Evaluation of the Effects of the Enriched-Organic Diets Composition on European Sea Bass Welfare through a Multi-Parametric Approach

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Consideration
2.2. Experimental Design and Rearing Conditions
2.3. Blood and Hepatic Sampling and Morphometric Procedures
2.4. Quantification of the Haematological and Serological Parameters and Enzymatic Activities
2.5. Critical Swimming Speed Tests and EMG Activity Monitoring
2.6. Growth Parameters Calculations
2.7. Statistical Analyses
3. Results
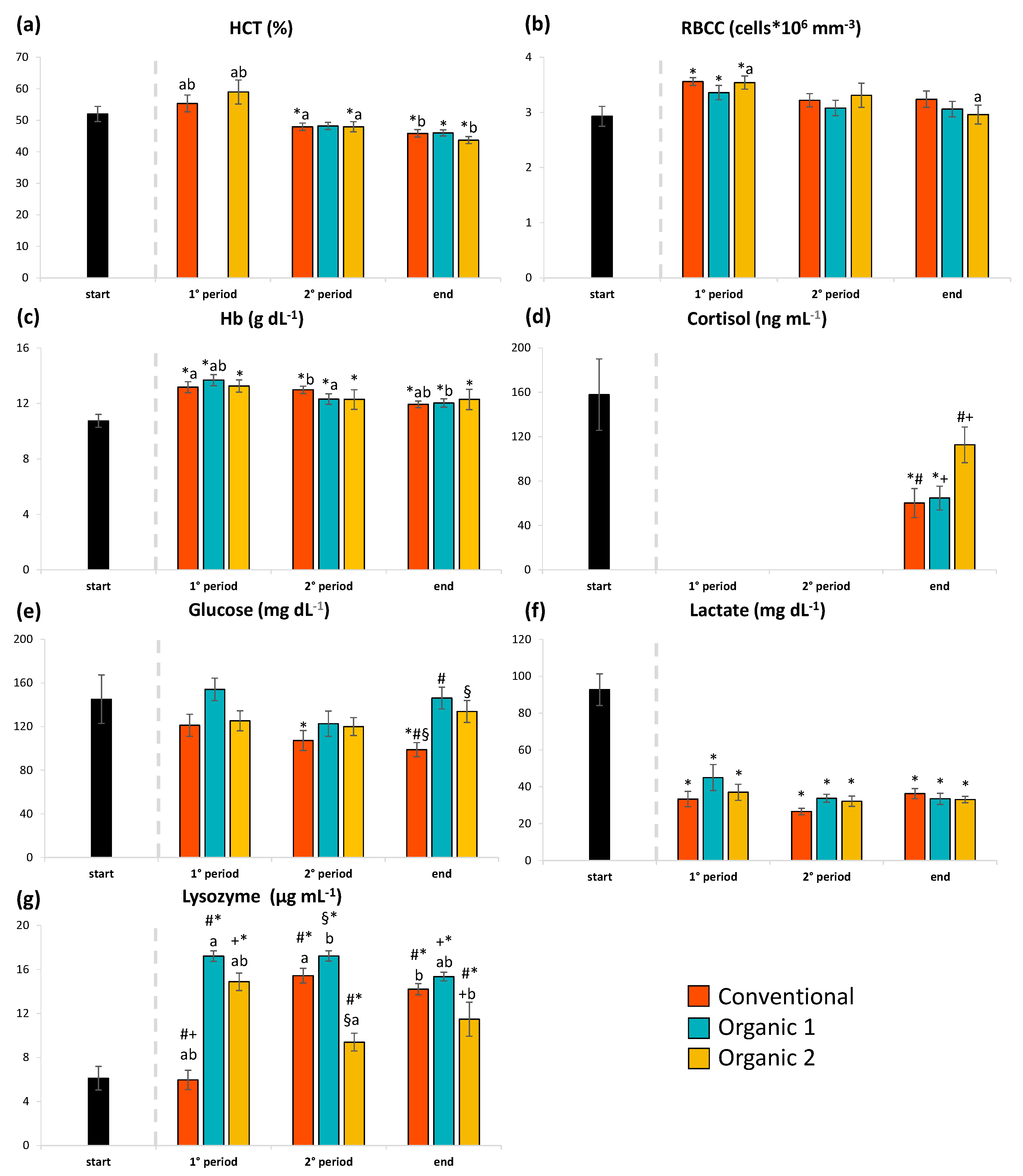
3.1. Haematological and Serological Parameters
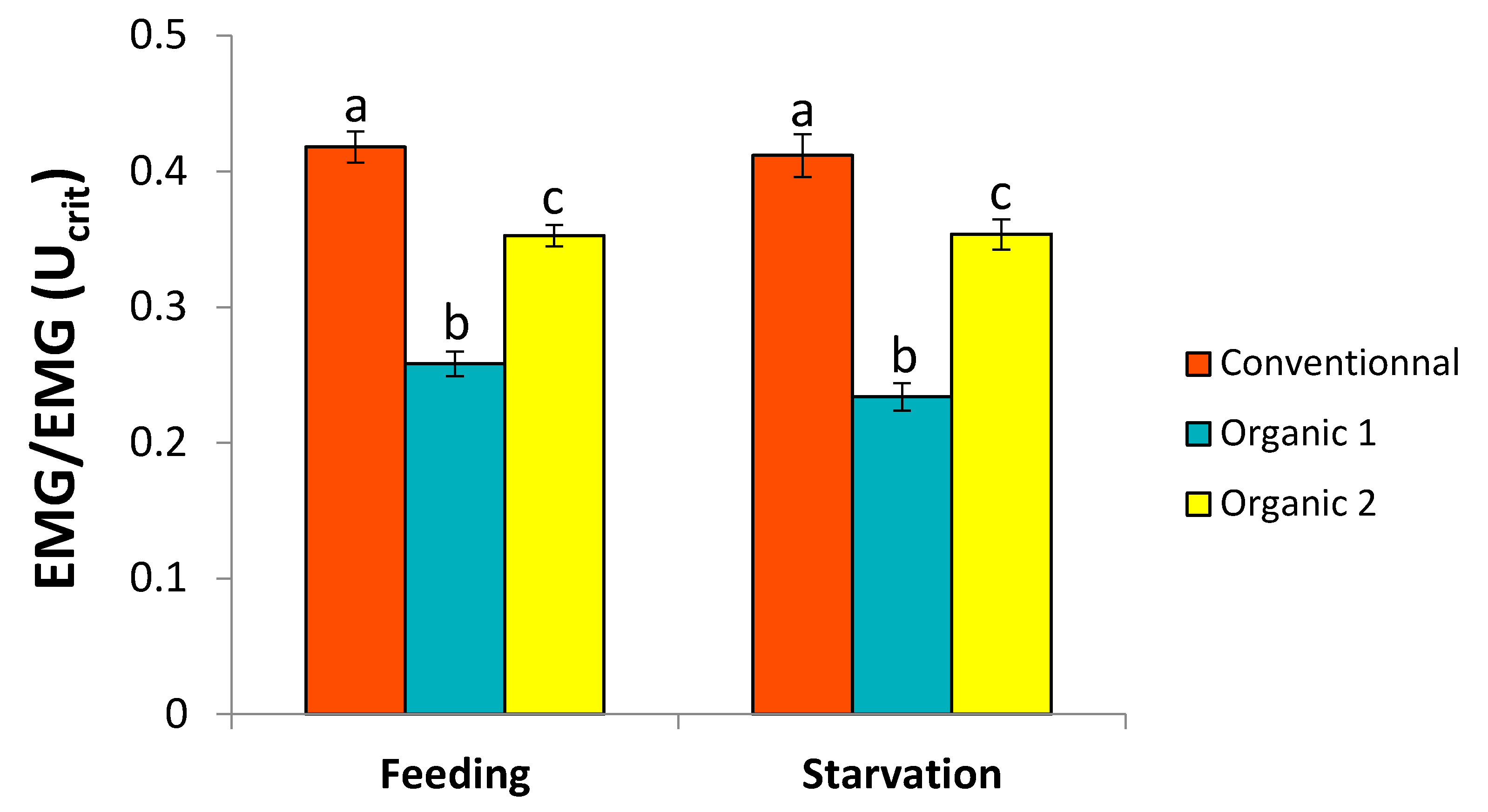
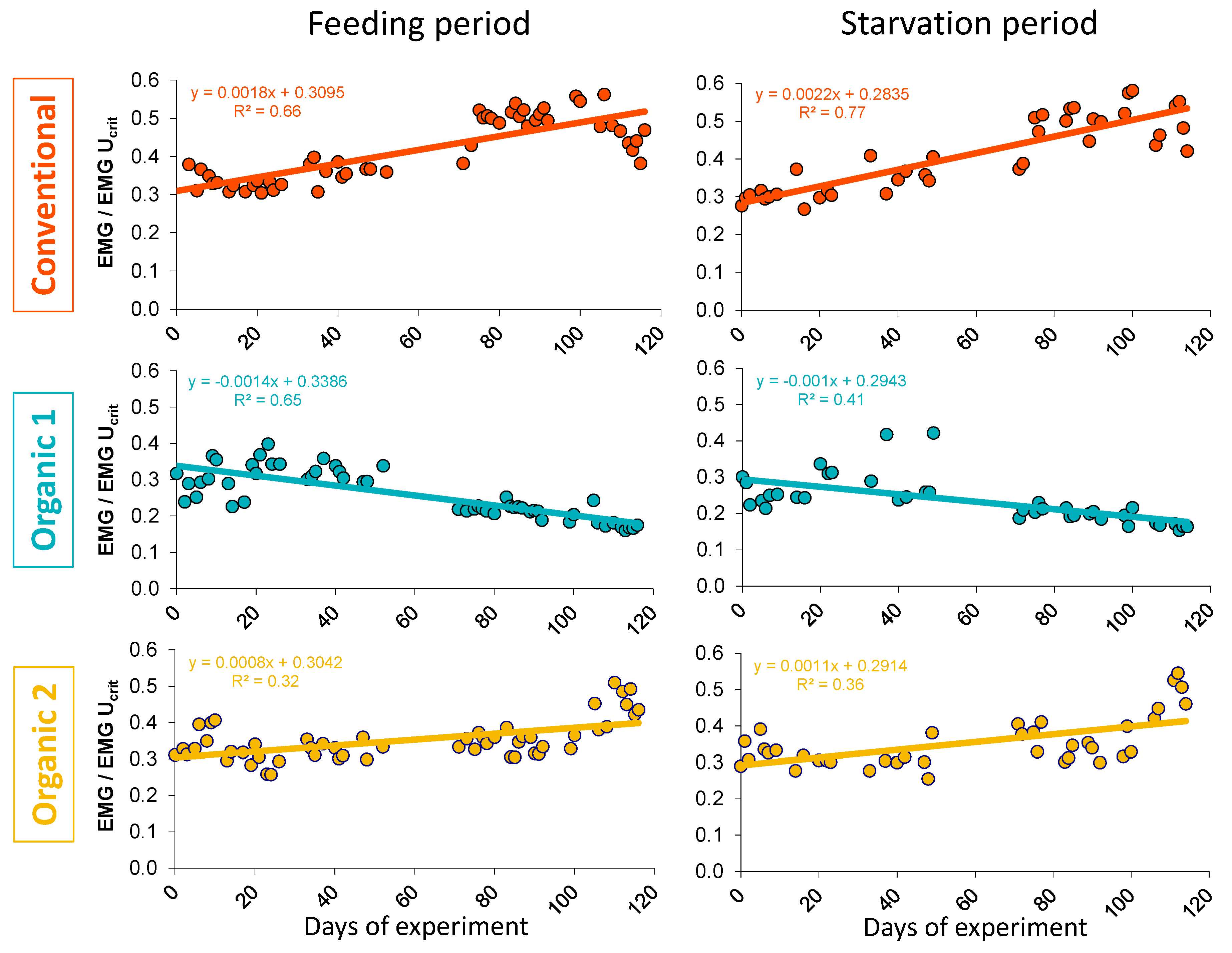
3.2. Physiological Parameters—Critical Swimming Speed and EMG
3.3. Specific Biomarkers—EROD and GST
3.4. Growth Parameters
3.5. PCA and NSFDSS Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The State of Fisheries and Aquaculture in the World 2018—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018; ISBN 9789251305621. [Google Scholar]
- Focardi, S.; Corsi, I.; Franchi, E. Safety issues and sustainable development of European aquaculture: New tools for environmentally sound aquaculture. Aquac. Int. 2005, 13, 3–17. [Google Scholar] [CrossRef]
- Ulbrich, B.; Stahlmann, R. Developmental toxicity of polychlorinated biphenyls (PCBs): A systematic review of experimental data. Arch. Toxicol. 2004, 78, 252–268. [Google Scholar] [CrossRef] [PubMed]
- Gould, D.; Compagnoni, A.; Lembo, G. Organic Aquaculture: Principles, Standards and Certification. In Organic Aquaculture Impacts and Future Developments; Springer: Cham, Switzerland, 2019; pp. 1–22. [Google Scholar]
- Mente, E.; Jokumsen, A.; Carter, C.G.; Antonopoulou, E.; Tacon, A.G. Chapter 8 Nutrition in relation to organic aquaculture: Sources and strategies. In Organic Aquaculture Impacts and Future Developments; Springer: Cham, Switzerland, 2019; pp. 141–188. [Google Scholar]
- Mente, E.; Karalazos, V.; Karapanagiotidis, I.T.; Pita, C. Nutrition in organic aquaculture: An inquiry and a discourse. Aquac. Nutr. 2011, 17. [Google Scholar] [CrossRef]
- Lembo, G.; Mente, E. Organic Aquaculture. Impacts and Future Developments; Springer: Cham, Switzerland, 2019; ISBN 9783030056025. [Google Scholar]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.-K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Kaushik, S.J.; Seiliez, I. Protein and amino acid nutrition and metabolism in fish: Current knowledge and future needs. Aquac. Res. 2010, 41, 322–332. [Google Scholar] [CrossRef]
- Palmegiano, G.B.; Gai, F.; Gasco, L.; Lembo, G.; Spedicato, M.T.; Trotta, P.; Zoccarato, I. Partial replacement of fish meal by T-Iso in gilthead sea bream (Sparus aurata) juveniles diets. Ital. J. Anim. Sci. 2009, 8, 869–871. [Google Scholar] [CrossRef]
- Montero, D.; Izquierdo, M. Welfare and health of fish fed vegetable oils as alternative lipid sources to fish oil. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; CRC Press: Boca Raton, FL, USA, 2010; pp. 439–486. [Google Scholar] [CrossRef]
- Geay, F.; Ferraresso, S.; Zambonino-Infante, J.L.; Bargelloni, L.; Quentel, C.; Vandeputte, M.; Kaushik, S.; Cahu, C.L.; Mazurais, D. Effects of the total replacement of fish meal and fish oil by plant sources on the hepatic transcriptome of European sea bass (Dicentrarchus labrax). In Proceedings of the Aquaculture Europe 11, Mediterranean Aquaculture 2020, Rhodes, Greece, 18–21 October 2011; pp. 377–378. [Google Scholar]
