Environmental Influence on the Spatiotemporal Variability of Spawning Grounds in the Western Guangdong Waters, South China Sea

Abstract
:1. Introduction
2. Materials and Methods
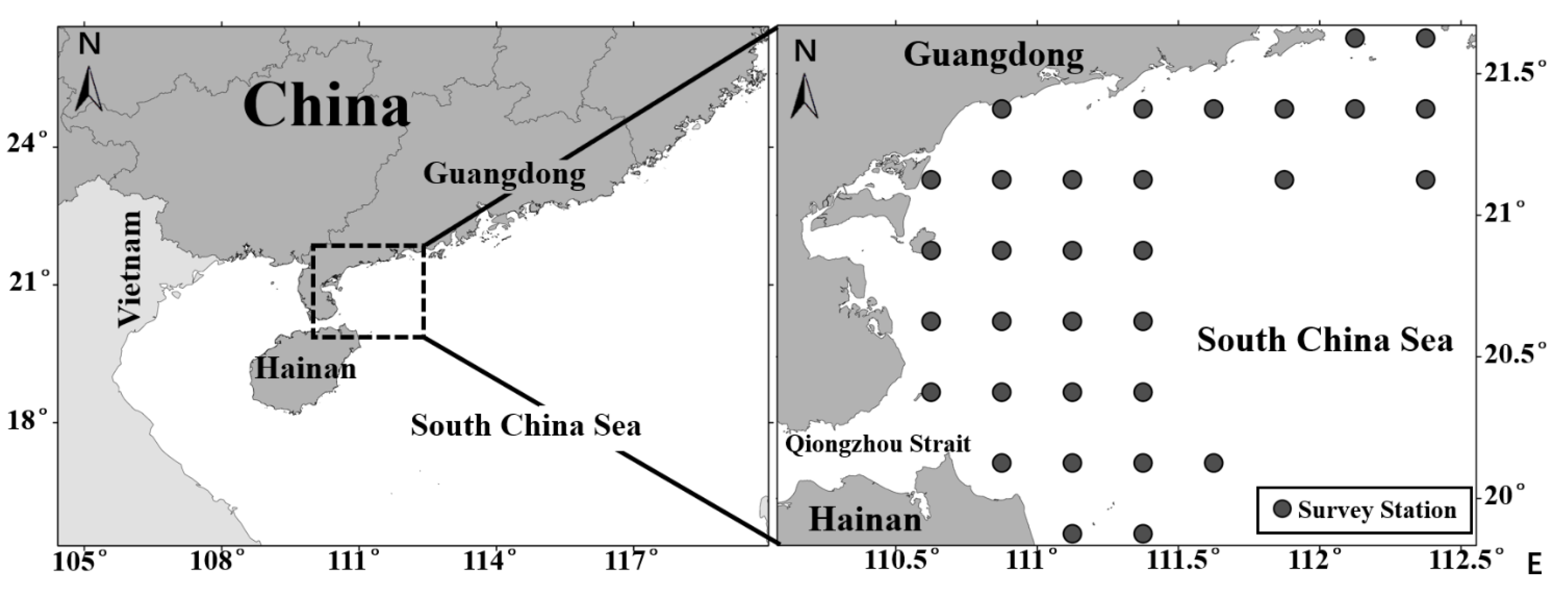
2.1. Fishery Data
2.2. Environmental Data
2.3. GAMs Fitting Procedures
2.4. Center of Gravity of Spawning Grounds
3. Results
3.1. GAMs Analysis
3.2. Relationship between Fish-egg Distribution and Environmental Factors
3.3. Relationship between Fish-egg Distribution and Spatial Factors
3.4. CoSGG Variations of Spawning Grounds in WGWs
4. Discussion
4.1. Effects of Environmental Factors on Fish-Egg Density
4.2. Effects of Spatial Factors on Fish-Egg Density
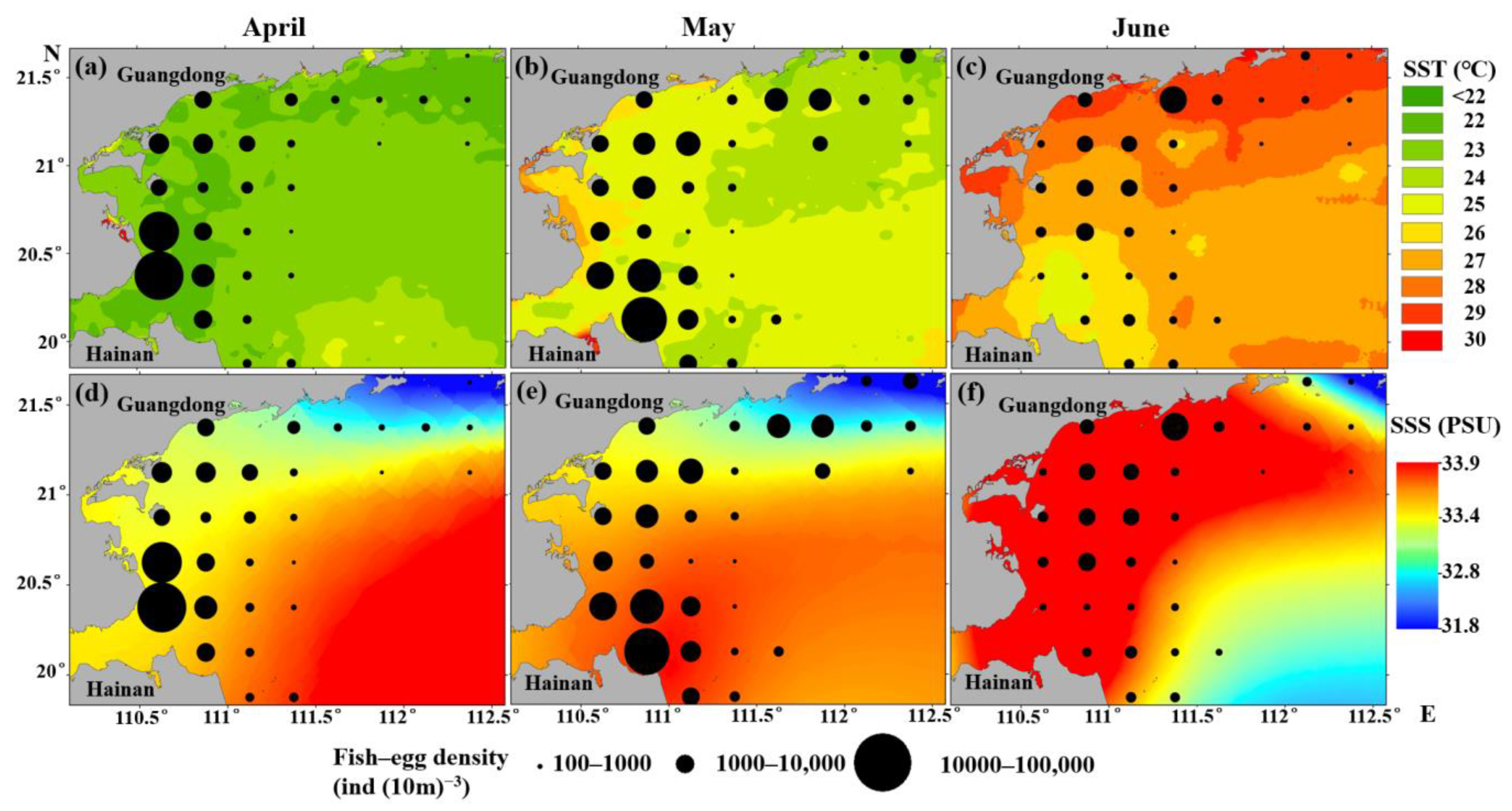
4.3. Spatiotemporal Distribution of Fish-Egg Density in WGWs
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yu, J.; Hu, Q.W.; Tang, D.L.; Chen, P.M. Environmental effects on the spatiotemporal variability of purpleback flying squid in Xisha-Zhongsha waters, South China Sea. Mar. Ecol. Prog. Ser. 2019, 623, 25–37. [Google Scholar] [CrossRef]
- Wang, D.X.; Hong, B.; Gan, J.P.; Xu, H.Z. Numerical investigation on propulsion of the counter-wind current in the northern South China Sea in winter. Deep Sea Res. Part I Oceanogr. Res. Pap. 2010, 57, 1206–1221. [Google Scholar] [CrossRef]
- Hovenkamp, F. Growth-dependent mortality of larval plaice Pleuronectes platessa in the North Sea. Mar. Ecol. Prog. Ser. 1992, 82, 95–101. [Google Scholar] [CrossRef]
- Houde, E.D.; Hoyt, R.D. Fish early life dynamics and recruitment variability. In Proceedings of the 10 Annual Larval Fish Conference, Miami, FL, USA, 18–23 May 1986. [Google Scholar]
- Peck, M.A.; Huebert, K.B.; Llopiz, J.K. Intrinsic and extrinsic factors driving match–mismatch dynamics during the early life history of marine fishes. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 2012; Volume 47, pp. 177–302. [Google Scholar]
- Xiao, Y.Z.; Wang, R.; Qiang, O.; Fang, H.D. Relationship between abundance distribution of fish eggs, larvae and juveniles and environmental factors in the Pearl River Estuary waters in spring. J. Appl. Oceanogr. 2010, 29, 488–495. [Google Scholar]
- Blood, D.M. Low-temperature incubation of walleye pollock (Theragra chalcogramma) eggs from the southeast Bering Sea shelf and Shelikof Strait, Gulf of Alaska. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 6095–6108. [Google Scholar] [CrossRef]
- Bernal, M.; Somarakis, S.; Witthames, P.R.; Van Damme, C.J.; Uriarte, A.; Lo, N.C.; Dickey-Collas, M. Egg production methods in marine fisheries: An introduction. Fish Res. 2012, 117, 1–5. [Google Scholar] [CrossRef]
