
Shifts in the Assemblage of Summer Mesopelagic Fish Larvae in the Gaoping Waters of Southwestern Taiwan: A Comparison between El Niño Events and Regular Years

Abstract
:1. Introduction
2. Materials and Methods
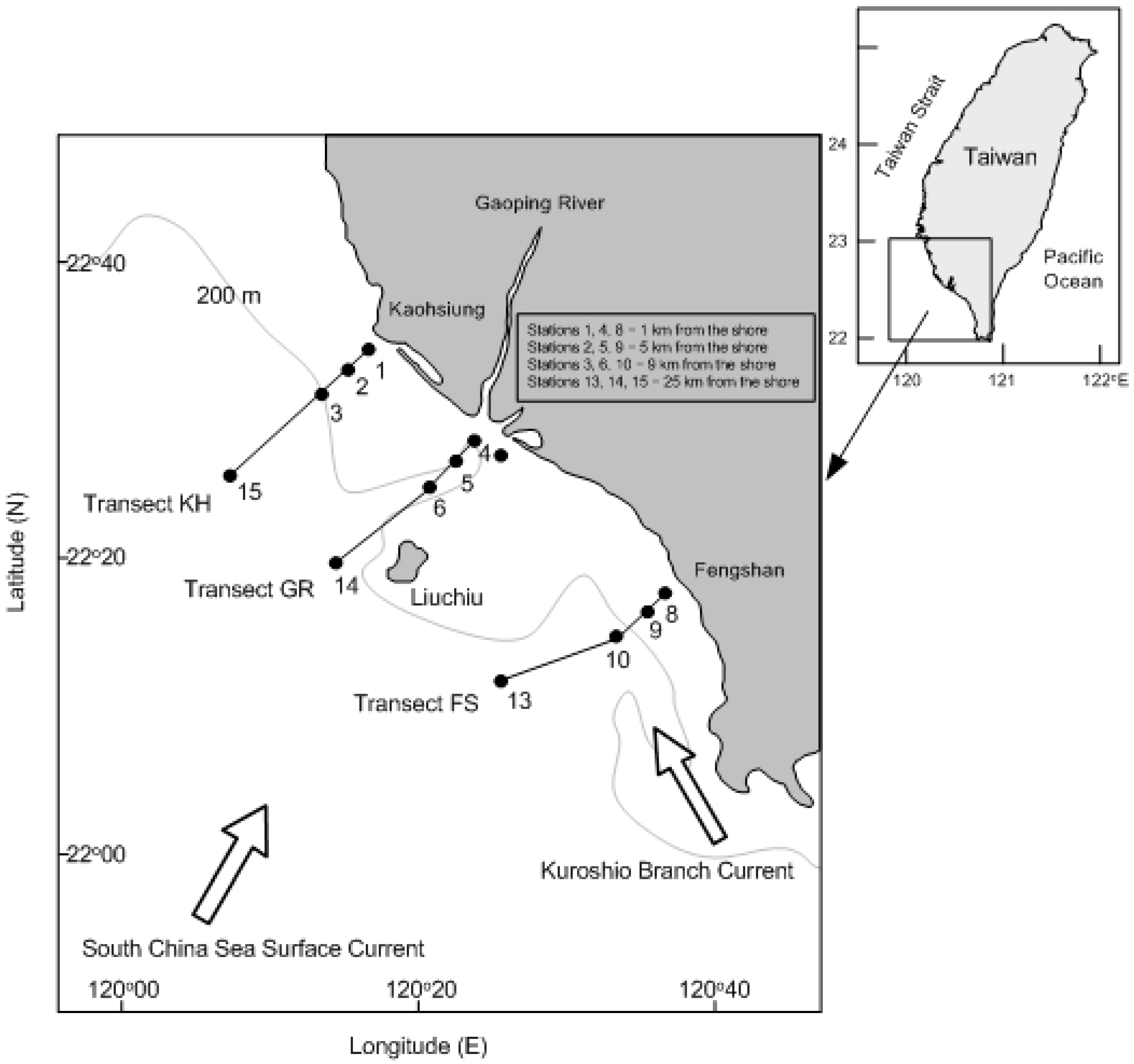
2.1. Field Sampling Work
2.2. Laboratory Work
2.3. Data Analysis
3. Results
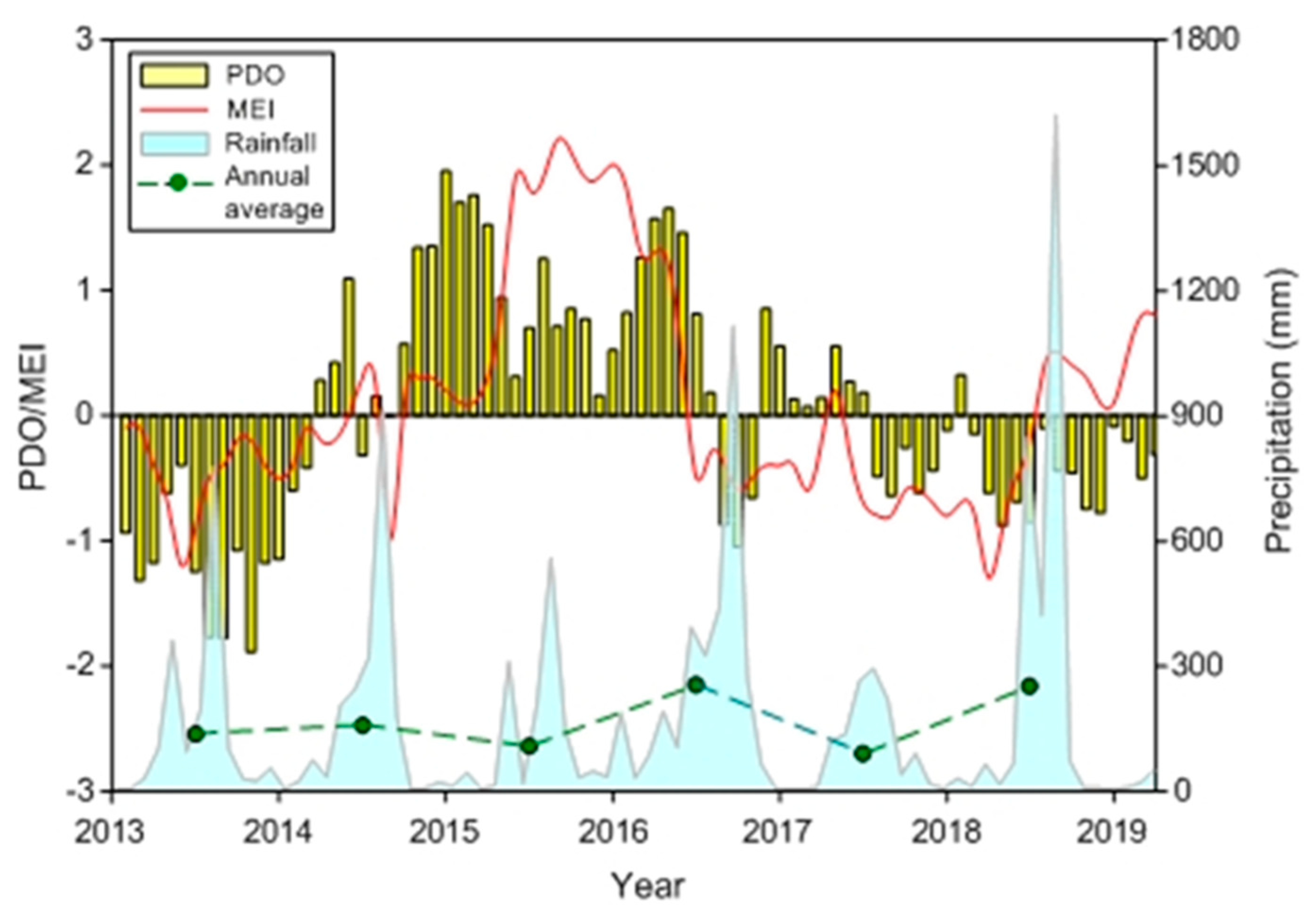
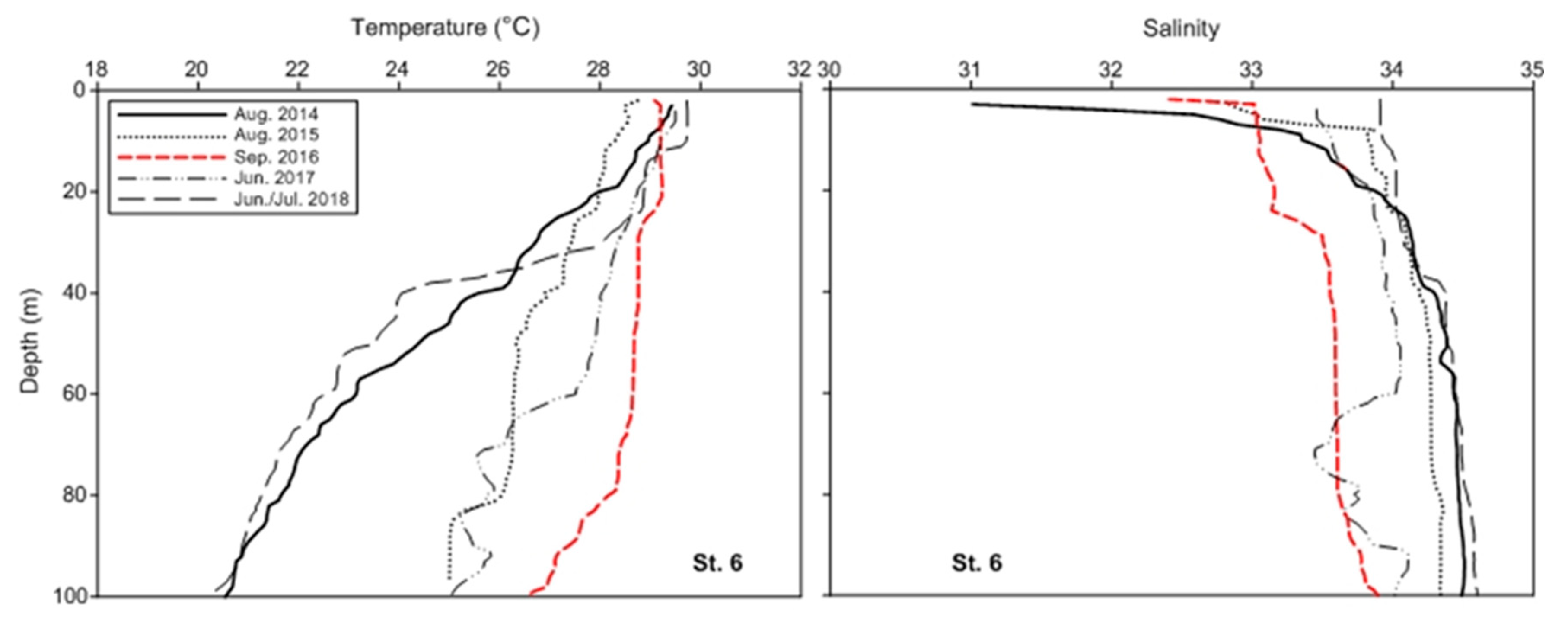
3.1. Climatic and Hydrographic Conditions
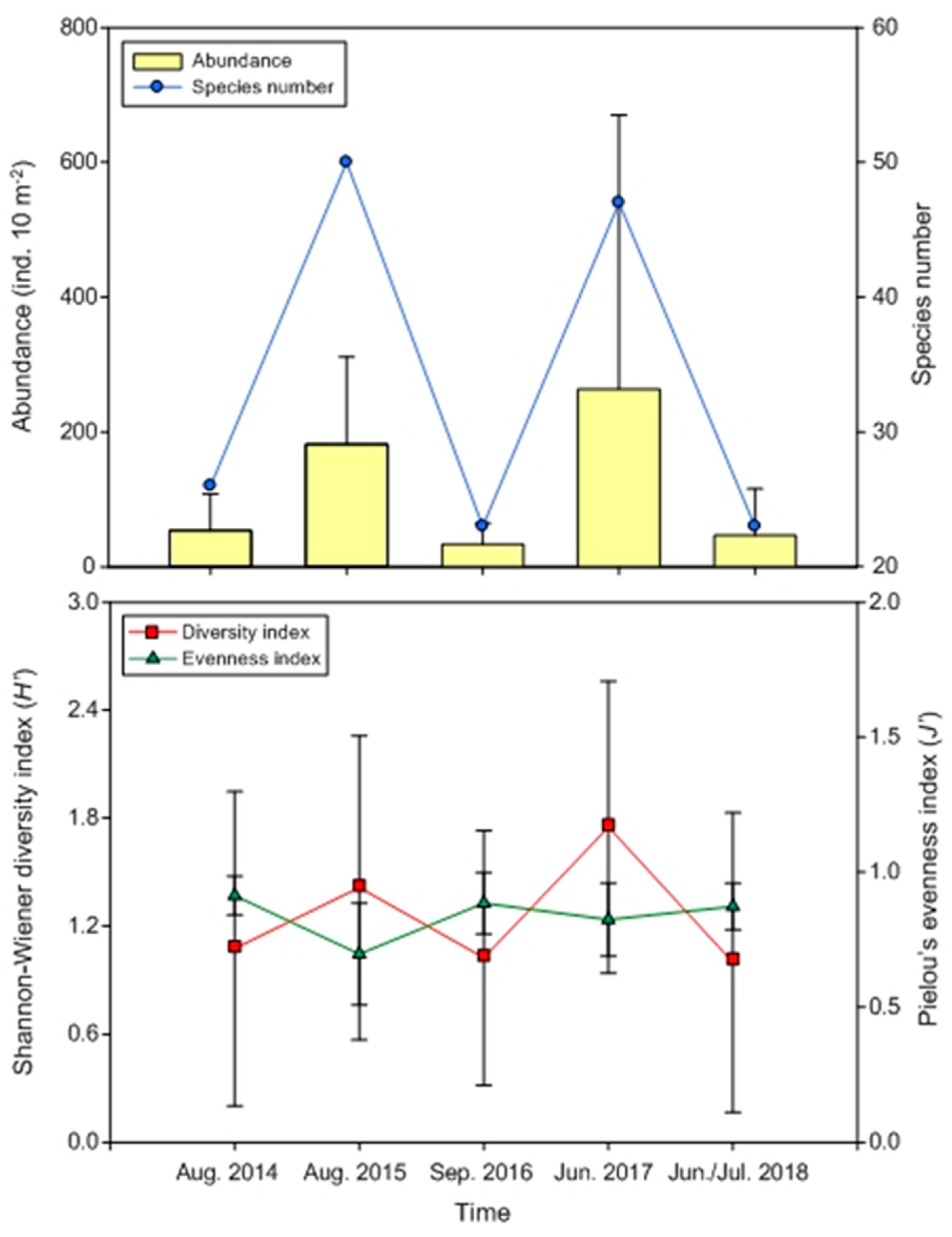
3.2. Biological Variables
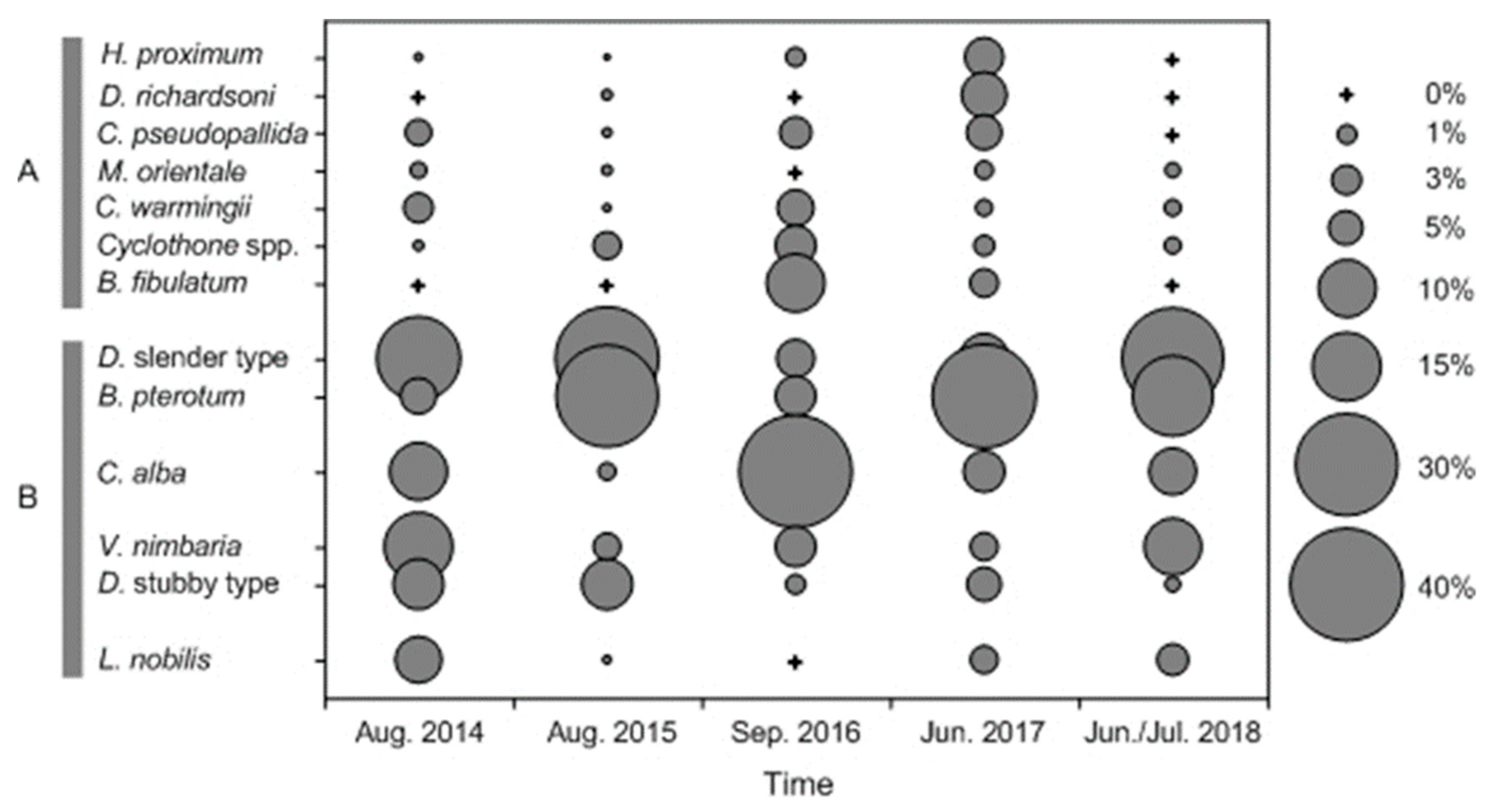
3.3. Composition of Summer Mesopelagic Fish Larvae
3.4. Interannual Differences in Abundance and Diversity of Summer Mesopelagic Fish Larvae
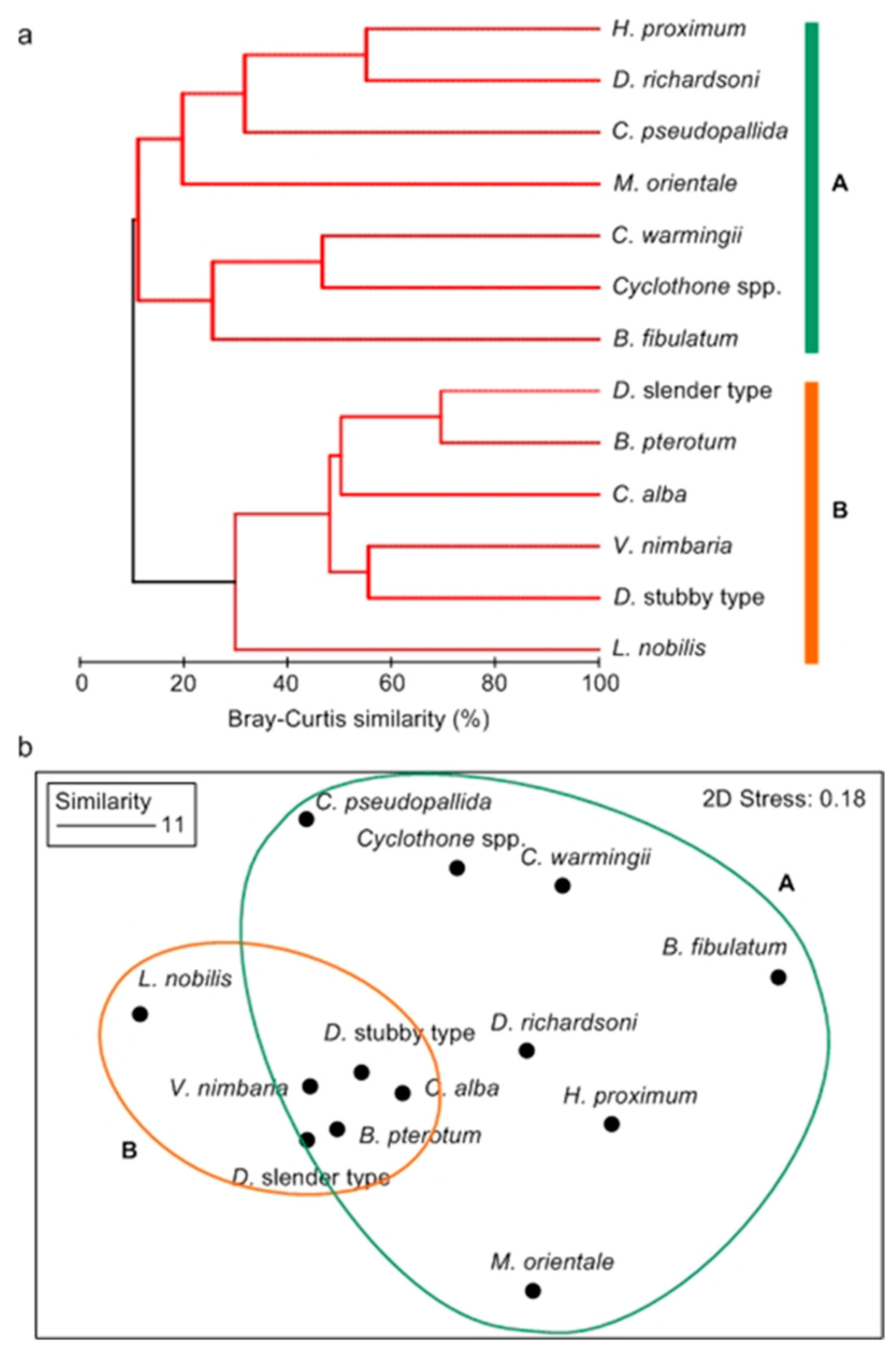
3.5. Species Groups of Summer Mesopelagic Fish Larvae
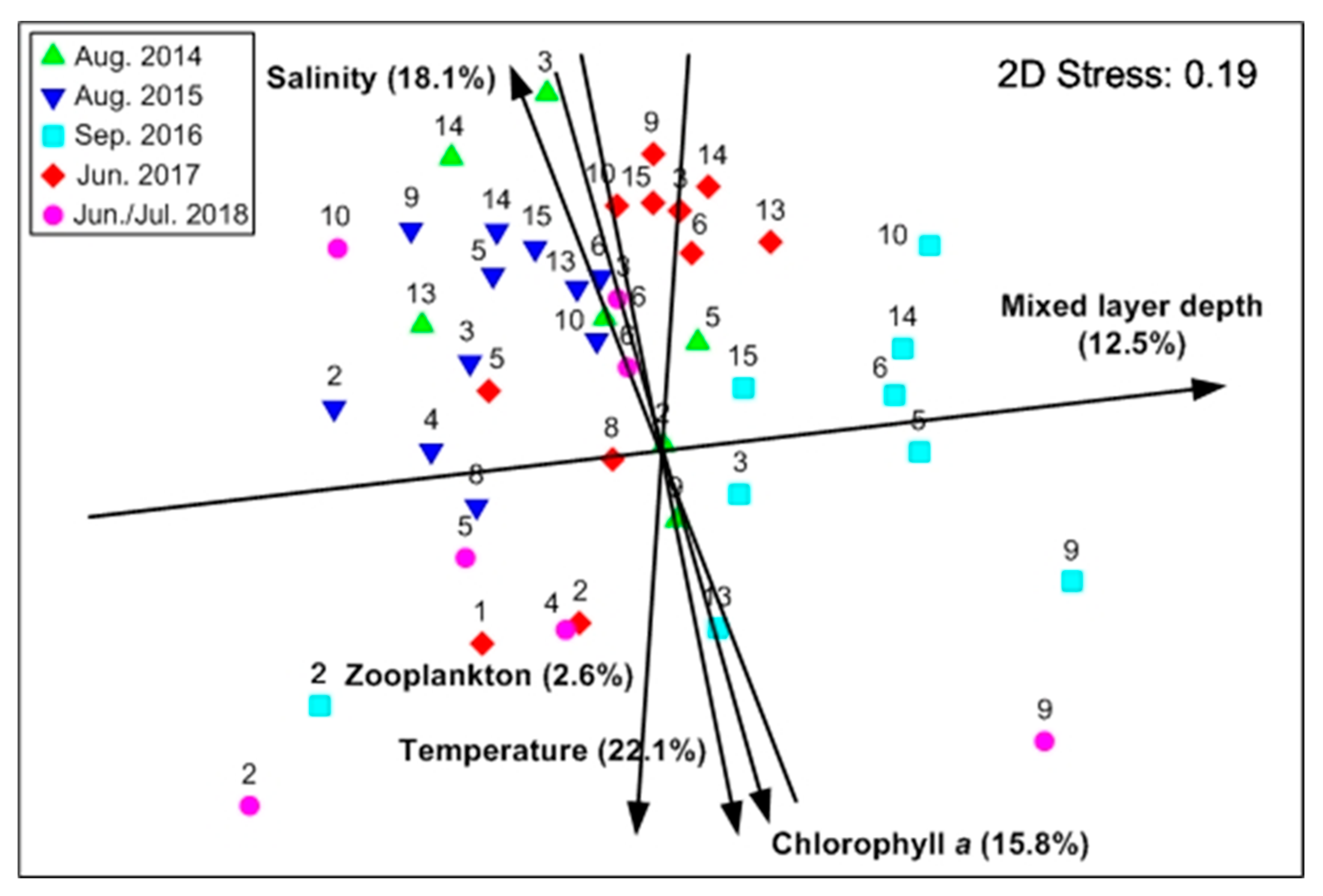
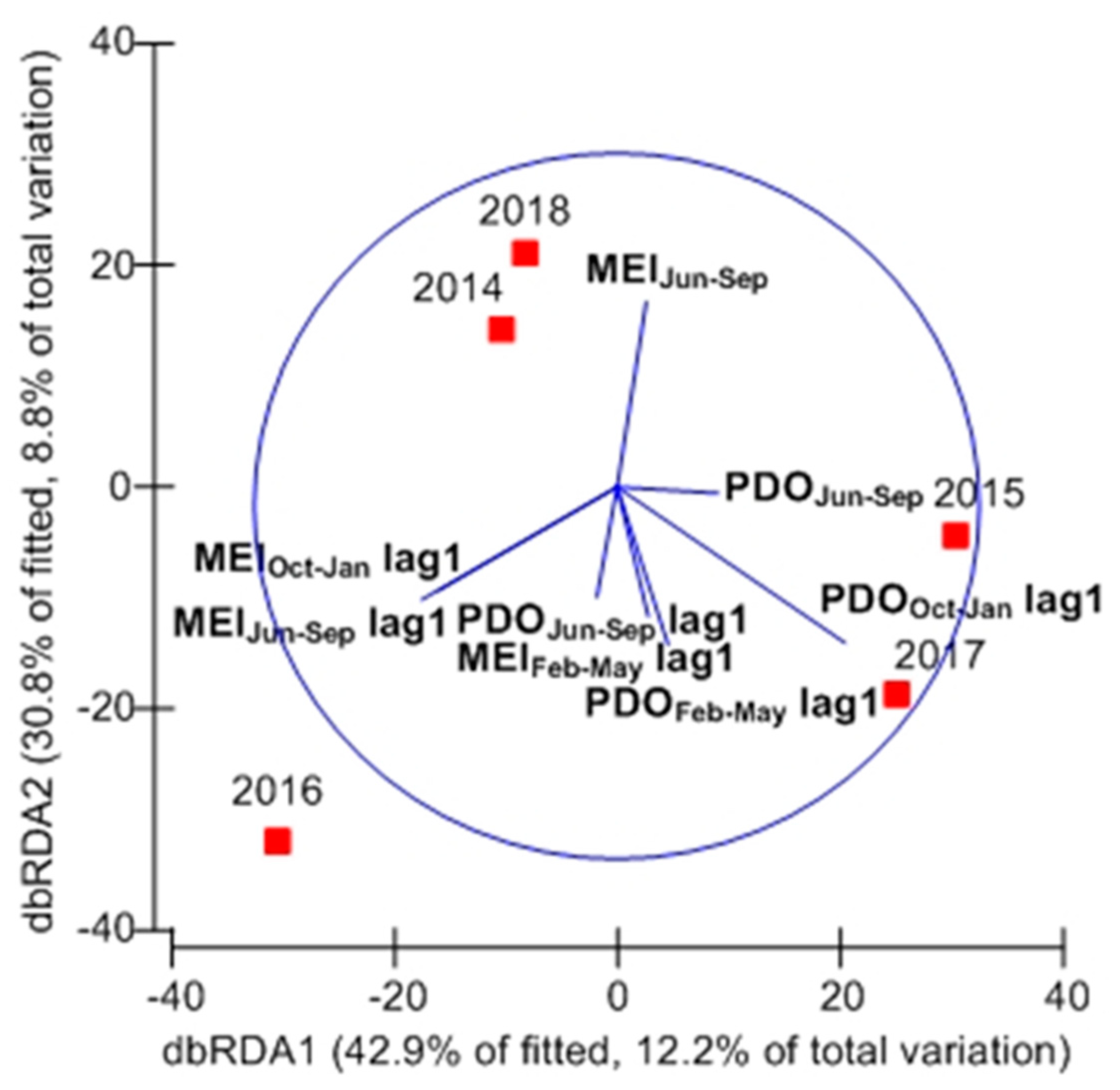
3.6. Summer Mesopelagic Fish Larvae Assemblage in Relation to Environmental Variables
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Sassa, C.; Kawaguchi, K.; Kinoshita, T.; Watanabe, C. Assemblages of vertical migratory mesopelagic fish in the transitional region of the western North Pacific. Fish. Oceanogr. 2002, 11, 193–204. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Irigoien, X.; Klevjer, T.A.; Røstad, A.; Martinez, U.; Boyra, G.; Acuña, J.L.; Bode, A.; Echevarria, F.; Gonzalez-Gordillo, J.I.; Hernandez-Leon, S.; et al. Large mesopelagic fishes biomass and trophic efficiency in the open ocean. Nat. Commun. 2014, 5, 3271–3280. [Google Scholar] [CrossRef] [Green Version]
- FAO. FAO Yearbook: Fishery and Aquaculture Statistics. Capture Production 2011; FAO: Rome, Italy, 2013. [Google Scholar]
- Sabatés, A. Diel vertical distribution of fish larvae during the winter-mixing period in the Northwestern Mediterranean. ICES J. Mar. Sci. 2004, 61, 1243–1252. [Google Scholar] [CrossRef]
- Sassa, C.; Hirota, Y. Seasonal occurrence of mesopelagic fish larvae on the onshore side of the Kuroshio off southern Japan. Deep Sea Res. Part I Oceanogr. Res. Pap. 2013, 81, 49–61. [Google Scholar] [CrossRef]
- Sassa, C.; Kawaguchi, K.; Hirota, Y.; Ishida, M. Distribution patterns of larval myctophid fish assemblages in the subtropical-tropical waters of the western North Pacific. Fish. Oceanogr. 2004, 13, 267–282. [Google Scholar] [CrossRef]
- Olivar, M.P.; Bernal, A.; Molí, B.; Peña, M.; Balbín, R.; Castellón, A.; Miquel, J.; Massutí, E. Vertical distribution, diversity and assemblages of mesopelagic fishes in the western Mediterranean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2012, 62, 53–69. [Google Scholar] [CrossRef]