- Borquez, A.; Serrano, E.; Dantagnan, P.; Carrasco, J.; Hernandez, A. Feeding high inclusion of whole grain white lupin (Lupinus albus) to rainbow trout (Oncorhynchus mykiss): Effects on growth, nutrient digestibility, liver and intestine histology and muscle fatty acid composition. Aquac. Res. 2011, 42, 1067–1078. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.C.; Mai, K.S.; Tan, B.P.; Liu, Y.J. Partial replacement of fishmeal by soybean meal in diets for juvenile cobia (Rachycentron canadum). Aquac. Nutr. 2005, 11, 175–182. [Google Scholar] [CrossRef]
- Vandeputte, M.; Gagnaire, P.-A.; Allal, F. The European sea bass: A key marine fish model in the wild and in aquaculture. Anim. Genet. 2019, 195–206. [Google Scholar] [CrossRef]
- Teletchea, F. Domestication of marine fish species: Update and perspectives. J. Mar. Sci. Eng. 2015, 3, 1227–1243. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, M.S.; Obach, A. Dietary lipid sources for seabream and seabass: Growth performance, tissue composition and flesh quality. Aquaculture 2003, 9, 397–407. [Google Scholar] [CrossRef]
- Bonvini, E.; Bonaldo, A.; Mandrioli, L.; Sirri, R.; Dondi, F.; Bianco, C.; Fontanillas, R.; Mongile, F.; Gatta, P.P.; Parma, L. Effects of feeding low fishmeal diets with increasing soybean meal levels on growth, gut histology and plasma biochemistry of sea bass. Animal 2018, 12, 923–930. [Google Scholar] [CrossRef] [PubMed]
- FSBI. Fish Welfare. Briefing Paper; FSBI: Cambridge, UK, 2002; Volume 44. [Google Scholar]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Huntingford, F.A.; Kadry, S. Welfare and fish. In Fish Welfare; Branson, E.J., Ed.; Wiley: Blackwell, UK, 2008. [Google Scholar]
- Lembo, G.; Carbonara, P.; Fabris, A.; Manfrin, A.; Zupa, W. Welfare Issues and Veterinary Treatments. In Organic Aquaculture; Springer: Cham, Switzerland, 2019; pp. 119–140. [Google Scholar]
- Saraiva, J.L.; Arechavala-Lopez, P. Welfare of fish—No longer the elephant in the room. Fishes 2019, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, J.L.; Arechavala-Lopez, P.; Castanheira, M.F.; Volstorf, J.; Studer, B.H. A global assessment of welfare in farmed fishes: The fishethobase. Fishes 2019, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Dawkins, M.S. From an animal’s point of view: Motivation, fitness, and animal welfare. Behav. Brain Sci. 1990, 13, 1–9. [Google Scholar] [CrossRef]
- Arlinghaus, R.; Cowx, I.G.; Key, B.; Diggles, B.K.; Schwab, A.; Cooke, S.J.; Skiftesvik, A.B.; Browman, H.I. Pragmatic animal welfare is independent of feelings. Science (80) 2020, 370, 180. [Google Scholar]
- Huntingford, F.A.; Kadri, S. Taking account of fish welfare: Lessons from aquaculture. J. Fish Biol. 2009, 75, 2862–2867. [Google Scholar] [CrossRef]
- Huntingford, F.A.; Adams, C.; Braithwaite, V.A.; Kadri, S.; Pottinger, T.G.; Sandøe, P.; Turnbull, J.F. Current issues in fish welfare. J. Fish Biol. 2006, 44, 332–372. [Google Scholar] [CrossRef] [Green Version]
- Prunet, P.; Øverli, O.; Douxfils, J.; Bernardini, G.; Kestemont, P.; Baron, D. Fish welfare and genomics. Fish Physiol. Biochem. 2012, 38, 43–60. [Google Scholar] [CrossRef]
