
Exploring Deep-Sea Biodiversity in the Porcupine Bank (NE Atlantic) through Fish Integrative Taxonomy

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
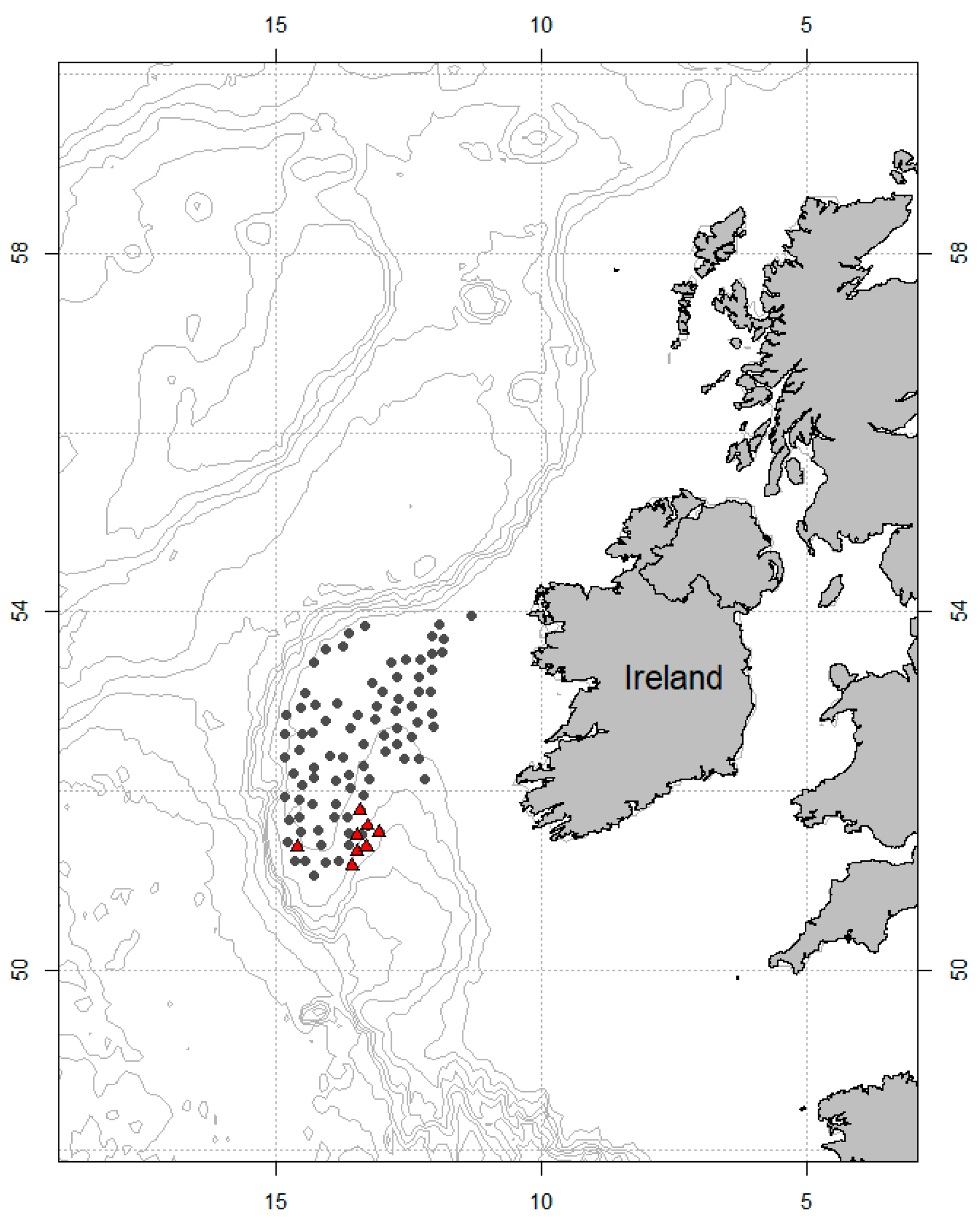
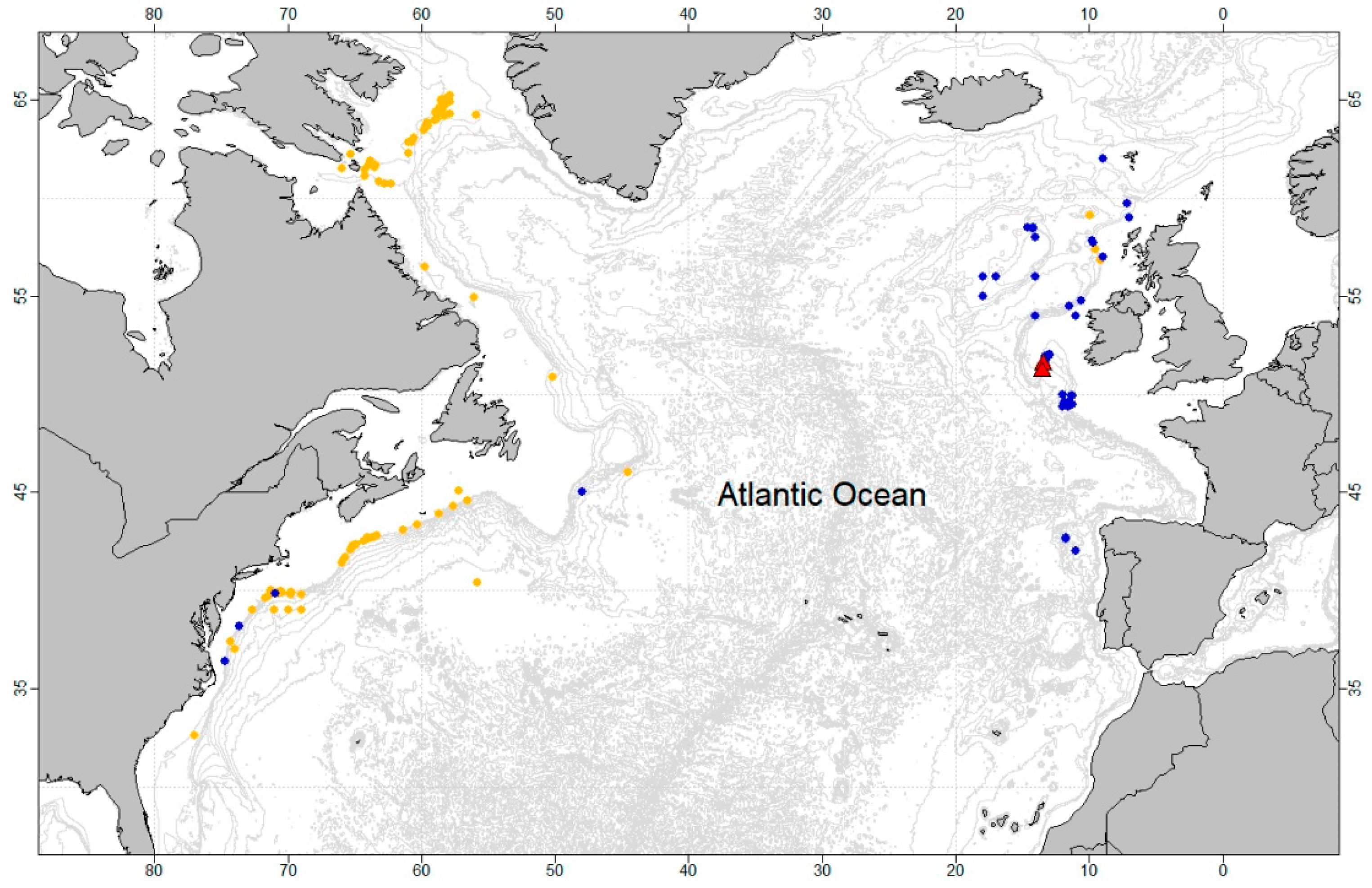
2.1. Study Site and Sampling
2.2. Morphological Analysis
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Molecular Analysis and Assignment of Specimens
3. Results
3.1. Morphological Traits
3.1.1. Nessorhamphus ingolfianus (Schmidt, 1912) (Anguilliformes: Derichthyidae)
3.1.2. Borostomias antarcticus (Lönnberg, 1905) (Stomiiformes: Stomiidae)
3.1.3. Scopelosaurus lepidus (Krefft and Maul, 1955) (Aulopiformes: Notosudidae)
3.1.4. Bathypterois dubius Vaillant, 1888 (Aulopiformes: Ipnopidae)
3.1.5. Evermannella balbo (Risso, 1820) (Aulopiformes: Evermannellidae)
3.1.6. Antimora rostrata (Günther, 1878) (Gadiformes: Moridae)
3.1.7. Melanonus zugmayeri Norman, 1930 (Gadiformes: Melanonidae)
3.1.8. Lyconus brachycolus Holt and Byrne, 1906 (Gadiformes: Lyconidae)
3.1.9. Paraliparis hystrix Merrett, 1983 (Scorpaeniformes: Liparidae)
3.1.10. Neocyttus helgae (Holt and Byrne, 1908) (Zeiformes: Oreosomatidae)
3.1.11. Platyberyx opalescens Zugmayer, 1911 (Perciformes: Caristiidae)
3.1.12. Howella atlantica Post and Quéro, 1991 (Perciformes: Howellidae)
3.1.13. Lycodes terraenovae Collett, 1896 (Perciformes: Zoarcidae)
3.1.14. Pseudoscopelus altipinnis Collett, 1896 (Perciformes: Chiasmodontidae)
3.2. NJ Trees and Genetic Distances