- Williams, A.; Koslow, J.A.; Terauds, A.; Haskard, K. Feeding ecology of five fishes from the mid-slope micronekton community off southern Tasmania, Australia. Mar. Biol. 2001, 139, 1177–1192. [Google Scholar]
- Cherel, Y.; Ducatez, S.; Fontaine, C.; Richard, P.; Guinet, C. Stable isotopes reveal the trophic position and mesopelagic fish diet of female southern elephant seals breeding on the Kerguelen Islands. Mar. Ecol. Prog. Ser. 2008, 370, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Davison, P.C.; Checkley, D.M.; Koslow, J.A.; Barlow, J. Carbon export mediated by mesopelagic fishes in the northeast Pacific Ocean. Prog. Oceanogr. 2013, 116, 14–30. [Google Scholar] [CrossRef]
- Young, P.C.; Leis, J.M.; Hausfeld, H.F. Seasonal and spatial distribution of fish larvae in waters over the North West Continental Shelf of Western Australia. Mar. Ecol. Prog. Ser. 1986, 31, 209–222. [Google Scholar] [CrossRef]
- Doyle, M.J.; Morse, W.W.; Kendall, A.W., Jr. A comparison of larval fish assemblages in the temperate zone of the northeast Pacific and northwest Atlantic Oceans. Bull. Mar. Sci. 1993, 53, 588–644. [Google Scholar]
- Olivar, M.P.; Emelianov, M.; Villate, F.; Uriarte, I.; Maynou, F.; Álvarez, I.; Morote, E. The role of oceanographic conditions and plankton availability in larval fish assemblages off the Catalan coast (NW Mediterranean). Fish. Oceanogr. 2010, 19, 209–229. [Google Scholar] [CrossRef]
- Olivar, M.P.; Beckley, L.E. Influence of the Agulhas Current on the distribution of lanterfish larvae off southeast coast of Africa. J. Plankton Res. 1994, 16, 1759–1780. [Google Scholar] [CrossRef]
- Sabatés, A.; Olivar, M.P.; Salat, J.; Palomera, I.; Alemany, F. Physical and biological processes controlling the distribution of fish larvae in the NW Mediterranean. Prog. Oceanogr. 2007, 74, 355–376. [Google Scholar] [CrossRef] [Green Version]
- Moser, H.G.; Smith, P.E. Larval fish assemblages and oceanic boundaries. Bull. Mar. Sci. 1993, 53, 283–289. [Google Scholar]
- Shepherd, F.G.; Cushing, D.H. A mechanism for density-dependent survival of larval fish as the basis of a stock-recruitment relationship. ICES J. Mar. Sci. 1980, 39, 160–167. [Google Scholar] [CrossRef]
- Takahashi, M.; Watanabe, Y. Growth rate-dependent recruitment of Japanese Anchovy Engraulis japonicus in the Kuroshio-Oyashio transitional waters. Mar. Ecol. Prog. Ser. 2004, 266, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Cheung, W.L.; Lam, V.Y.; Sarmiento, J.L.; Kearney, K.; Watson, R.; Pauly, D. Projecting global marine biodiversity impacts under climate change scenarios. Fish Fish. 2009, 10, 235–251. [Google Scholar] [CrossRef]