- Lin, Z.J.; Wang, X.H.; Jiang, Y.E. Distribution and species composition of fish eggs in Daya Bay. J. Fish. Sci. China 2010, 17, 543–550. [Google Scholar]
- Wan, R.; Zhou, F.; Shan, X.; Sun, S. Impacts of variability of habitat factors on species composition of ichthyoplankton and distribution of fish spawning ground in the Changjiang River estuary and its adjacent waters. Acta Ecol. Sin. 2010, 30, 155–165. [Google Scholar] [CrossRef]
- Stenseth, N.C.; Mysterud, A.; Ottersen, G.; Hurrell, J.W.; Chan, K.; Lima, M. Ecological effects of climate fluctuations. Science 2002, 297, 1292–1296. [Google Scholar] [CrossRef] [Green Version]
- Hastie, T.; Tibshirani, R. Generalized Additive Models; Chapman and Hall: London, UK, 1990; pp. 587–602. [Google Scholar]
- Yu, J.; Hu, Q.W.; Tang, D.L.; Zhao, H.; Chen, P.M. Response of Sthenoteuthis oualaniensis to marine environmental changes in the north-central South China Sea based on satellite and in situ observations. PLoS ONE 2019, 14, e211474. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Liu, Z.N.; Chen, P.M.; Yao, L.J. Environmental factors affecting the spatiotemporal distribution of Decapterus maruadsi in the western Guangdong waters, China. Appl. Ecol. Environ. Res. 2019, 17. [Google Scholar] [CrossRef]
- Wang, Y.F.; Hu, Q.W.; Yu, J.; Chen, P.M.; Shu, L.M. Effect assessment of fishery resources proliferation in Zhelin Bay marine ranching in eastern Guangdong. South China Fish. Sci. 2019, 15, 12–19. [Google Scholar]
- Stratoudakis, Y.; Gallego, A.; Morrison, J.A. Spatial distribution of developmental egg ages within a herring Clupea harengus spawning ground. Mar. Ecol. Prog. Ser. 1998, 174, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Ustups, D.; Müller-Karulis, B.; Bergstrom, U.; Makarchouk, A.; Sics, I. The influence of environmental conditions on early life stages of flounder (Platichthys flesus) in the central Baltic Sea. J. Sea Res. 2013, 75, 77–84. [Google Scholar] [CrossRef]
- Schismenou, E.; Giannoulaki, M.; Valavanis, V.D.; Somarakis, S. Modeling and predicting potential spawning habitat of anchovy (Engraulis encrasicolus) and round sardinella (Sardinella aurita) based on satellite environmental information. Hydrobiologia 2008, 612, 201. [Google Scholar] [CrossRef]
- Li, Z.G. Distribution of Main Species of Stow Net in the South Yellow Sea based on GAM and Preliminary Study of Characteristics of Ichthyoplankton Assemblages in Haizhou Bay; Ocean University of China: Qingdao, China, 2013. [Google Scholar]