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Snelgrove, P.V.R. The deep-sea under global change. Curr. Biol. 2017, 27, R461–R465. [Google Scholar] [CrossRef] [Green Version]
- Glover, A.G.; Higgs, N.; Horton, T. World Register of Deep-Sea species (WoRDSS). Available online: http://www.marinespecies.org/deepsea (accessed on 15 February 2021). [CrossRef]
- Clark, M.R.; Althaus, F.; Williams, A.; Niklitschek, E.; Menezes, G.M.; Hareide, N.R.; Sutton, P.; O’Donnell, C. Are deep-sea demersal fish assemblages globally homogenous?—insights from seamounts. Mar. Ecol. 2010, 31, 39–51. [Google Scholar] [CrossRef]
- Henriques, M.; Murta, A.G.; das Neves, H. Melanonus zugmayeri Norman, 1930, captured off Portugal. A review of the current knowledge on this species. Sci. Mar. 2001, 65, 43–46. [Google Scholar] [CrossRef]
- Prokofiev, A.M. New findings of rare fish species in Indian and Pacific Oceans with the description of two new species from the families Gobiidae and Platycephalidae. J. Ichthyol. 2017, 57, 803–820. [Google Scholar] [CrossRef]
- Barros-García, D.; Froufe, E.; Bañón, R.; Arronte, J.C.; Baldó, F.; de Carlos, A. Phylogeography highlights two different Atlantic/Mediterranean lineages and a phenotypic latitudinal gradient for the deep-sea morid codling Lepidion lepidion (Gadiformes: Moridae). Deep Sea Res. Part I Oceanogr. Res. Pap. 2020, 157, 103212. [Google Scholar] [CrossRef]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- Bañón, R.; Ruiz-Pico, S.; Baldó, F.; de Carlos, A. Unexpected deep-sea fish species on the Porcupine Bank (NE