- Drinkwater, K.F.; Beaugrand, G.; Kaeriyama, M.; Kim, S.; Ottersen, G.; IanPerry, R.; Pörtner, H.-O.; Polovina, J.J.; Takasukaj, A. On the processes linking climate to ecosystem changes. J. Mar. Syst. 2010, 79, 374–388. [Google Scholar] [CrossRef]
- Ganias, K.; Somarakis, S.; Machias, A.; Koutsikopoulos, C. Factors affecting the spawning period of sardine in two highly oligotrophic Seas. Mar. Biol. 2007, 151, 1559–1569. [Google Scholar] [CrossRef]
- Stratoudakis, Y.; Coombs, S.; de Lanzós, A.L.; Halliday, N.; Costas, G.; Caneco, B.; Franco, C.; Conway, D.; Santos, M.B.; Silva, A.; et al. Sardine (Sardina pilchardus) spawning seasonality in European waters of the northeast Atlantic. Mar. Biol. 2007, 152, 201–212. [Google Scholar] [CrossRef]
- Bernal, M.; Stratoudakis, Y.; Coombs, S.; Angelico, M.M.; de Lanzós, A.L.; Porteiro, C.; Sagarminaga, Y.; Santos, M.; Uriarte, A.; Cunha, E.; et al. Sardine spawning off the European Atlantic coast: Characterization of and spatio-temporal variability in spawning habitat. Prog. Oceanogr. 2007, 74, 210–227. [Google Scholar] [CrossRef]
- Planque, B.; Bellier, E.; Lazure, P. Modelling potential spawning habitat of sardine (Sardina pilchardus) and anchovy (Engraulis encrasicolus) in the Bay of Biscay. Fish. Oceanogr. 2007, 16, 16–30. [Google Scholar] [CrossRef]
- Somarakis, S.; Palomera, I.; Garcia, A.; Quintanilla, L.; Koutsikopoulos, C.; Uriarte, A.; Motos, L. Daily egg production of anchovy in European waters. ICES J. Mar. Sci. 2004, 61, 944–958. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.-H.; Reiss, C.; Watson, W.; Allen, M.J.; Hunter, J.R.; Lea, R.N.; Rosenblatt, R.H.; Smith, P.E.; Sugihara, G. A comparison of long-term trends and variability in populations of larvae of exploited and unexploited fishes in the Southern California region: A community approach. Prog. Oceanogr. 2005, 67, 160–185. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Chen, C.S.; Chiu, T.S.; Lee, K.T.; Shieh, F.J.; Pan, J.Y.; Lee, M.A. Time series analyses reveal transient relationships between abundance of larval anchovy and environmental variables in the coastal waters southwest of Taiwan. Fish. Oceanogr. 2009, 18, 102–117. [Google Scholar] [CrossRef]
- Busby, M.S.; Duffy-Anderson, J.T.; Mier, K.L.; de Forest, L.G. Spatial and temporal patterns in summer ichthyoplankton assemblages on the eastern Bering Sea shelf 1996–2007. Fish. Oceanogr. 2014, 23, 270–287. [Google Scholar] [CrossRef]
- Guan, L.; Dower, J.F.; Mckinnell, S.M.; Pepin, P.; Pakhomov, E.A.; Hunt, B.P.V. A comparison of spring larval fish assemblages in the Strait of Georgia (British Columbia, Canada) between the early 1980s and late 2000s. Prog. Oceanogr. 2015, 138, 45–57. [Google Scholar] [CrossRef]