- Zhang, R.Y.; Bian, X.D.; Shan, X.J.; Jin, X.S.; Guan, L.S. Distribution of the age 0 group Pacific cod (Gadus macrocephalus) in the Yellow Sea and its relationship with environmental factors. J. Fish. China 2018, 42, 870–880. [Google Scholar] [CrossRef]
- Yu, J.; Chen, P.M.; Tang, D.L.; Qin, C.X. Ecological effects of artificial reefs in Daya Bay of China observed from satellite and in situ measurements. Adv. Space Res. 2015, 55, 2315–2324. [Google Scholar] [CrossRef]
- Müller, D. Estimation of algae concentration in cloud covered scenes using geostatistical methods. In Proceedings of the ENVISAT Symposium, Montreux, Switzerland, 23–27 April 2007. [Google Scholar]
- Yu, J.; Hu, Q.W.; Yuan, H.R.; Chen, P.M. Effect assessment of summer fishing moratorium in Daya Bay based on remote sensing data. South China Fish. Sci. 2018, 14, 1–9. [Google Scholar]
- Wang, Y.F.; Yu, J.; Chen, P.M.; Yu, J.; Liu, Z.N. Relationship between spatial-temporal distribution of light falling-net fishing ground and marine environments. J. Trop. Oceanogr. 2019, 38, 68–76. [Google Scholar]
- Wood, S.N. Stable and efficient multiple smoothing parameter estimation for generalized additive models. J. Am. Stat. Assoc. 2004, 99, 673–686. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Venables, W.N.; Dichmont, C.M. GLMs, GAMs and GLMMs: An overview of theory for applications in fisheries research. Fish. Res. 2004, 69, 319–337. [Google Scholar] [CrossRef]
- Hilborn, R.; Mangel, M. The Ecological Detective: Confronting Models with Data; Princeton University Press: Princeton, NJ, USA, 1997. [Google Scholar]
- Quinn, T.J.; Deriso, R.B. Quantitative Fish Dynamics; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Stone, C.J. Additive regression and other nonparametric models. Ann. Stat. 1985, 13, 689–705. [Google Scholar] [CrossRef]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Breakthroughs in Statistics; Springer: New York, NY, USA, 1992; pp. 610–624. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model. Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Chen, G.B.; Li, Y.Z. Distribution of the Carangidae fishes in the continental shelf waters of northern South China Sea. J. Shanghai Ocean Univ. 2003, 12, 146–151. [Google Scholar] [CrossRef]
- La Mesa, M.; La Mesa, G.; Catalano, B.; Jones, C.D. Spatial distribution pattern and physical–biological interactions in the larval notothenioid fish assemblages from the Bransfield Strait and adjacent waters. Fish. Oceanogr. 2016, 25, 624–636. [Google Scholar] [CrossRef]