- Martos-sitcha, J.A.; Mancera, J.M.; Prunet, P.; Magnoni, L.J. Editorial: Welfare and Stressors in Fish: Challenges Facing Aquaculture. Front. Physiol. 2020, 11, 162. [Google Scholar] [CrossRef] [PubMed]
- Sadoul, B.; Geffroy, B. Measuring cortisol, the major stress hormone in fishes. J. Fish Biol. 2019, 94, 540–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, B.A. Stress in Fishes: A Diversity of Responses with Particular Reference to Changes in Circulating Corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Schreck, C.B.; Tort, L. The concept of stress in fish. In Fish Physiology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 35, pp. 1–34. ISBN 1546-5098. [Google Scholar]
- Carbonara, P.; Scolamacchia, M.; Spedicato, M.T.; Zupa, W.; Mckinley, R.S.; Lembo, G. Muscle activity as a key indicator of welfare in farmed European sea bass (Dicentrarchus labrax L. 1758). Aquac. Res. 2015, 46, 2133–2146. [Google Scholar] [CrossRef]
- Montero, D.; Izquierdo, M.S.; Tort, L.; Robaina, L.; Vergara, J.M. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiol. Biochem. 1999, 20, 53–60. [Google Scholar] [CrossRef]
- Huntingford, F.A. Implications of domestication and rearing conditions for the behaviour of cultivated fishes. J. Fish Biol. 2004, 65, 122–142. [Google Scholar] [CrossRef]
- Carbonara, P.; Alfonso, S.; Zupa, W.; Manfrin, A.; Fiocchi, E.; Pretto, T.; Spedicato, M.T.; Lembo, G. Behavioral and physiological responses to stocking density in sea bream (Sparus aurata): Do coping styles matter? Physiol. Behav. 2019, 212, 112698. [Google Scholar] [CrossRef]
- Varsamos, S.; Flik, G.; Pepin, J.F.; Bonga, S.E.W.; Breuil, G. Husbandry stress during early life stages affects the stress response and health status of juvenile sea bass, Dicentrarchus labrax. Fish Shellfish. Immunol. 2006, 20, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, J.; Bell, A.; Adams, C.; Bron, J.; Huntingford, F. Stocking density and welfare of cage farmed Atlantic salmon: Application of a multivariate analysis. Aquaculture 2005, 243, 121–132. [Google Scholar] [CrossRef]
- North, B.P.; Turnbull, J.F.; Ellis, T.; Porter, M.J.; Migaud, H.; Bron, J.; Bromage, N.R. The impact of stocking density on the welfare of rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 255, 466–479. [Google Scholar] [CrossRef]
- Carbonara, P.; Corsi, I.; Focardi, S.; Lembo, G.; Rochira, S.; Scolamacchia, M.; Spedicato, M.T.; Mckinley, R.S. The effects of stress induced by cortisol administration on the repeatability of swimming performance tests in the European sea bass (Dicentrarchus labrax L.). Mar. Freshw. Behav. Physiol. 2010, 43, 283–296. [Google Scholar] [CrossRef]
- Mormède, P.; Andanson, S.; Aupérin, B.; Beerda, B.; Guémené, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; van Reenen, C.G.; et al. Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Ellis, T.; Yildiz, H.Y.; López-Olmeda, J.; Spedicato, M.T.; Tort, L.; Øverli, Ø.; Martins, C.I.M. Cortisol and finfish welfare. Fish Physiol. Biochem. 2012, 38, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.I.M.; Galhardo, L.; Noble, C.; Damsgård, B.; Spedicato, M.T.; Zupa, W.; Beauchaud, M.; Kulczykowska, E.; Massabuau, J.C.; Carter, T.; et al. Behavioural indicators of welfare in farmed fish. Fish Physiol. Biochem. 2012, 38, 17–41. [Google Scholar] [CrossRef] [Green Version]