- Jan, S.; Wang, J.; Chern, C.S.; Chao, S.Y. Seasonal variation of the circulation in the Taiwan Strait. J. Mar. Syst. 2002, 35, 249–268. [Google Scholar] [CrossRef]
- Jan, S.; Sheu, D.D.; Kuo, H.M. Water mass and throughflow transport variability in the Taiwan Strait. J. Geophys. Res. 2006, 111, C12012. [Google Scholar] [CrossRef]
- Hsieh, H.Y.; Lo, W.T.; Chen, H.H.; Meng, P.J. Larval Fish Assemblages and Hydrographic Characteristics in the Coastal Waters of Southwestern Taiwan during Non- and Post-typhoon Summers. Zool. Stud. 2016, 55, 18. [Google Scholar]
- Hsieh, H.Y.; Meng, P.J.; Chang, Y.C.; Lo, W.T. Temporal and spatial occurrence of mesopelagic fish larvae during epipelagic drift associated with hydrographic features in the Gaoping coastal waters off southwestern Taiwan. Mar. Coast. Fish. 2017, 9, 244–259. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.A.; Lee, K.T.; Shiah, G.Y. Environmental factors associated with the formation of larval anchovy fishing ground in coastal waters of southwestern Taiwan. Mar. Biol. 1995, 121, 621–625. [Google Scholar] [CrossRef]
- Tsai, C.F.; Chen, P.Y.; Chen, C.P.; Lee, M.A.; Shiah, G.Y.; Lee, K.T. Fluctuation in abundance of larval anchovy and environmental conditions in coastal waters off southwestern Taiwan as associated with the El Niño-Southern Oscillation. Fish. Oceanogr. 1997, 6, 238–249. [Google Scholar] [CrossRef]
- Sassa, C.; Konishi, Y. Late winter larval fish assemblage in the southern East China Sea, with emphasis on spatial relations between mesopelagic and commercial pelagic fish larvae. Cont. Shelf Res. 2015, 108, 97–111. [Google Scholar] [CrossRef]
- Sassa, C.; Takahashi, M. Comparative larval growth and mortality of mesopelagic fishes and their predatory impact on zooplankton in the Kuroshio region. Deep Sea Res. Part I Oceanogr. Res. Pap. 2018, 131, 121–132. [Google Scholar] [CrossRef]
- Leis, J.M.; Rennis, D.S. The Larvae of Indo-Pacific Coral Reef Fishes; New South Wales University Press: Sidney, Australia, 1983. [Google Scholar]
- Ozawa, T. Studies on the Oceanic Ichthyoplankton in the Western North Pacific; Kyushu University Press: Fukuoka, Japan, 1986. [Google Scholar]
- Okiyama, M. (Ed.) An Atlas of the Early Stage Fishes in Japan; Tokai University Press: Tokyo, Japan, 1988. (In Japanese) [Google Scholar]
- Leis, J.M.; Trnski, T. The Larvae of Indo-Pacific Shorefishes; New South Wales University Press: Sidney, Australia, 1989. [Google Scholar]
- Neira, F.J.; Miskiewicz, A.G.; Trnski, T. Larvae of Temperate Australian Fishes; University of Western Australia Press: Perth, Australia, 1998. [Google Scholar]