Atlantic): Biogeographical implications. J. Fish Biol. 2020, 97, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Bañón, R.; Arronte, J.C.; Vázquez-Dorado, S.; del Río, J.L.; de Carlos, A. DNA barcoding of the genus Lepidion (Gadiformes: Moridae) with recognition of Lepidion eques as a junior synonym of Lepidion lepidion. Mol. Ecol. Resour. 2013, 13, 189–199. [Google Scholar] [CrossRef]
- Pires, A.C.; Marinoni, L. DNA barcoding and traditional taxonomy unified through integrative taxonomy: A view that challenges the debate questioning both methodologies. Biota Neotrop. 2010, 10, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Ardura, A.; Morote, E.; Kochzius, M.; García-Vázquez, E. Diversity of planktonic fish larvae along a latitudinal gradient in the Eastern Atlantic Ocean estimated through DNA barcodes. PeerJ 2016, 4, e2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, H.R.; Harris, W.E. An emergent science on the brink of irrelevance: A review of the past 8 years of DNA barcoding. Mol. Ecol. Resour. 2012, 12, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.M.; Friedman, S.T.; Corn, K.A.; Larouche, O.; Price, S.A.; Wainwright, P.C. The deep sea is a hot spot of fish body shape evolution. Ecol. Lett. 2021, 24, 1788–1799. [Google Scholar] [CrossRef] [PubMed]
- De Carlos, A.; Bañón, R.; Cobo-Arroyo, S.; Arronte, J.C.; Del Río, J.L.; Barros-García, D. DNA barcoding flags the existence of sympatric cryptic species in the slender codling Halargyreus johnsonii (Gadiformes, Moridae). Mar. Biodivers. 2020, 50, 49. [Google Scholar] [CrossRef]