- Sadoul, B.; Mengues, P.E.; Friggens, N.C.; Prunet, P.; Colson, V. A new method for measuring group behaviours of fish shoals from recorded videos taken in near aquaculture conditions. Aquaculture 2014, 430, 179–187. [Google Scholar] [CrossRef]
- Alfonso, S.; Sadoul, B.; Cousin, X.; Bégout, M.-L. Spatial distribution and activity patterns as welfare indicators in response to water quality changes in European sea bass, Dicentrarchus labrax Article. Appl. Anim. Behav. Sci. 2020, 104974. [Google Scholar] [CrossRef]
- Shingles, A.; McKenzie, D.J.; Taylor, E.W.; Moretti, A.; Butler, P.J.; Ceradini, S. Effects of sublethal ammonia exposure on swimming performance in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 2001, 204, 2691–2698. [Google Scholar]
- McKenzie, D.J.; Höglund, E.; Dupont-Prinet, A.; Larsen, B.K.; Skov, P.V.; Pedersen, P.B.; Jokumsen, A. Effects of stocking density and sustained aerobic exercise on growth, energetics and welfare of rainbow trout. Aquaculture 2012, 338–341, 216–222. [Google Scholar] [CrossRef]
- Muñoz, L.; Aspillaga, E.; Palmer, M.; Saraiva, J.L.; Arechavala-Lopez, P. Acoustic Telemetry: A Tool to Monitor Fish Swimming Behavior in Sea-Cage Aquaculture. Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Cooke, S.J.; Thorstad, E.B.; Hinch, S.G. Activity and energetics of free-swimming fish: Insights from electromyogram telemetry. Fish Fish. 2004, 5, 21–52. [Google Scholar] [CrossRef]
- Whyte, J.J.; Jung, R.E.; Schmitt, C.J.; Tillitt, D.E. Ethoxyresorufin-O-deethylase (EROD) activity in fish as a biomarker of chemical exposure. Crit. Rev. Toxicol. 2000, 30, 347–570. [Google Scholar] [CrossRef] [PubMed]
- Teles, M.; Gravato, C.; Pacheco, M.; Santos, M. a Juvenile sea bass biotransformation, genotoxic and endocrine responses to beta-naphthoflavone, 4-nonylphenol and 17 beta-estradiol individual and combined exposures. Chemosphere 2004, 57, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Iversen, M.; Finstad, B.; McKinley, R.S.; Eliassen, R.A. The efficacy of metomidate, clove oil, Aqui-STM and Benzoak® as anaesthetics in Atlantic salmon (Salmo salar L.) smolts, and their potential stress-reducing capacity. Aquaculture 2003, 221, 549–566. [Google Scholar] [CrossRef]
- Corsi, I.; Mariottini, M.; Sensini, C.; Lancini, L.; Focardi, S. Cytochrome P450, acetylcholinesterase and gonadal histology for evaluating contaminant exposure levels in fishes from a highly eutrophic brackish ecosystem: The Orbetello Lagoon, Italy. Mar. Pollut. Bull. 2003, 46, 203–212. [Google Scholar] [CrossRef]
- Vissali, M.; Maricchiolo, M.V.; Genovese, L. Utilizzo dell’HPLC per la determinazione del cortisolo in specie ittiche. Biol. Mar. Mediterr. 2000, 7, 933–935. [Google Scholar]
- Burke, M.D.; Mayer, R.T. Ethoxyresorufin: Direct fluorimetric assay of a microsomal o-dealkylation which is preferentially inducible by 3-methylcholanthrene. Drug Metab. Dispos. 1974, 2, 583–588. [Google Scholar] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar]
- Bradford, M. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lembo, G.; Carbonara, P.; Scolamacchia, M.; Spedicato, M.T.; McKinley, R.S. Use of muscle activity indices as a relative measure of well-being in cultured sea bass Dicentrarchus labrax(Linnaeus, 1758). Hydrobiologia 2007, 582, 271–280. [Google Scholar] [CrossRef]