- Bacha, M.; Jeyid, M.A.; Vantrepotte, V.; Dessailly, D.; Amara, R. Environmental effects on the spatio-temporal patterns of abundance and distribution of Sardina pilchardus and sardinella off the Mauritanian coast (North-West Africa). Fish. Oceanogr. 2017, 26, 282–298. [Google Scholar] [CrossRef]
- Singh, A.K.; Hasnain, S.I.; Banerjee, D.K. Grain size and geochemical partitioning of heavy metals in sediments of the Damodar River—A tributary of the lower Ganga, India. Environ. Geol. 1999, 39, 90–98. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, P.; Chen, Z.Z.; Chen, G.B.; Zhang, K.; Xue, Y.W.; Sun, M.S. Biomass and distribution of carangoid fish resources in the offshore South China Sea. South China Fish. Sci. 2016, 12, 38–48. [Google Scholar]
- Giffard-Mena, I.; Hernández-Montiel, Á.H.; Pérez-Robles, J.; David-True, C. Effects of salinity on survival and plasma osmolarity of Totoaba macdonaldi eggs, larvae, and juveniles. J. Exp. Mar. Biol. Ecol. 2020, 526, 151339. [Google Scholar] [CrossRef]
- Lin, J.Q. Studies on the natural regulaive adaptibility of the Hairtail (Trichiurus haumela) in the oceanic environment. Trans. Oceanol. Limnol. 1981, 3, 60–65. [Google Scholar]
- Zhang, R.Z.; Lu, S.F. On the eggs and larvae of the golden-thread Nemipterus Virgatus (Houttuyn). Curr. Zool. 1980, 26, 39–42. [Google Scholar]
- Nissling, A.; Nyberg, S.; Petereit, C. Egg buoyancy of flounder, Platichthys flesus, in the Baltic Sea—Adaptation to salinity and implications for egg survival. Fish. Res. 2017, 191, 179–189. [Google Scholar] [CrossRef]
- Zeng, B.G.; Zhang, S.J.; Chen, G.X. Investigation and Division of Fishery Resources in the South China Sea; In Chinese with English Abstract; Guangdong Science and Technology Press: Guangdong, China, 1989. [Google Scholar]
- Loots, C.; Vaz, S.; Koubbi, P.; Planque, B.; Coppin, F.; Verin, Y. Inter-annual variability of North Sea plaice spawning habitat. J. Sea Res. 2010, 64, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.Y.; Gao, Y.J.; Wang, J.P.; Xu, B.Q.; Sun, C.X. Community structure of ichthyoplankton and its relationship with environmental factors in Laizhou Bay. Chin. J. Ecol. 2018, 37, 2976–2984. [Google Scholar]
- Edwards, M.; Richardson, A.J. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature 2004, 430, 881. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Ruan, C.X.; Yuan, C.G. Effects of different temperature and food on growth and survival rates of the larvae of Trichogaster trichopterus. J. Fuji. Fish. 2008, 30, 9–12. [Google Scholar]
- Wan, R.J.; Meng, Z.N. The artificial insemination and hatching of Trichiurus lepturus. J. Fish. China 2003, 27, 188–192. [Google Scholar]