- Johnston, G.; O’Hea, B.; Dransfeld, L. Fish species recorded during deepwater trawl surveys on the continental shelf and the Porcupine Bank, 2006–2008. Ir. Nat. J. 2010, 31, 130–134. [Google Scholar]
- O’Riordan, C.E. Some interesting fishes and other marine fauna from the Porcupine Bank. Ir. Nat. J. 1984, 21, 321–323. [Google Scholar]
- Ordines, F.; Fricke, R.; González, F.; Baldó, F. First record of Neoscopelus macrolepidotus Johnson, 1863 (Actinopterygii: Myctophiformes: Neoscopelidae) from Irish waters (Porcupine Bank, north-eastern Atlantic). Acta Ichthyol. Piscat. 2017, 47, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Howell, K.L.; Hilário, A.; Allcock, A.L.; Bailey, D.M.; Baker, M.; Clark, M.R.; Colaço, A.; Copley, J.; Cordes, E.E.; Danovaro, R.; et al. A Blueprint for an Inclusive, Global Deep-Sea Ocean Decade Field Program. Front. Mar. Sci. 2020, 7, 584861. [Google Scholar] [CrossRef]
- Howell, K.L.; Hilário, A.; Allcock, A.L.; Bailey, D.M.; Baker, M.; Clark, M.R.; Colaço, A.; Copley, J.; Cordes, E.E.; Danovaro, R.; et al. A decade to study deep-sea life. Nat. Ecol. Evol. 2021, 5, 265–267. [Google Scholar] [CrossRef]
- Porteiro, F.M.; Sutton, T.; Byrkjedal, I.; Orlov, A.M.; Heino, M.; Menezes, G.; Bergstad, O.A. Fishes of the Northern Mid-Atlantic Ridge Collected During the MAR-ECO Cruise in June-July 2004: An Annotated Checklist. Arquipelago 2017, 10, 1–126. Available online: https://nsuworks.nova.edu/occ_facreports/102 (accessed on 15 April 2021).