- McFarlane, W.J.; Cubitt, K.F.; Williams, H.; Rowsell, D.; Moccia, R.; Gosine, R.; McKinley, R.S. Can feeding status and stress level be assessed by analyzing patterns of muscle activity in free swimming rainbow trout (Oncorhynchus mykiss Walbaum)? Aquaculture 2004, 239, 467–484. [Google Scholar] [CrossRef]
- Lembo, G.; Carbonara, P.; Scolamacchia, M.; Spedicato, M.T.; Bjørnsen, J.E.; Holand, B.; McKinley, R.S. Introduction of a new physiological sensor: An acoustic EMG transmitter. Fish. Manag. Ecol. 2008, 15, 333–338. [Google Scholar] [CrossRef]
- McKinley, R.S.; Power, G. Measurement of activity and oxygen consumption for adult lake sturgeon (Acipenser fulvescens) in the wild using radiotransmitted EMG signals. In Wildlife Telemetry: Remote Monitoring and Tracking of Animals; Priede, I.G., Swift, S.M., Eds.; Ellis Horwood: West Sussex, UK, 1992; pp. 307–318. [Google Scholar]
- Alfonso, S.; Zupa, W.; Manfrin, A.; Fiocchi, E.; Dioguardi, M.; Dara, M.; Lembo, G.; Carbonara, P.; Cammarata, M. Surgical implantation of electronic tags does not induce medium-term effect: Insights from growth and stress physiological profile in two marine fish species. Anim. Biotelemetry 2020, 8, 21. [Google Scholar] [CrossRef]
- Jain, K.E.; Birtwell, I.K.; Farrell, A.P. Repeat swimming performance of mature sockeye salmon following a brief recovery period: A proposed measure of fish health and water quality. Can. J. Zool. 1998, 1496, 1488–1496. [Google Scholar] [CrossRef]
- Hopkins, K.D. Reporting Fish Growth: A Review of the Basics. J. World Aquac. Soc. 1992, 23, 173–179. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25. [Google Scholar] [CrossRef] [Green Version]
- Raîche, G.; Walls, T.A.; Magis, D.; Riopel, M.; Blais, J.G. Non-graphical solutions for Cattell’s scree test. Methodology 2013, 9, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Tam, C.M.; Tong, T.K.L.; Leung, A.W.T.; Chiu, G.W.C. Site Layout Planning using Nonstructural Fuzzy Decision Support System. J. Constr. Eng. Manag. 2002, 128, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Leung, P. Multiple-criteria decision-making (MCDM) applications in fishery management. Int. J. Environ. Technol. Manag. 2006, 6. [Google Scholar] [CrossRef]
- Belton, V.; Stewart, T. Multiple Criteria Decision Analysis—An Integrated Approach; Kluwer Academic: Boston, MA, USA, 2002. [Google Scholar]
- Chen, S.Y. Engineering Fuzzy Set Theory and Application; National Defense Industry Press: Beijing, China, 1998. [Google Scholar]
- Tam, C.M.; Tong, T.K.L.; Chiu, G.W.C. Comparing non-structural fuzzy decision support system and analytical hierarchy process in decision-making for construction problems. Eur. J. Oper. Res. 2006, 174, 1317–1324. [Google Scholar] [CrossRef]
- Lembo, G.; Bellido, J.M.; Bitetto, I.; Facchini, M.T.; García-Jiménez, T.; Stithou, M.; Vassilopoulou, V.C.; Spedicato, M.T. Preference modeling to support stakeholder outreach toward the common fishery policy objectives in the north Mediterranean Sea. Front. Mar. Sci. 2017, 4, 328. [Google Scholar] [CrossRef] [Green Version]
- Tong, T.K.L. Free Multi-Criteria Decision Aiding (MCDA) Tools for Research Students 2008. Available online: https://sites.google.com/site/mcdafreeware/ (accessed on 17 November 2020).