- Targońska, K.; Żarski, D.; Kupren, K.; Palińska-żarska, K.; Mamcarz, A.; Kujawa, R.; Skrzypczak, A.; Furgała-Selezniow, G.; Czarkowski, T.K.; Hakuć-Błażowska, A.; et al. Influence of temperature during four following spawning seasons on the spawning effectiveness of common bream, Abramis brama (L.) under natural and controlled conditions. J. Therm. Biol. 2014, 39, 17–23. [Google Scholar] [CrossRef]
- Wan, R.J.; Zeng, D.Y.; Bian, X.D.; Ni, X.B. Species composition and abundance distribution pattern of ichthyoplankton and their relationship with environmental factors in the East China Sea ecosystem. J. Fish. China 2014, 38, 1375–1398. [Google Scholar]
- Lelièvre, S.; Vaz, S.; Martin, C.S.; Loots, C. Delineating recurrent fish spawning habitats in the North Sea. J. Sea Res. 2014, 91, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.Z. The ecological characteristics of distribution of eggs, larvae and juveniles of the Engr-aulis japonicus (Temminck & Schlegel) and Anchoviella commersonii in the changjiang river estuary. Oceanol. Limnol. Sin. 1989, 20, 217–229. [Google Scholar]
- Zhang, J.B.; Huang, Z.Y. An investigation on fish eggs and larvae in sea area around planning Yangjiang nuclear plant. J. Trop. Oceanogr. 2003, 22, 78–84. [Google Scholar]
- Fernandez-Arcaya, U.; Drazen, J.C.; Murua, H.; Ramirez-Llodra, E.; Bahamon, N.; Recasens, L.; Rotllant, G.; Company, J.B. Bathymetric gradients of fecundity and egg size in fishes: A Mediterranean case study. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 116, 106–117. [Google Scholar] [CrossRef]
- Gibson, R.N.; Nash, R.D.; Geffen, A.J.; Van der Veer, H.W. Flatfishes: Biology and Exploitation; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Aneer, G. Herring (Clupea harengus L.) spawning and spawning ground characteristics in the Baltic Sea. Fish. Res. 1989, 8, 169–195. [Google Scholar] [CrossRef]
- Yin, M.C.; Blaxter, J.H.S. Cruising speeds during early development and starvation of marine fish larvae. Oceanol. Limnol. Sin. 1989, 20, 1–9. [Google Scholar]
- Yi, Y.J.; Zhang, S.H.; Wang, Z.Y. The bedform morphology of Chinese sturgeon spawning sites in the Yangtze River. Int. J. Sediment. Res. 2013, 28, 421–429. [Google Scholar] [CrossRef]
- Šaškov, A.; Šiaulys, A.; Bučas, M.; Daunys, D. Baltic herring (Clupea harengus membras) spawning grounds on the Lithuanian coast: Current status and shaping factors. Oceanologia 2014, 56, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.L.; Cao, R.X.; Shang, Q.T. Progress of Study on Coastal Circulation near the Shore of Western Guangdong. J. Guangdong Ocean Univ. 2012, 32, 94–98. [Google Scholar]