- Sutton, T.T.; Hulley, P.A.; Wienerroither, R.; Zaera-Perez, D.; Paxton, J.R. Identification Guide to the Mesopelagic Fishes of the Central and South East Atlantic Ocean; FAO Species Identification Guide for Fishery Purposes; FAO: Rome, Italy, 2020; pp. 1–356. [Google Scholar]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D.N. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Møller, P.R.; Gravlund, P. Phylogeny of the eelpout genus Lycodes (Pisces, Zoarcidae) as inferred from mitochondrial cytochrome B and 12S RDNA. Mol. Phylogen. 2003, 26, 369–388. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. PNAS 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Messing, J. New M13 vectors for cloning. Methods Enzymol. 1983, 101, 20–78. [Google Scholar] [CrossRef]
- Villesen, P. FaBox: An online toolbox for fasta sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics, 1st ed.; Oxford University Press: New York, NY, USA, 2000; pp. 1–333. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.J.; Steinke, D.; McMillan, P.J.; Stewart, A.L.; McVeagh, S.M.; Diaz de Astarloa, J.M.; Welsford, D.; Ward, R.D. DNA barcoding highlights a cryptic species of grenadier Macrourus in the Southern Ocean. J. Fish Biol. 2011, 78, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Bañón, R.; Arronte, J.C.; Rodríguez-Cabello, C.; Piñeiro, C.-G.; Punzón, A.; Serrano, A. Commented checklist of marine fishes from the Galicia Bank seamount (NW Spain). Zootaxa 2016, 4067, 293–333. [Google Scholar] [CrossRef] [Green Version]
- Chernova, N.V.; Stein, D.L.; Andriashev, A.P. Family Liparidae Scopoli 1777 snailfishes. Calif. Acad. Sci. Annot. Checkl. Fishes 2004, 31, 1–72. [Google Scholar]
- Kukuev, E.I.; Parin, N.V.; Trunov, I.A. Materials for the Revision of the Family Caristiidae (Perciformes). 2. Manefishes from the East Atlantic (Redescription of Platyberyx opalescens Zugmayer and Description of Two New Species Platyberyx mauli sp. n. and Caristius andriashevi sp. n.). J. Ichthyol. 2012, 52, 185–199. [Google Scholar] [CrossRef]
- Eduardo, L.N.; Villarins, B.T.; Martins, J.R.; Lucena Frédou, F.; Frédou, T.; Lira, A.S.; Travassos, P.E.; Bertrand, A.; Mincarone, M.M. Deep-sea oceanic basslets (Perciformes, Howellidae) from Brazil: New records and range extensions. Check List 2019, 15, 965–971. [Google Scholar] [CrossRef]
- Anderson, M.E. Zoarcidae. In The Living Marine Resources of the Eastern Central Atlantic, Bony Fishes Part 2 (Perciformes to Tetradontiformes) and Sea Turtles; Carpenter, K.E., De Angelis, N., Eds.; FAO: Rome, Italy, 2016; Volume 4, pp. 2759–2761. [Google Scholar]
- Melo, M.R.S. A revision of the genus Pseudoscopelus Lütken (Chiasmodontidae: Acanthomorphata) with descriptions of three new species. Zootaxa 2010, 2710, 1–78. [Google Scholar] [CrossRef]
- Barros-García, D.; Bañón, R.; Arronte, J.C.; Fernández-Peralta, L.; García, R.; Iglesias, S.; Sellos, D.; Barreiros, J.P.; Sebastián-Comesaña, A.; de Carlos, A. New insights into the systematics of North Atlantic Gaidropsarus (Gadiformes, Gadidae): Flagging synonymies and hidden diversity. Mar. Biol. Res. 2018, 14, 17–29. [Google Scholar] [CrossRef]
- Zemlak, T.S.; Ward, R.D.; Connell, H.D.; Holmes, B.H.; Hebert, P.D.N. DNA barcoding reveals overlooked marine fishes. Mol. Ecol. Resour. 2009, 9, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Burke, C.V. A new genus and six new species of fishes of the family Cyclogasteridae. Proc. United States Natl. Mus. 1912, 43, 567–574. [Google Scholar] [CrossRef]
- Burke, C.V. Revision of the fishes of the family Liparidae. Bull. United States Natl Mus. 1930, 150, 1–204. [Google Scholar] [CrossRef]
- Merrett, N.R. A new species of the deep-sea fish genus Paraliparis Collett (Liparididae) from the eastern North Atlantic, with notes on its ecology. J. Fish Biol. 1983, 23, 429–439. [Google Scholar] [CrossRef]
- Steinke, D.; Zemlak, T.S.; Boutillier, J.A.; Hebert, P.D.N. DNA barcoding of Pacific Canada’s fishes. Mar. Biol. 2009, 156, 2641–2647. [Google Scholar] [CrossRef]
- Hartel, K.E.; Kenaley, C.P.; Galbraith, J.K.; Sutton, T.T. Additional Records of Deep-sea Fishes from off Greater New England. Northeast. Nat. 2008, 15, 317–334. [Google Scholar] [CrossRef] [Green Version]
- GBIF Occurrence. Available online: https://www.gbif.org/es/ (accessed on 15 April 2021).