- FAO. Fish as Feed Inputs for Aquaculture. Practices, Sustainability and Implications; Hasan, M.R., Halwart, M., Eds.; FAO: Rome, Italy, 2009; ISBN 9789251064191. [Google Scholar]
- Okey, A.B. Enzyme induction in the cytochrome P450 system. Pharmacology 1990, 45, 241–298. [Google Scholar]
- Novi, S.; Pretti, C.; Cognetti, A.M.; Longo, V.; Marchetti, S.; Gervasi, P.G. Biotransformation enzymes and their induction by β-naphtoflavone in adult sea bass (Dicentrarchus labrax). Aquat. Toxicol. 1998, 41, 63–81. [Google Scholar] [CrossRef]
- Gravato, C.; Santos, M.A. Beta-naphthoflavone liver EROD and erythrocytic nuclear abnormality induction in juvenile Dicentrarchus labrax L. Ecotoxicol. Environ. Saf. 2002, 52, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Teles, M.; Pacheco, M.; Santos, M. a Biotransformation, stress and genotoxic effects of 17beta-estradiol in juvenile sea bass (Dicentrarchus labrax L.). Environ. Int. 2006, 32, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Tysklind, M.; Bosveld, A.B.T.; Patrik, A.; Eske, V.; Theo, S.; Willem, S.; Rappe, R.C.; van den Berg, M. Inhibition of ethoxyresorufin-O-deethylase (EROD) activity in mixtures of 2,3,7,8-tetrachlorodibenzo-p-dioxin and polychlorinated biphenyls. Environ. Sci. Pollut. Res. 1995, 211. [Google Scholar] [CrossRef]
- Sadoul, B.; Vijayan, M.M. Stress and Growth. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 167–205. ISBN 9780128027288. [Google Scholar]
- Samaras, A.; Santo, C.E.; Papandroulakis, N.; Mitrizakis, N.; Pavlidis, M.; Höglund, E.; Pelgrim, T.N.M.; Zethof, J.; Spanings, F.A.T.; Vindas, M.A.; et al. Allostatic load and stress physiology in European seabass (Dicentrarchus labrax L.) and gilthead seabream (Sparus aurata L.). Front. Endocrinol. (Lausanne) 2018, 9, 451. [Google Scholar] [CrossRef] [Green Version]
- Alfonso, S.; Sadoul, B.; Gesto, M.; Joassard, L.; Chatain, B.; Geffroy, B.; Bégout, M.-L. Coping styles in European sea bass: The link between boldness, stress response and neurogenesis. Physiol. Behav. 2019, 207, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Mourente, G.; Good, J.E.; Thompson, K.D.; Bell, J.G. Effects of partial substitution of dietary fish oil with blends of vegetable oils, on blood leucocyte fatty acid compositions, immune function and histology in European sea bass (Dicentrarchus labrax L). Br. J. Nutr. 2007, 98, 770–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balfry, S.K.; Maule, G.; Iwama, G.K. Coho salmon Oncorhynchus kisutchstrain differences in disease resistance and non-specific immunity, following immersion challenges with Vibrio anguillarum. Dis. Aquat. Organ. 2001, 47, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Weyts, F.A.A.; Cohen, N.; Flik, G.; Verburg, V.K.B.M.L. Interactions between the immune system and the hypothalamo-pituitary-interrenal axis in fish. Fish Shellfish Immunol. 1999, 9, 1–20. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Pérez-Sànchez, J. Diet related changes in non-specific immune response of European sea bass (Dicentrarchus labrax L.). Fish Shellfish Immunol. 1999, 9, 637–640. [Google Scholar] [CrossRef]
- Boshra, H.; Li, J.; Sunyer, J.O. Recent advances on the complement system of teleost fish. Fish Shellfish Immunol. 2006, 20, 239–262. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.A.M.; Król, E. Nutrigenomics and immune function in fish: New insights from omics technologies. Dev. Comp. Immunol. 2017, 75, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.M.; Friesen, E.N.; Higgs, D.A.; Farrell, A.P. The effect of dietary lipid and protein source on the swimming performance, recovery ability and oxygen consumption of Atlantic salmon (Salmo salar). Aquaculture 2007, 273, 687–699. [Google Scholar] [CrossRef]
- McKenzie, D.J.; Higgs, D.A.; Dosanjh, B.S.; Deacon, G.; Randall, D.J. Dietary fatty acid composition influences swimming performance in Atlantic salmon (Salmo salar) in seawater. Fish Physiol. Biochem. 1998, 19, 111–122. [Google Scholar] [CrossRef]
- Wagner, G.N.; Balfry, S.K.; Higgs, D.A.; Lall, S.P.; Farrell, A.P. Dietary fatty acid composition affects the repeat swimming performance of Atlantic salmon in seawater. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2004, 137, 567–576. [Google Scholar] [CrossRef]
- Milligan, C.L. A regulatory role for cortisol in muscle glycogen metabolism in rainbow trout Oncorhynchus mykiss Walbaum. J. Exp. Biol. 2003, 206, 3167–3173. [Google Scholar] [CrossRef] [Green Version]