- Omori, M.; Ikeda, T. Methods in Marine Zooplankton Ecology; Wiley: New York, NY, USA, 1984. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Dunn, O.J.; Clark, V.A. Applied Statistics: Analysis of Variance and Regression; John Wiley: New York, NY, USA, 1974. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Kruskal, J.B.; Wish, M. Multidimensional Scaling; Sage University Paper Series on Quantitative Application in the Social Sciences; Sage Publications: Beverly Hills, CA, USA, 1978. [Google Scholar]
- Hosie, G.W.; Cochran, T.G. Mesoscale distribution patterns of macrozooplankton communities in Prydz bay, Antarctica—January to February 1991. Mar. Ecol. Prog. Ser. 1994, 106, 21–39. [Google Scholar] [CrossRef]
- Somarakis, S.; Drakopoulos, P.; Filippou, V. Distribution and abundance of larval fishes in the northern Aegean Sea—Eastern Mediterranean—In relation to early summer oceanographic conditions. J. Plankton Res. 2002, 24, 339–357. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Aust. Ecol. 2001, 26, 32–46. [Google Scholar]
- Mantua, N.J.; Hare, S.R.; Zhang, Y.; Wallace, J.M.; Francis, R.C. A Pacific interdecadal climate oscillation with impacts on salmon production. Bull. Am. Meteorol. Soc. 1997, 78, 1069–1079. [Google Scholar] [CrossRef]
- Wolter, K.; Timlin, M.S. Measuring the strength of ENSO events: How does 1997/98 rank? Weather 1998, 53, 315–324. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A common on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Chen, J.M.; Wang, F.J.; Lu, F.C.; Kuo, S.L. El Nifño and 1998 Climate Variability of Taiwan: Persistent Warming and Excessive Spring Rains. Atmos. Sci. 2002, 30, 331–349. [Google Scholar]
- Wang, B.; Wu, R.G.; Fu, X.H. Pacific-East Asian teleconnection: How does ENSO affect East Asian climate? J. Clim. 2000, 13, 1517–1536. [Google Scholar] [CrossRef]
- Kuo, N.J.; Ho, C.R. ENSO effect on the sea surface wind and sea surface temperature in the Taiwan Strait. Geophys. Res. Lett. 2004, 31, L13309. [Google Scholar] [CrossRef]
- Schwing, F.B.; Murphree, T.; Green, P.M. The Northern Oscillation Index (NOI): A new climate index for the northeast Pacific. Prog. Oceanogr. 2002, 53, 115–119. [Google Scholar] [CrossRef]
- Lee, M.A.; Kuo, Y.C.; Chan, J.W.; Chen, Y.K.; Teng, S.Y. Long-term (1982–2012) summertime sea surface temperature variability in the Taiwan Strait. Terr. Atmos. Ocean. Sci. 2015, 26, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Yan, X.H.; Zheng, Q.; Liu, W.T.; Klemas, V.V. Interpretation of scatterometer ocean surface wind vector EOFs over the Northwestern Pacific. Remote Sens. Environ. 2002, 84, 53–68. [Google Scholar] [CrossRef]