- OBIS. Available online: www.iobis.org (accessed on 15 April 2021).
- Stevenson, D.E.; Kenaley, C.P. Revision of the Manefish genera Caristius and Platyberyx (Teleostei: Percomorpha: Caristiidae), with descriptions of five new species. Copeia 2013, 2013, 415–434. [Google Scholar] [CrossRef]
- Nazarkin, M.V. Morphology of two eastern pacific eelpouts (Pisces, Zoarcidae, Lycodes) in connection with features of distribution of species with a double lateral line. J. Ichthyol. 2011, 51, 209–216. [Google Scholar] [CrossRef]
- Nielsen, J.G.; Fossa, S.V. Lycodes adolfi, a new species of eelpout (Zoarcidae) from Greenland. Cybium 1993, 17, 39–44. [Google Scholar]
- Møller, P.R. Identity of the Atlantic eelpouts Lycodes terraenovae Collett, 1896, L. atlanticus Jensen, 1902 and L. agulhensis Andriashev, 1959 (Pisces, Zoarcidae). Steenstrupia 1997, 22, 45–58. [Google Scholar]
- Quéro, J.C.; Lorance, P.; Tardy, J. Observations ichtyologiques effectuées en 1999 à bord du navire de recherche “Thalassa”. Annal. Soc. Sci. Nat. Charente-Marit. 2000, 8, 1047–1058. [Google Scholar]
- Coleman, C.O.; Radulovici, A.E. Challenges for the future of taxonomy: Talents, databases and knowledge growth. Megataxa 2020, 1, 28–34. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Taxonomic ID | GenBank Acc. No. | BLASTn ID | Query Cover (%) | E-Value | Identity (%) | Accession |
|---|---|---|---|---|---|---|
| Antimora rostrata | MW907994 | A. rostrata | 100 | 0.0 | 100 | JF265134 |
| Bathypterois dubius | MW907995 | B. dubius | 99 | 0.0 | 99.85 | KC015247 |
| Borostomias antarcticus | MW907998 | B. antarcticus | 100 | 0.0 | 99.85 | KF929668 |
| Borostomias antarcticus | MW907996 | B. antarcticus | 100 | 0.0 | 99.85 | KF929668 |
| Borostomias antarcticus | MW907999 | B. antarcticus | 100 | 0.0 | 99.69 | KF929668 |
| Borostomias antarcticus | MW907997 | B. antarcticus | 100 | 0.0 | 99.54 | KY033872 |
| Evermannella balbo | MW908000 | E. balbo | 99 | 0.0 | 99.85 | KY033618 |
| Howella atlantica | MW908001 | H. brodiei | 100 | 0.0 | 99.69 | EU148199 |
| Lycodes terraenovae | MW908002 | L. adolfi | 100 | 0.0 | 99.69 | GU804885 |
| Lyconus brachycolus | MW908003 | L. brachycolus | 99 | 0.0 | 98.92 | EU148230 |
| Lyconus brachycolus | MW908004 | L. brachycolus | 99 | 0.0 | 98.38 | EU148230 |
| Melanonus zugmayeri | MW908005 | M. zugmayeri | 100 | 0.0 | 99.39 | EU148249 |
| Melanonus zugmayeri | MW908007 | M. zugmayeri | 100 | 0.0 | 99.85 | EU148249 |