- Korte, S.M.; Olivier, B.; Koolhaas, J.M. A new animal welfare concept based on allostasis. Physiol. Behav. 2007, 92, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Zupa, W.; Carbonara, P.; Spedicato, M.T.; Lembo, G. Modelling swimming activities and energetic costs in European sea bass (Dicentrarchus labrax L., 1758) during critical swimming tests. Mar. Freshw. Behav. Physiol. 2015, 48, 341–357. [Google Scholar] [CrossRef]
- Chandroo, K.P.; Cooke, S.J.; McKinley, R.S.; Moccia, R.D. Use of electromyogram telemetry to assess the behavioural and energetic responses of rainbow trout, Oncorhynchus mykiss (Walbaum) to transportation stress. Aquac. Res. 2005, 36, 1226–1238. [Google Scholar] [CrossRef]
- Cooke, S.J.; Chandroo, K.P.; Beddow, T.A.; Moccia, R.D.; McKinley, R.S. Swimming activity and energetic expenditure of captive rainbow trout Oncorhynchus mykiss (Walbaum) estimated by electromyogram telemetry. Aquac. Res. 2000, 31, 495–505. [Google Scholar] [CrossRef]
- Paige, J.A.; Liao, R.; Hajjar, R.J.; Foisy, R.L.; Cory, C.R.; O’Brien, P.J.; Gwathmey, J.K. Effect of a high omega-3 fatty acid diet on cardiac contractile performance in Oncorhynchus mykiss. Cardiovasc. Res. 1996, 31, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Mckenzie, D.J. Effects of dietary fatty acids on the respiratory and cardiovascular physiology of fish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 128, 605–619. [Google Scholar] [CrossRef]
- Bonaldo, A.; Roem, A.J.; Fagioli, P.; Pecchini, A.; Cipollini, I.; Gatta, P.P. Influence of dietary levels of soybean meal on the performance and gut histology of gilthead sea bream (Sparus aurata L.) and European sea bass (Dicentrarchus labrax L.). Aquac. Res. 2008, 39, 970–978. [Google Scholar] [CrossRef]
- Yildiz, M.; Sener, E.; Timur, M. Effects of variations in feed and seasonal changes on body proximate composition of wild and cultured sea bass (Dicentrarchus labrax L.). Turkish J. Fish. Aquat. Sci. 2007, 7, 45–51. [Google Scholar]
- Montero, D.; Robaina, L.; Caballero, M.J.; Ginés, R.; Izquierdo, M.S. Growth, feed utilization and flesh quality of European sea bass (Dicentrarchus labrax) fed diets containing vegetable oils: A time-course study on the effect of a re-feeding period with a 100% fish oil diet. Aquaculture 2005, 248, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Lanari, D.; Agaro, E.D. Alternative plant protein sources in sea bass diets. Ital. J. Anim. Sci. 2005, 4, 365–374. [Google Scholar] [CrossRef]
- Kavadias, B.S.; Dessypris, A. Annual cycles of growth rate, feeding rate, food conversion, plasma glucose and plasma lipids in a population of European sea bass (Dicentrarchus labrax L.) farmed in floating marine cages. J. Appl. Ichthyol. 2003, 19, 29–34. [Google Scholar] [CrossRef]
- Sadoul, B.; Leguen, I.; Colson, V.; Friggens, N.C.; Prunet, P. A multivariate analysis using physiology and behavior to characterize robustness in two isogenic lines of rainbow trout exposed to a confinement stress. Physiol. Behav. 2015, 140, 139–147. [Google Scholar] [CrossRef]
- Botreau, R.; Veissier, I.; Perny, P.; Capdeville, J. Multicriteria evaluation of animal welfare at farm level: An application of MCDA methodologies. Found. Comput. Decis. Sci. 2008, 33, 287–316. [Google Scholar]
- Houston, A.H. Review: Are the Classical Hematological Variables Acceptable Indicators of Fish Health? Trans. Am. Fish. Soc. 2004, 126, 879–894. [Google Scholar] [CrossRef]
- Carbonara, P.; Dioguardi, M.; Cammarata, M.; Zupa, W.; Vazzana, M.; Spedicato, M.T.; Lembo, G. Basic knowledge of social hierarchies and physiological profile of reared sea bass Dicentrarchus labrax (L.). PLoS ONE 2019, 14, e0208688. [Google Scholar] [CrossRef] [PubMed]
- Sonesson, U.; Davis, J.; Ziegler, F. Food Production and Emissions of Greenhouse Gases: An Overview of the Climate Impact of Different Product Groups; SIK Institutet för Livsmedel Och Bioteknik: Gothenborg, Sweden, 2010; p. 26. [Google Scholar]
- Angel, D.; Jokumsen, A.; Lembo, G. Aquaculture Production Systems and Environmental Interactions. In Organic Aquaculture; Springer: Cham, Switzerland, 2019; pp. 103–118. ISBN 9783030056032. [Google Scholar]
- Mie, A.; Andersen, H.R.; Gunnarsson, S.; Kahl, J.; Kesse-Guyot, E.; Rembiałkowska, E.; Quaglio, G.; Grandjean, P. Human health implications of organic food and organic agriculture: A comprehensive review. Environ. Heal. A Glob. Access Sci. Source 2017, 16, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | Tank | n | Mean Weight (g) | Biomass (kg) | Mean Length (cm) | Density (kg m−3) |
|---|---|---|---|---|---|---|
| DAN-EX 1754 (Conventional) | 1 | 42 | 297.8 ± 15.4 | 12.51 | 302 ± 6.00 | 10.43 |
| 2 | 41 | 336.4 ± 24 | 13.79 | 313 ± 7.93 | 11.49 | |
| 3 | 42 | 324.9 ± 16.6 | 13.65 | 312 ± 5.40 | 11.38 | |
