.org/10.3390/jmse9121417
Chan-Keb CA, Agraz-Hernández CM, Pérez-Balan RA, Gutiérrez-Alcántara EJ, Muñiz-Salazar R, Reyes-Castellano JE, Osti-Sáenz J. Phytotoxicity in Seedlings of Rhizophora mangle (L.) Exposed to 2,4-Dichlorophenoxyacetic Acid under Experimental Conditions. Journal of Marine Science and Engineering. 2021; 9(12):1417. https://doi.org/10.3390/jmse9121417
Chicago/Turabian StyleChan-Keb, Carlos A., Claudia M. Agraz-Hernández, Román A. Pérez-Balan, Eduardo J. Gutiérrez-Alcántara, Raquel Muñiz-Salazar, Jordán E. Reyes-Castellano, and Juan Osti-Sáenz. 2021. "Phytotoxicity in Seedlings of Rhizophora mangle (L.) Exposed to 2,4-Dichlorophenoxyacetic Acid under Experimental Conditions" Journal of Marine Science and Engineering 9, no. 12: 1417. https://doi.org/10.3390/jmse9121417