| Melanonus zugmayeri | MW908006 | M. zugmayeri | 100 | 0.0 | 99.69 | EU148249 |
| Neocyttus helgae | MW908008 | N. helgae | 100 | 0.0 | 100 | EU148264 |
| Nessorhamphus ingolfianus | MW908009 | N. ingolfianus | 100 | 0.0 | 100 | EU148266 |
| Nessorhamphus ingolfianus | MW908010 | N. ingolfianus | 100 | 0.0 | 100 | EU148266 |
| Paraliparis hystrix | MW908014 | Paraliparis sp. | 99 | 0.0 | 94.93 | KX676118 |
| Paraliparis hystrix | MW908011 | Paraliparis sp. | 99 | 0.0 | 95.08 | KX676118 |
| Paraliparis hystrix | MW908012 | Paraliparis sp. | 100 | 0.0 | 95.09 | KX676118 |
| Paraliparis hystrix | MW908013 | Paraliparis sp. | 100 | 0.0 | 95.09 | KX676118 |
| Platyberyx opalescens | MW908015 | P. maderensis | 96 | 0.0 | 99.84 | EU148108 |
| Pseudoscopelus altipinnis | MW908016 | P. altipinnis | 100 | 0.0 | 98.93 | MT323772 |
| Pseudoscopelus altipinnis | MW908017 | P. altipinnis | 100 | 0.0 | 98.77 | MT323772 |
| Scopelosaurus lepidus | MW908018 | S. lepidus | 100 | 0.0 | 99.39 | KF930385 |
| Paraliparis garmani [40,41] | Paraliparis hystrix [42] | |
|---|---|---|
| Contained in standard length | ||
| Head length | 4.5 | 4.2–5.7 |
| Body depth | 4.6 | 3.9–5.1 |
| Contained in head length | ||
| Eye diameter | 5.4 | 2.3–3.4 |
| Snout length | – | 2.2–3.4 |
| Gill slit length | 4.2 | 4.7 |
| Upper pectoral lobe length | 2 | 1.3–1.8 |
| Counts | ||
| Dorsal fin rays | 54 | 51–60 |
| Upper pectoral fin rays | 14 | 11–15 |
| Notch fin rays | 4 | 2–5 |
| Lower pectoral fin rays | 3–4 | 2–3 |
| Total pectoral fin rays | 21 | 16–21 |
| Caudal fin rays | 8 | 7–8 |
| Pyloric caeca | 6 | 4–8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bañón, R.; de Carlos, A.; Farias, C.; Vilas-Arrondo, N.; Baldó, F. Exploring Deep-Sea Biodiversity in the Porcupine Bank (NE Atlantic) through Fish Integrative Taxonomy. J. Mar. Sci. Eng. 2021, 9, 1075. https://doi.org/10.3390/jmse9101075
Bañón R, de Carlos A, Farias C, Vilas-Arrondo N, Baldó F. Exploring Deep-Sea Biodiversity in the Porcupine Bank (NE Atlantic) through Fish Integrative Taxonomy. Journal of Marine Science and Engineering. 2021; 9(10):1075. https://doi.org/10.3390/jmse9101075
Chicago/Turabian StyleBañón, Rafael, Alejandro de Carlos, Carlos Farias, Nair Vilas-Arrondo, and Francisco Baldó. 2021. "Exploring Deep-Sea Biodiversity in the Porcupine Bank (NE Atlantic) through Fish Integrative Taxonomy" Journal of Marine Science and Engineering 9, no. 10: 1075. https://doi.org/10.3390/jmse9101075