| DAN-EC 1650 (Organic 1) | 4 | 42 | 317.5 ± 14.1 | 13.34 | 308 ± 4.15 | 11.12 |
| 5 | 42 | 312.7 ± 21.4 | 13.13 | 295 ± 9.69 | 10.94 | |
| 6 | 42 | 339.1 ± 28.9 | 14.24 | 315 ± 9.78 | 11.87 | |
| DAN-EC 2640 (Organic 2) | 7 | 42 | 327.8 ± 17.5 | 13.77 | 305 ± 6.85 | 11.48 |
| 8 | 40 | 319.3 ± 23.6 | 12.77 | 306 ± 7.75 | 10.64 | |
| 9 | 41 | 322.2 ± 13.3 | 13.2 | 306 ± 7.46 | 11 |
| Raw Material (%) | Conventional | Organic 1 | Organic 2 | |
|---|---|---|---|---|
| Rough proteins | 54 | 50 | 40 | |
| Gross fats | 17 | 16 | 26 | |
| Rough ashes | 9.16 | 10.59 | 8.64 | |
| Rough fibers | 2.01 | 2.89 | 3.39 | |
| Phosphorus | 1.27 | 1.74 | 1.31 | |
| Additives | ||||
| Copper (mg kg−1) | 5 | 5 | 5 | |
| Vitamin A (I.U. g−1) | 0.72 | 0.72 | 0.72 | |
| Vitamin D3 (I.U. g−1) | 0.11 | 0.11 | 0.11 | |
| Vitamin E (mg kg−1) | 198 | 198 | 198 | |
| BHT (mg kg−1) | 79 | 0 | 0 | |
| Rosmary essential oil (mg kg−1) | 0 | 200 | 200 | |
| Proteic compounds (%) | Proteic compounds in organic diets (%) | |||
| Fish flour | 54.65 | Fish flour | 56.88 | 38.25 |
| Soybean proteic concentrate | 15 | Roasted organic soy extract | 10.49 | 11.10 |
| Fish oil | 10.07 | Fish oil | 5.23 | 17.29 |
| Wheat | 8.83 | Organic wheat | 12.50 | 12 |
| Wheat gluten | 4 | Extract of organic colza | 2.20 | 5.99 |
| Hulled roasted soy flour | 4 | Roasted organic soybeans | 10 | 11.10 |
| Peas | 3 | Extract of organic sunflower seed | 2.20 | 3.76 |
| Vitamin pre-mix | 0.45 | Vitamin pre-mix | 0.50 | 0.50 |
| Diet | Mean Weight (g) | Ucrit1 (BL s−1) | Ucrit2 (BL s−1) | Recovery Ratio |
|---|---|---|---|---|
| Conventional | 403.7 ± 8.9 | 4.1 ± 0.14 | 3.8 ± 0.15 | 0.93 ± 0.03 * |
| Organic 1 | 416.9 ± 19.1 | 3.9 ± 0.10 | 3.8 ± 0.14 | 0.99 ± 0.02 |
| Organic 2 | 403.8 ± 15.14 | 3.6 ± 0.10 | 3.5 ± 0.02 | 0.98 ± 0.02 |
| Parameter | Start | Conventional | Organic 1 | Organic 2 |
|---|---|---|---|---|
| EROD (nmol min−1 mg−1) | 50.1 ± 3.58 | * 37.5 ± 4.96 | 47.3 ± 3.28 | * 38.4 ± 3.38 |
| GST (nmol min−1 mg−1) | 60.0 ± 2.66 | 56.3 ± 4.21 | 58.2 ± 5.65 | 50.4 ± 4.65 |
| HSI (%) | 1.03 ± 0.05 | *, #, + 1.66 ± 0.06 | *, #, § 1.99 ± 0.07 | *, +, § 2.3 ± 0.09 |
| TL (mm) | 306.9 ± 2.4 | #, + 350 ± 2.3 | # 357± 2.7 | + 360 ± 2.6 |
| Weigth (g) | 321.9 ± 6.7 | #, + 509.9 ± 11.43 | # 546.7 ± 12.76 | + 545.6 ± 11.76 |
| SGR (%) | #, + 0.29 ± 0.011 | # 0.33 ± 0.01 | + 0.33 ± 0.005 | |
| FCR | 5.31 ± 0.680 | 4.92 ± 0.213 | 5.06 ± 0.356 | |
| PER (%) | 36.01 ± 4.37 | 40.82 ± 4.29 | 49.90 ± 3.54 | |
| Survival rate (%) | 82 | 78 | 77 |
| Variables | Component 1 | Component 2 | Component 3 |
|---|---|---|---|
| HCT | 0.38 | ||
| RBCC | 0.23 | 0.54 | |
| HB | 0.26 | 0.52 | |
| Cortisol | 0.19 | 0.58 | 0.52 |
| Glucose | 0.31 | 0.2 | 0.63 |
| Lactate | |||
| Lysozyme | −0.51 | −0.29 | |
| EROD | 0.64 | −0.67 | 0.31 |
| GST | −0.17 | −0.83 | 0.5 |
| HSI | 0.41 | 0.32 | |
| SGR | 0.98 | ||
| RR | 0.16 | 0.31 | |
| EMG | −0.54 | 0.45 | |
| PER | 0.67 | 0.59 | −0.41 |
| FCR | −0.95 | 0.26 | |
| Weight gain | 0.98 | ||
| Variance explained (%) | 27.3 | 16.4 | 13.2 |
| Diet | Hypothesis 1 | Hypothesis 2 |
|---|---|---|
| Organic 1 | 1.000 | 0.928 |
| Conventional | 0.953 | 0.901 |
| Organic 2 | 0.428 | 0.520 |
| Diet | EMG | RR | Cortisol | SGR | HSI | Glucose | Lysozyme |
|---|---|---|---|---|---|---|---|
| Conventional | 1 | 1 | 1 | 0.333 | 1 | 0.333 | 1 |
| Organic 1 | 1 | 0.333 | 1 | 1 | 0.538 | 1 | 1 |
| Organic 2 | 0.333 | 0.333 | 0.333 | 1 | 0.25 | 1 | 0.333 |
| Parameters (Criteria) | Score |
|---|---|
| EMG | 1 |
| RR | 0.333 |
| Cortisol | 0.176 |
| SGR | 0.111 |
| HSI | 0.053 |
| Glucose | 0.00 |
| Lysozyme | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbonara, P.; Zupa, W.; Bitetto, I.; Alfonso, S.; Dara, M.; Cammarata, M. Evaluation of the Effects of the Enriched-Organic Diets Composition on European Sea Bass Welfare through a Multi-Parametric Approach. J. Mar. Sci. Eng. 2020, 8, 934. https://doi.org/10.3390/jmse8110934
Carbonara P, Zupa W, Bitetto I, Alfonso S, Dara M, Cammarata M. Evaluation of the Effects of the Enriched-Organic Diets Composition on European Sea Bass Welfare through a Multi-Parametric Approach. Journal of Marine Science and Engineering. 2020; 8(11):934. https://doi.org/10.3390/jmse8110934
Chicago/Turabian StyleCarbonara, Pierluigi, Walter Zupa, Isabella Bitetto, Sébastien Alfonso, Mariano Dara, and Matteo Cammarata. 2020. "Evaluation of the Effects of the Enriched-Organic Diets Composition on European Sea Bass Welfare through a Multi-Parametric Approach" Journal of Marine Science and Engineering 8, no. 11: 934. https://doi.org/10.3390/jmse8110934