- García-Comas, C.; Stemmann, L.; Ibanez, F.; Berline, L.; Mazzocchi, M.G.; Gasparini, S.; Picheral, M.; Gorsky, G. Zooplankton long-term changes in the NW Mediterranean Sea: Decadal periodicity forced by winter hydrographic conditions related to large-scale atmospheric changes? J. Mar. Syst. 2011, 87, 216–226. [Google Scholar] [CrossRef]
- Feng, J.; Stige, L.C.; Durant, J.M.; Hessen, D.O.; Zhu, L.; Hjermann, D.O.; Llope, M.; Stenseth, N.C. Large-scale season-dependent effects of temperature and zooplankton on phytoplankton in the North Atlantic. Mar. Ecol. Prog. Ser. 2014, 502, 25–37. [Google Scholar] [CrossRef]
- Lo, W.T.; Yu, S.F.; Hsieh, H.Y. Hydrographic processes driven by seasonal monsoon system affect siphonophore assemblages in tropical-subtropical waters (western North Pacific Ocean). PLoS ONE 2014, 9, e100085. [Google Scholar] [CrossRef] [Green Version]
- Marques, S.C.; Primo, A.L.; Martinho, F.; Azeiteiro, U.M.; Pardal, M.Â. Shifts in estuarine zooplankton variability following extreme climate events: A comparison between drought and regular years. Mar. Ecol. Prog. Ser. 2014, 499, 65–76. [Google Scholar] [CrossRef]
- Tseng, H.-C.; You, W.-L.; Huang, W.; Chung, C.-C.; Tsai, A.-Y.; Chen, T.-Y.; Lan, K.-W.; Gong, G.-C. Seasonal Variations of Marine Environment and Primary Production in the Taiwan Strait. Front. Mar. Sci. 2020, 7, 38. [Google Scholar] [CrossRef]
- Auth, T.D.; Brodeur, R.D.; Soulen, H.L.; Ciannelli, L.; Peterson, W.T. The response of fish larvae to decadal changes in environmental forcing factors off the Oregon coast. Fish. Oceanogr. 2011, 20, 314–328. [Google Scholar] [CrossRef]
- Raya, V.; Sabatés, A. Diversity and distribution of early life stages of carangid fishes in the northwestern Mediterranean: Responses to environmental drivers. Fish. Oceanogr. 2015, 24, 118–134. [Google Scholar] [CrossRef]
- Ottersen, G.; Stenseth, N.C. Atlantic climate governs oceanographic and ecological variability in the Barents Sea. Limnol. Oceanogr. 2001, 46, 1774–1780. [Google Scholar] [CrossRef]
- Colton, J.B., Jr. A field observation of mortality of marine fish larvae due to warming. Limnol. Oceanogr. 1959, 4, 219–222. [Google Scholar] [CrossRef]
- McGowen, G.E. Coastal ichthyoplankton assemblages, with emphasis on the southern California bight. Bull. Mar. Sci. 1993, 53, 692–722. [Google Scholar]
- Sanvicente-Añorve, L.; Flores-Coto, C.; Chiappa-Carrara, X. Temporal and spatial scales of ichthyoplankton distribution in the southern Gulf of Mexico. Estuar. Coast. Shelf Sci. 2000, 51, 463–475. [Google Scholar] [CrossRef]