- Wu, B.Y. A study on the circulation in shelf waters west to zhujiang estuary. J. Appl. Oceanogr. 1990, 9, 14–21. [Google Scholar]
- Yang, S.Y.; Bao, X.W.; Chen, C.S.; Chen, F. Analysis on characteristics and mechanism of current system in west coast of Guangdong Province in the summer. Acta Oceanol. Sin. 2003, 25, 1–8. [Google Scholar]
- Guan, B.X. Evidence for a counter-wind current in winter off the southeast coast of China. Chin. J. Oceanol. Limn. 1986, 4, 319–332. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Factors | AIC Value | GCV Value | Adjusted R2 Value | Deviance Explained(%) | Residual Deviance |
|---|---|---|---|---|---|
| Log(Y + 1) = NULL | 1354.16 | 6.30 | 0 | 0.00 | 1808.76 |
| Log(Y + 1) = s(Month) | 1349.69 | 6.21 | 0.02 | 2.6 | 1761.76 |
| Log(Y + 1) = s(Month) + s(SSS) | 1251.74 | 4.43 | 0.32 | 34.7 | 1181.80 |
| Log(Y + 1) = s(Month) + s(SSS) + s(Depth) | 1214.40 | 3.89 | 0.41 | 43.5 | 1022.39 |
| Log(Y + 1) = s(Month) + s(SSS) + s(Depth) + s(SST) | 1169.28 | 3.34 | 0.51 | 54.2 | 829.06 |
| Log(Y + 1) = s(Month) + s(SSS) + s(Depth) + s(SST) + s(Chl-a) | 1161.02 | 3.26 | 0.52 | 56.8 | 781.28 |
| Log(Y + 1) = s(Month) + s(SSS) + s(Depth) + s(SST) + s(Chl-a) + s(Distance) | 1159.03 | 3.24 | 0.53 | 57.2 | 774.32 |
| Variables | Contribution (%) | d.f. | Pr(F) | Pr(chi) |
|---|---|---|---|---|
| SSS | 32.1 | 10.57 | 2.2 × 10−16 *** | 2.2 × 10−16 *** |
| SST | 10.7 | 9.02 | 5.7 × 10−9 *** | 4.6 × 10−10 *** |
| Depth | 8.8 | 2.82 | 8.3 × 10−9 *** | 1.9 × 10−9 *** |
| Month | 2.6 | 1.81 | 0.019 * | 0.018 * |
| Chl-a | 2.6 | 4.28 | 0.004 ** | 0.004 ** |
| Distance | 0.4 | 0.45 | 0.047 * | 0.046 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Yu, J.; Lin, Z.; Chen, P. Environmental Influence on the Spatiotemporal Variability of Spawning Grounds in the Western Guangdong Waters, South China Sea. J. Mar. Sci. Eng. 2020, 8, 607. https://doi.org/10.3390/jmse8080607
Lu Y, Yu J, Lin Z, Chen P. Environmental Influence on the Spatiotemporal Variability of Spawning Grounds in the Western Guangdong Waters, South China Sea. Journal of Marine Science and Engineering. 2020; 8(8):607. https://doi.org/10.3390/jmse8080607
Chicago/Turabian StyleLu, Yao, Jing Yu, Zhaojin Lin, and Pimao Chen. 2020. "Environmental Influence on the Spatiotemporal Variability of Spawning Grounds in the Western Guangdong Waters, South China Sea" Journal of Marine Science and Engineering 8, no. 8: 607. https://doi.org/10.3390/jmse8080607