- Rodriguez, J.M.; Hernandez-Leon, S.; Barton, E.D. Vertical distribution of fish larvae in the Canaries-African coastal transition zone in summer. Mar. Biol. 2006, 149, 885–897. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, H.; Sassa, C.; Ishida, M. Late winter vertical distribution of mesopelagic fish larvae in the Kuroshio Current region of the western North Pacific. Bull. Jpn. Soc. Fish. Oceanogr. 2010, 74, 153–158. [Google Scholar]
- Beaugrand, G.; Brander, C.M.; Lindley, A.J.; Souissi, S.; Reid, P.C. Plankton effect on cod recruitment in the North Sea. Nature 2003, 426, 661–664. [Google Scholar] [CrossRef]
- Frank, K.T.; Petrie, B.; Shackell, N.L.; Choi, J.S. Reconciling differences in trophic control in mid-latitude marine ecosystems. Ecol. Lett. 2006, 9, 1096–1105. [Google Scholar] [CrossRef]
- Perry, R.I.; Schweigert, J. Primary productivity and the carrying capacity for herring in NE Pacific marine ecosystems. Prog. Oceanogr. 2008, 77, 241–251. [Google Scholar] [CrossRef]
- DiLorenzo, E.; Schneider, N.; Cobb, K.M.; Chhak, K.; Franks, P.J.S.; Miller, A.J.; McWilliams, J.C.; Bograd, S.J.; Arango, H.; Curchitser, E.; et al. North Pacific Gyre Oscillation links ocean climate and ecosystem change. Geophys. Res. Lett. 2008, 35, L08607. [Google Scholar]
- Mackas, D.; Galbraith, M.; Faust, D.; Masson, D.; Young, K.; Shaw, W.; Romaine, S.; Trudel, M.; Dower, J.; Campbell, R.; et al. Zooplankton time series from the Strait of Georgia: Results from year-round sampling at deep water locations, 1990–2010. Prog. Oceanogr. 2013, 115, 129–159. [Google Scholar] [CrossRef]
- Sassa, C.; Tsukamoto, Y. Distribution and growth of Scomber japonicas and S. australasicus larvae in the southern East China Sea in response to oceanographic conditions. Mar. Ecol. Prog. Ser. 2010, 419, 185–199. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Aug. 2014 | Aug. 2015 | Sep. 2016 | Jun. 2017 | Jun./Jul. 2018 |
|---|---|---|---|---|---|
| Precipitation (mm) | 902 | 548 | 1098 | 256 | 835/410 |
| Mixed layer depth | 18 | 25 | 79 | 28 | 24 |
| Temperature (°C) | 24.52 ± 2.90 | 26.69 ± 1.10 | 28.50 ± 0.67 | 27.32 ± 1.43 | 24.46 ± 3.36 |
| Salinity | 34.15 ± 0.57 | 34.14 ± 0.31 | 33.48 ± 0.26 | 33.82 ± 0.20 | 34.32 ± 0.23 |
| Chlorophyll a (μg L−1) | 0.20 ± 0.20 | 0.14 ± 0.03 | 0.26 ± 0.14 | 0.17 ± 0.07 | 0.37 ± 0.47 |
| Zooplankton (individuals m−3) | 648 | 624 | 402 | 302 | 2037 |
| Family/Taxon | Aug. 2014 | Aug. 2015 | Sep. 2016 | Jun. 2017 | Jun./Jul. 2018 | N (Count of Individuals) | Percentage of the Total Catch (%) |
|---|---|---|---|---|---|---|---|
| Gonostomatidae | |||||||
| Cyclothone alba | 6.2 | 1.3 | 14.9 | 15.7 | 3.6 | 115 | 7.2 |
| C. pseudopallida | 1.8 | 0.7 | 1.0 | 8.9 | – | 40 | 2.2 |
| Cyclothone spp. | 0.4 | 3.7 | 2.2 | 3.2 | 0.5 | 30 | 1.7 |
| Myctophidae | |||||||
| Benthosema fibulatum | – | – | 3.2 | 9.1 | – | 14 | 2.1 |
| B. pterotum | 1.7 | 60.1 | 2.2 | 105.5 | 9.8 | 290 | 31.2 |
| Ceratoscopelus warmingii | 0.8 | 0.8 | 1.3 | 2.4 | 0.5 | 19 | 1.0 |
| Diaphus richardsoni | – | 1.2 | – | 17.4 | – | 47 | 3.3 |
| Diaphus slender type | 12.2 | 64.5 | 1.8 | 22.5 | 15.4 | 287 | 19.9 |
| Diaphus stubby type | 5.4 | 16.1 | 0.6 | 10.9 | 0.5 | 81 | 5.9 |
| Hygophum proximum | 0.3 | 0.5 | 0.6 | 12.5 | – | 24 | 2.4 |
| Lampanyctus nobilis | 5.1 | 0.8 | – | 4.7 | 1.7 | 29 | 2.1 |
| Myctophum orientale | 0.8 | 1.2 | – | 4.3 | 0.5 | 15 | 1.2 |
| Phosichthyidae | |||||||
| Vinciguerria nimbaria | 8.8 | 4.3 | 1.3 | 6.8 | 5.1 | 60 | 4.4 |
| Variable | X | Y | R2 | F | P |
|---|---|---|---|---|---|
| Temperature | 0.14 | −0.99 | 0.221 | 5.947 | 0.005 ** |
| Salinity | −0.41 | 0.91 | 0.181 | 4.648 | 0.015 * |
| Mixed layer depth | 0.98 | 0.18 | 0.125 | 3.011 | 0.060 |
| Chlorophyll a | 0.23 | −0.97 | 0.158 | 3.941 | 0.027 * |
| Zooplankton | −0.09 | −0.99 | 0.026 | 0.563 | 0.574 |
| Variables | SS (Trace) | Pseudo-F | P | % Variation Explained |
|---|---|---|---|---|
| PDOJun.-Sep. | 1337.5 | 0.796 | 0.702 | 2.09 |
| PDOFeb.-May × lag1 (lag of up to one year) | 1497.6 | 0.920 | 0.570 | 2.34 |
| PDOOct.-Jan. × lag1 | 1591.7 | 0.997 | 0.549 | 2.49 |
| PDOJun.-Sep. × lag1 | 1467.5 | 0.896 | 0.655 | 2.30 |
| MEIJun.-Sep. | 1398.4 | 0.842 | 0.591 | 2.19 |
| MEIFeb.-May × lag1 | 1454.5 | 0.886 | 0.551 | 2.28 |
| MEIOct.-Jan. × lag1 | 1965.3 | 1.336 | 0.245 | 3.08 |
| MEIJun.-Sep. × lag1 | 2004.9 | 1.375 | 0.249 | 3.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, H.-Y.; Lo, W.-T.; Liao, C.-C.; Meng, P.-J. Shifts in the Assemblage of Summer Mesopelagic Fish Larvae in the Gaoping Waters of Southwestern Taiwan: A Comparison between El Niño Events and Regular Years. J. Mar. Sci. Eng. 2021, 9, 1065. https://doi.org/10.3390/jmse9101065
Hsieh H-Y, Lo W-T, Liao C-C, Meng P-J. Shifts in the Assemblage of Summer Mesopelagic Fish Larvae in the Gaoping Waters of Southwestern Taiwan: A Comparison between El Niño Events and Regular Years. Journal of Marine Science and Engineering. 2021; 9(10):1065. https://doi.org/10.3390/jmse9101065
Chicago/Turabian StyleHsieh, Hung-Yen, Wen-Tseng Lo, Chien-Chun Liao, and Pei-Jie Meng. 2021. "Shifts in the Assemblage of Summer Mesopelagic Fish Larvae in the Gaoping Waters of Southwestern Taiwan: A Comparison between El Niño Events and Regular Years" Journal of Marine Science and Engineering 9, no. 10: 1065. https://doi.org/10.3390/jmse9101065